
Christopher C. Onyeke
Department of Plant Science and Biotechnology, University of Nigeria, Nsukka, Enugu State, Nigeria
Godswill C. Ajuziogu
Department of Plant Science and Biotechnology, University of Nigeria, Nsukka, Enugu State, Nigeria
Emmanuel E. Osayi
Department of Plant Science and Biotechnology, University of Nigeria, Nsukka, Enugu State, Nigeria
Christopher D. Nwani
Department of Zoology and Environmental Biology, University of Nigeria, Nsukka, Enugu State, Nigeria
Gregory E. Odo
Department of Zoology and Environmental Biology, University of Nigeria, Nsukka, Enugu State, Nigeria
Plant Pathology Journal
Year: 2014 | Volume: 13 | Issue: 2 | Page No.: 112-119







ABSTRACT
Studies were carried out in screen house to determine the effects of different inoculum densities (2,000 and 4,000 eggs of Meloidogyne javanica) and different levels (15, 30, 45, 60 and 75 t ha-1) of leaf powder of Hyptis suaveolens soil amendment on the pathogenicity of M. javanica on African yam bean. Results showed that organic soil amendment with H. suaveolens improved on the growth of plants infected with eggs of M. javanica as against the infected plants but unamended. It was also observed in this study that higher soil amendment levels (>30 t ha-1) led to sharp progressive decline in the fresh shoot and root weights of amended plants, which indicates possible phytotoxicity of H. suaveolens leaf powder at amendment levels above 30 t ha-1. H. suaveolens leaf powder amendment reduced significantly (p≤0.05) the number of galls, egg masses, gall index and egg mass index as the amendment levels increased at both low and high inoculum densities. M. javanica reduced significantly (p≤0.05) the number of nodules in infected test plants compared to the uninfected controls. This study also revealed that the pathogenicity of M. javanica was more at high inoculum density generally. Conclusively, this study has shown that application of H. suaveolens as soil amendment reduced the pathogenicity of M. javanica and improved on the growth parameters of the test plant but at higher levels (above 30 t ha-1) it became phytotoxic and highly nematicidal.
PDF Abstract XML References Citation
Received: December 15, 2013;
Accepted: January 18, 2014;
Published: April 19, 2014
How to cite this article
Christopher C. Onyeke, Godswill C. Ajuziogu, Emmanuel E. Osayi, Christopher D. Nwani and Gregory E. Odo, 2014. Efficacy of Soil Amendment with Leaf Powder of Hyptis suaveolens
(L.) Poit in the Control of Meloidogyne javanica (Treub) Chitwood on the African Yam
Bean, Sphenostylis stenocarpa (Hochst Ex A. Rich) Harms. Plant Pathology Journal, 13: 112-119.
DOI: 10.3923/ppj.2014.112.119
URL: https://scialert.net/abstract/?doi=ppj.2014.112.119
DOI: 10.3923/ppj.2014.112.119
URL: https://scialert.net/abstract/?doi=ppj.2014.112.119
INTRODUCTION
Meloidogyne species are economically the most destructive plant parasitic nematode world-wide causing serious losses in agricultural productivity and quality (Sasser, 1979; Sasser and Carter, 1985; Cristobal-Alejo et al., 2006). They infect a wide range of plants, world-wide in distribution and have been reported in many important economic crops where they reduce crop productivity (Whitehead, 1998; Ray et al., 1995; Afolami and Orisajo, 2003). On the average, losses in crop productivity due to root knot nematodes are estimated to be about 5% though much larger for developing countries in the tropics and subtropics (Taylor and Sasser, 1978).
In recent times, management of plant parasitic nematodes using chemical nematicides has become unpopular because of high cost, scarcity, inherent toxicity of many synthentic pesticides to many non-target organisms and their residual effects on the environment (Haseeb and Butool, 1996; Olabiyi et al., 2011). Also, the restriction of some of the effective chemical nematicides (Methyl bromide, EDB, DBCP etc.) from the pesticide market necessitated studies on the possible roles of organic soil amendments in the control of plant parasitic nematode populations in the soil (Ogbuji, 1981; Agu, 2008; Olabiyi, 2008; Muazu and Umar, 2012).
The use of plant based/organic material as soil amendment to manage the activities of plant parasitic nematodes on crop plants has the advantages of biodegradability, environmental safety, renewability and selective toxicity to target pests (Saravanapriya and Sivakumar, 2005). Encouraging results have been obtained by researchers using organic material to discourage reproduction and development of nematodes and losses in crop plants. Some of the organic materials that have shown antinematode properties include, African marigold (Tagetes erecta), rattle weed (Crotolaria retusa), basil (Ocimum gratissimum), lemon grass (Cymbopogon citratus), neem (Azadirachta indica), siam weed (Chromolaena odorata) and sugar cane bagasse (Saravanapriya and Sivakumar, 2005; Hayat et al., 2012; Onyeke and Akueshi, 2012).
African yam bean, Sphenostylis stenocarpa (Hochst Ex A. Rich) Harms belongs to the family Fabaceae (Syn. Leguminosae) and classified under the sub-Family Papilionoideae (Heywood, 1971). It is reported to have total seed protein content which varies from 19.5-29% on a dry weight basis, values similar to that reported for cowpea (Vigna unguiculata (L.) Walp; 24%) but low compared to those of winged bean (Psophocarpus tetragonolobus (L.) D.C.; 32.8%) and soybean (Glycine max (L.) Merril; 35.1% (Duke, 1981). Nutritionally, S. stenocarpa has the potential of substituting animal proteins in the diets of many people who cannot afford animal proteins. Unfortunately, African yam bean is susceptible to plant parasitic nematodes and has been identified as a major constraint limiting the growth and yield of this important legume (Agu, 2008). The present study was carried out to determine the effect of different inoculum densities and levels of leaf powder of H. suaveolens soil amendment on the pathogenicity of M. javanica on African yam bean.
MATERIALS AND METHODS
Viable seeds of the African yam bean (TSS 11 and TSS 112) used in this study were collected from the seed bank of the International Institute for Tropical Agriculture (IITA), Ibadan, Nigeria.
Leaves of Hyptis suaveolens used for the experiment were collected from the University of Nigeria, Nsukka premises. The leaves were air-dried indoors until completely dried and then ground into fine powder with Thomas Wiley Laboratory Mill Model 4, Arthur Thomas Company.
The preliminary qualitative phytochemical screening of leaves of H. suaveolens was performed according to procedures outlined by Harborne (1973) and Trease and Evans (1996).
Nsukka population of Meloidogyne javanica maintained for up to 8 weeks on susceptible African yam bean accession (TSS 112) served as source of inoculum. The galled roots of the African yam bean were carefully uprooted, washed free of soil and cut into 1-2 cm segments.The eggs were extracted from the galls using the NaOCL-extraction method (Baker, 1985). Extracted eggs were passed through a 75 μm sieve and poured on 5 μm sieve immersed in a water bath for a proper dilution of NaOCL. Furthermore, eggs were carefully washed with sterile distilled water to wash away any possible residual NaOCL. The extracted eggs were washed into a graduated beaker and the volume made up to 1,000 mL with sterile distilled water. The egg suspension was calibrated in such a way that 1 mL aliquots contained 2, 00 eggs; 20 mL of the egg suspension contained 4, 000 eggs of M. javanica and 10 mL of the egg suspension contained 2, 000 eggs of M. javanica.
The experiment was conducted at the Department of Plant Science and Biotechnology Screen House, University of Nigeria, Nsukka (Latitude 06°86' 39.6" N, Longitude 007°41' 20.4" E and Altitude 433 m above sea level). Top garden soil collected from fallow land was sterilized in a large drum by cooking to a temperature of 100°C for 3 h and later air-dried for a period of 4 weeks before commencing the study to avoid any possible side effects of cooking (Anderson and Ingram, 1989). The physical and chemical properties of the soil were determined according to standard methods (IITA, 1989). The textural class of the soil was sandy loam (48% coarse sand, 27% fine sand, 10% silt, 15% clay) pH (in H20) 6.2, pH (in Kcl) 5.7, OM 1.58%, BS 58.06%, N 0.098%, CEC 12.4 μ 1 00 g-1, Na 0.11 μ 100 g-1, K 0.09 μ100 g-1, Ca 4.8 μ100 g-1, Mg 2.2 μ100 g-1, exchangeable acidity 0.8 μ100 g-1 and P 13.07 ppm. The sterilized and air-dried soil was mixed thoroughly with each of the leaf powder at different amendment levels (15, 30, 45, 60, 75 t ha-1). The amended soils were poured into polyethylene planting bags of 20.3 cm diameter and 25.5 cm depth. Soils without amendments (0 t ha-1) served as controls. Wholesome seeds of the African yam bean were surface sterilized with 0.1% mercuric chloride for 2 min and rinsed in three changes of distilled sterile water and planted two per bag but later reduced to one after germination. Two weeks after germination the polyethylene bags were inoculated with (2,000; 4,000 eggs) the egg suspensions by pouring into a shallow trench created around the root tips of each of the test plants (Hussey and Bourma, 1981) and covered immediately with top soil (Goswami and Chenulu, 1974). Plants that were not inoculated served as controls. The planting bags were arranged in a Completely Randomized Design (CRD) with ten replications. Each bag was watered (250 mL) on a day interval basis. Eight weeks after inoculation with nematode eggs, the following data were collected: Number of leaflets per plant, shoot fresh weight, number of galls per root system, gall index per root system, number of egg mass per root system and egg mass index per root system. Root gall/egg mass index was determined using a rating scale according to Taylor and Sasser (1978); 0 = 0, 1 = 1-2, 2 = 3-10, 3 = 11-30, 4 = 31-100 and 5≥100 galls/egg masses per root system. The number of egg mass was determined using Phloxine B (MP Biomedicals, LLC, USA) to stain the roots (0.15 g L-1) for 15 min for easy counting (Daykin and Hussey, 1985).
| Table 1: | Phytochemical composition of leaves of H. suaveolens |
 | |
| +: Low, ++: Moderate, -: Not detectable | |
| Table 2: | Effects of soil amendment with powdered leaves of H. suaveolens on fresh root weight (g) of African yam bean 6 weeks after inoculation with eggs of M. javanica |
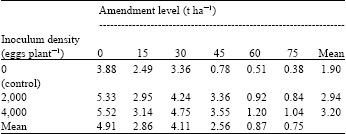 | |
| LSD0.05 for comparing amendment level means = 1.557, LSD0.05 for comparing inoculum density means = 1.101, LSD0.05 for comparing amendment level means x inoculum density means interaction = 2.697 | |
| Table 3: | Effects of soil amendment with powdered leaves of H. suaveolens on fresh shoot weight (g) of African yam bean 6 weeks after inoculation with eggs of M. javanica |
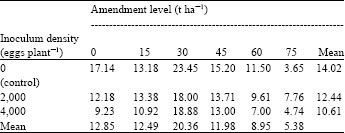 | |
| LSD0.05 for comparing amendment level means = 2.409, LSD0.05 for comparing inoculum density means = NS, LS for comparing amendment level means x inoculum density means interaction = 8.463 | |
| Table 4: | Effects of soil amendment with powdered leaves of H. suaveolens on the number of nodules per root of African yam bean 6 weeks after inoculation with eggs of M. javanica |
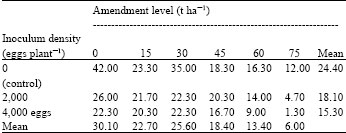 | |
| LSD0.05 for comparing amendment level means = 9.97, LSD0.05 for comparing inoculum density means = 6.90, LSD0.05 for comparing amendment level means x inoculum density means interaction = 16.91 | |
Data recorded were analyzed statistically with Genstat for windows, version 3.2. Treatment means were separated using Least Significant Difference (LSD) at 5% level of probability.
| Table 5: | Effects of varying levels of amendments with powdered leaves of H. suaveolens on the number of gall (NG) and gall index (GI) of African yam bean inoculated with eggs of M. javanica. |
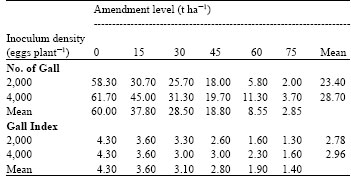 | |
| LSD0.05 for comparing amendment level means (NG) = 17.44, LSD0.05 for comparing inoculum density means (NG) = NS, LSD0.05 for comparing amendment level means (GI) = 0.794, LSD0.05 for comparing inoculum density means (GI) = NS, LSD0.05 for comparing amendment level means x inoculum density means interaction (NG) = 24.66, LSD0.05 for comparing amendment level means x inoculum density means interaction (GI) = 1.123 | |
| Table 6: | Effects of varying levels of amendments with powdered leaves of H. suaveolens on the number of egg mass (NEG) and egg mass index (EMI) of African yam bean inoculated with eggs of M. javanica |
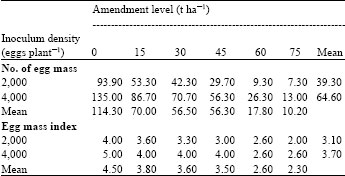 | |
| LSD0.05 for comparing amendment level means (NEG) = 19.84, LSD0.05 for comparing inoculum density means (NEG) = 11.46, LSD0.05 for comparing amendment level means (EMI) = 0.819, LSD0.05 for comparing inoculum density means (EMI) = 0.473, LSD0.05 for comparing amendment level means x inoculum density means interaction (NG) = 28.07, LSD0.05 for comparing amendment level means x inoculum density means interaction (EMI) = 1.158 | |
RESULTS
Results of the qualitative phytochemical screening performed on H. suaveolens leaves showed moderate presence for terpenoids, flavonoids, tannins and alkaloids, low presence for glycosides, resins and volatile oil and absence for saponin (Table 1). Results of the effects of powdered leaves of H. suaveolens on fresh root weight of the African yam bean inoculated with M. javanica showed that fresh root weights were highest at 30 t ha-1 for both infected and uninfected controls (Table 2). Root fresh weight declined progressively with increase in concentration of leaf amendment beyond 30 t ha-1. Root fresh weights were highest in plants infected with 4,000 eggs of M. javanica and lowest in plants without M. javanica infection but with leaf amendment. Results also showed significant differences (p≤0.05) among amendment levels and the inoculums densities differing significantly (p≤0.05) with the uninfected controls.
Results on the effects of powdered leaves of H. suaveolens on fresh shoot weight of African yam bean inoculated with M. javanica are shown in Table 3. It was observed that fresh shoot weights (g) were highest at 30 t ha-1 for both infected and uninfected controls. A steady decline in fresh shoot weight was observed at amendment levels above 30 t ha-1. Mean fresh shoot weight was highest in the uninfected but amended controls while the least mean fresh shoot weight was observed in plants amended and infected with 4,000 eggs of M. javanica. Results also showed significant differences (p≤0.05) among different amendment levels. No significant differences (p≥0.05) were observed between different inoculum densities and uninfected controls.
Results on the effects of powdered leaves of H. suaveolens on the number of nodules per root of African yam bean inoculated with M. javanica are shown in Table 4. Results showed that the highest number of root nodules was produced in control plants without infection and amendments. Generally, the mean number of root nodules in plants with amendment and infection was highest at 30 t ha-1. The mean number of nodules per root of African yam bean reduced progressively with increase in amendment levels above 30 t ha-1. Results showed significant differences (p≤ 0.05) in the number of nodules among different amendment levels.
Meanwhile, the mean number of root nodules in uninfected but amended controls differed significantly (p≤0.05) from the plants amended but infected with 4,000 eggs of M. javanica.
Results on the effects of powdered leaves of H. suaveolens on number of galls and gall index of African yam bean inoculated with M. javanica are shown in Table 5. It showed that the number of galls and gall index decreased with increase in amendment levels. The highest number of galls and gall index were observed at 0 t ha-1 while the least was recorded at 75 t ha-1. Also, plants inoculated with 4,000 eggs of M. javanica produced the highest mean number of galls and gall index while the least was recorded in plants inoculated with 2,000 eggs. It was observed that there were significant differences (p≤0.05) among different amendment levels but not between the inoculum densities for both the number of galls and gall index.
Results on the effects of powdered leaves of H. suaveolens on number of egg masses and egg mass index of African yam bean inoculated with eggs of M. javanica showed that the number of egg masses and egg mass index decreased with increase in amendment levels (Table 6). The highest number of egg masses and egg mass index were observed at 0 t ha-1 while the least was recorded at 75 t ha-1. Also, plants inoculated with 4,000 eggs of M. javanica produced the highest number of egg masses and egg mass index while the least was recorded in plants inoculated with 2,000 eggs of the nematode. It was also observed that there were significant differences (p≤0.05) among different amendment levels and between the different inoculum densities for both the number of egg masses and egg mass index.
DISCUSSION
The presence of certain phytochemicals in the leaves of H.suaveolens reported in this experiment could be responsible for the reduction in the M. javanica population and reproduction on African yam bean. Wang et al. (2012) observed nematicidal activity in their experiment with alkaloids. Chitwood (2002) also reported that terpenoids, alkaloids, steroids and flavonoids are nematicidal. In fact, some phytochemicals from spices have been reported to have inhibitory effect on the glutathione-S-Transferases (which are important in defense systems of nematodes) of M. incognita and as such nematicidal (Babu et al., 2012).
The observed higher fresh shoot weights among uninfected plants may be due to the absence of nematode infection and presence of nutrients supplied as organic soil amendment with leaf powder of H. suaveolens. Nematodes deprive host plants of the necessary nutrients for growth and yield and as such cause impaired growth and development (Olowe, 1981; Babatola and Adalemo, 1988). Results also showed that organic soil amendment with H. suaveolens improved the growth of plants infected with eggs of M. javanica as against the infected plants but unamended. This result is suggestive of a possible bio-nematicidal activity in H. suaveolens used for soil amendment. It was also observed in this study that higher soil amendment levels (>30 t ha-1) led to sharp progressive decline in the fresh shoot and root weights of amended plants. This phenomenon is indicative of a possible phytotoxicity of H. suaveolens leaf powder at amendment level beyond 30 t ha-1. Phytotoxicity in amended plants has been reported by some researchers at different amendment levels and stages of growth of test plants (Kaplan and Noe, 1993; Nwanguma and Fawole, 2004). Udo and Ugwuoke (2010) reported phytotoxicity in turmeric (Curcuma longa L.) using poultry manure soil amendment level beyond 20 t ha-1. Mian and Rodriguez-Kabana (1982) reported that organic amendments with low C: N ratios when applied to the soil at a rate of 1.0% w/w or above are greatly phytotoxic to test plants and nematicidal too. This type of adverse reaction by test plants underscores the need to work out optimal rate of a particular organic amendment for nematode control in a particular crop as there is a possibility of specificity in reactions to different types of amendment. Further work aimed at finding out the actual cause of the phytotoxicity of H. suaveolens amendment to the African yam bean and measures for ameliorating the adverse reaction possibly by combination with another type of organic amendment is thus proposed. It was also observed that the highest nematode inoculums density (4,000 eggs) gave the least fresh shoot weight and this means that the higher the inoculum density of the nematode the higher their infectivity and pathogenicity. Similar result obtained by Udo and Nwagwu (2007) in tomato suggests the need to work out a minimum threshold level of infection for nematodes on test plants in different agroecological regions.
It was also observed that there was an inverse relationship between gall formation by M. javanica and H. suaveolens soil amendment levels. The least number of galls of galls and gall index was recorded at the highest amendment level of H. suaveolens leaf powder. This result is indicative of the inherent nematicidal effects of H. suveolens since the amendment levels restrained M. javanica from formation of galls freely as observed in the unamended but infected plants. Gall formation is diagnostic of root knot nematode infection in most infected plants (Abad et al., 2003). This confirms earlier reports that H. suaveolens has bionematicidal activity (Olabiyi et al., 2006, 2011). Plant materials are known to possess alkaloids and many other phytochemical that could be inimical to many plant parasitic nematodes and when used as soil amendment could release aldehydes, ammonia and other gases that are nematicidal during decomposition (Alam et al., 1979; Badra et al., 1979). It has also been reported that application of organic amendment/fertilizer can improve and promote soil aggregations that retard movement of nematode larvae in the soil (Mbah and Onweremadu, 2009). This is also a pointer to the fact that the bionematicidal compounds in H. suaveolens were absorbed by the test plants and this made the plant tissue toxic for nematode development and reproduction. Reduced formation of galls by amended roots of test plants is a tissue response which suggests that antinematode substances were absorbed by the root tissues (Javed et al., 2007).
Soil amendment with H. suaveolens also affected the number of egg masses and egg mass index adversely. The number of egg masses and egg mass index rapidly decreased as the amendment levels increased. This means that reproduction in M. javanica was seriously disrupted by the amendment and this led to the ultimate reduction in nematode population of the soil. This is in agreement with similar reports where organic soil amendment were applied for the management of plant parasitic nematodes (Saravanapriya and Sivakumar, 2005; Karmani et al., 2011). Onyeke and Akueshi (2012) achieved significant reduction in number of galls and egg masses by M. incognita on African yam bean using Azadirachta indica, Moringa oleifera, Cymbopogon citratus, Nauclea latifolia and Crotolaria retusa leaves separately as soil amendment. Also, Hayat et al. (2012) reported effective prevention of the fecundity of M. incognita and subsequent significant reduction in the number of galls and egg masses on infected potato using organic soil amendments. Ivoke et al. (2009) reported that ethanolic and aqueous leaf extracts of H. suaveolens were both ovicidal and larvicidal against Anopheles gambiae. Another experiment by Nayak et al. (2010) reported significant anthelmintic activity against Pheretinia posthuma and Ascardia galli using both alcoholic and aqueous extracts of whole Hyptis suaveolens plant. Similar effects of the aforementioned experiments could have been expressed in the case of M. incognita eggs and larvae in the present study.
The observed fewer number of nodules in infected plants when compared with the uninfected plants could be attributed to the hyperplasia and hypertrophy (gall formation) taking place in infected roots thereby impeding nodulation activity in such roots. Also, presence of nematodes in the rhizosphere soil can cause competition for space and reduction in root hairs through which infection by nitrogen fixing bacteria can take place (Epps and Chambers, 1962; Malek and Jenkins, 1964). Past reports on reduction of nodulation activities by nematodes in some legumes confirm this result (Huang and Barker, 1983; Varschney et al., 1987; Babatola and Adalemo, 1988). The higher root weights recorded in infected and unamended test plants compared to the uninfected controls could be attributed to giant cell formation triggered by secretions from the salivary gland of root-knot nematode feeding on roots of test plants (Davis and Mitchum, 2005). Galls are nutrient sinks and cause nutrients to be re-directed from plant shoot to roots (Melakeborhan et al., 1990).
CONCLUSION
In conclusion, results from this study have shown the potential of H. suaveolens as a possible biopesticide for the management of M. javanica on African yam bean, S. stenocarpa and its potential phytotoxicity at amendment levels above 30 t ha-1.
ACKNOWLEDGMENT
The authors are grateful to the Department of Plant Science and Biotechnology, University of Nigeria, Nsukka and the International Institute for Tropical Agriculture (IITA), Ibadan, Nigeria for providing the facilities and the African Yam Bean seeds, respectively.
REFERENCES
- Abad, P., B. Favery, M.N. Rosso and P. Castagnone-Sereno, 2003. Root-knot nematode parasitism and host response: Molecular basis of a sophisticated interaction. Mol. Plant Pathol., 4: 217-224.
CrossRefPubMedDirect Link - Agu, C.M., 2008. Effects of organic manure types on root-gall nematode disease and African yam bean yield. Agric. J., 3: 14-16.
Direct Link - Haseeb, A. and F. Butool, 1996. Evaluation of nematicidal properties of some members of the family Solanaceae. Bioresour. Technol., 57: 95-97.
CrossRefDirect Link - Babu, R.O., D. Moorkoth, S. Azeez and S.J. Eapen, 2012. Virtual screening and in vitro assay of potential drug like inhibitors from spices against Glutathione-S-Transferase of Meloidogyne incognita. Bioinformation, 8: 319-325.
CrossRefDirect Link - Chitwood, D.J., 2002. Phytochemical based strategies for nematode control. Annu. Rev. Phytopathol., 40: 221-249.
CrossRefPubMedDirect Link - Davis, E.L. and M.G. Mitchum, 2005. Nematodes. Sophisticated parasites of legumes. Plant Physiol., 137: 1182-1188.
Direct Link - Daykin, M.E. and R.S. Hussey, 1985. Staining and Histopathological Techniques in Nematology. In: An Advanced treatise on Meloidogyne, Barker, K.R., C.C. Carter and J.N. Sasser (Eds.). North Carolina State University, Raleigh, NC., USA., pp: 39-48.
Direct Link - Huang, J.S. and K.R. Barker, 1983. Influence of Heterodera glycines on leghemglobins of soybean nodule. Phytopathology, 73: 1002-1004.
CrossRef - Hussey, R.S. and R.H. Bourma, 1981. A greenhouse screening procedure for root-knot nematode resistance in soybeans. Crop Sci., 21: 794-796.
CrossRefDirect Link - Ivoke, N., F.C. Okafor and L.O. Owoicho, 2009. Evaluation of ovicidal and larvicidal effects of leave extracts of Hyptis suaveolens (L) POIT (Lamiaceae) against Anopheles gambiae (Diptera: Anophelidae) complex. Anim. Res. Int., 6: 1072-1076.
Direct Link - Javed, N., S.R. Gowen, M. Inam-ul-Haq and S.A. Anwar, 2007. Protective and curative effect of neem ( Azadirachta indica) formulations on the development of root-knot nematode Meloidogyne javanica in roots of tomato plants. Crop Protect., 26: 530-534.
CrossRefDirect Link - Kaplan, M. and J.P. Noe, 1993. Effects of chicken-excrement amendments on Meloidogyne arenaria. J. Nematol., 25: 71-77.
Direct Link - Karmani, B.K., M.M. Jiskani, M.I. Khaskheli and K.H. Wagan, 2011. Influence of organic amendments on population and reproduction of root knot nematode, Meloidogyne incognita in egg plants. Pak. J. Agri. Agril. Eng. Vet. Sci., 27: 150-159.
Direct Link - Mbah, C.N. and E.U. Onweremadu, 2009. Effect of organic and mineral fertilizer inputs on soil and maize grain Yield in an Acid Ultisol in Abakaliki-South Eastern Nigeria. Am.-Eurasian J. Agron., 2: 7-12.
Direct Link - Melakeborhan, H., H. Ferris and J.M. Dias, 1990. Physiological response of resistant and sucepitible Vitis vinifera cultivars to Meloidogyne incognita. J. Nematol., 22: 224-230.
Direct Link - Mian, I.H. and R. Rodriguez-Kabana, 1982. Survey of the nematicidal properties of some organic materials available in Alabama as amendments to soil for control of Meloidogyne arenaria. Nematropica, 12: 235-246.
Direct Link - Muazu, A.A. and I. Umar, 2012. Evaluation of some plant materials for control of M. incognita on cowpea (Vigna unguiculata L. Walp). New Clues Sci., 2: 28-41.
Direct Link - Nayak, P.S., S. Nayak, D.M. Kar and P. Das, 2010. In vitro anthelmintic activity of whole plant extracts of Hyptis suaveolens poit. Int. J. Curr. Pharm. Res., 2: 50-51.
Direct Link - Nwanguma, E.I. and B. Fawole, 2004. Efficacy of organic soil amendments on the populations of Meloidogyne incognita on Okra in South-Western Nigeria. Niger. J. Hort. Sci., 9: 89-95.
Direct Link - Ogbuji, R.O., 1981. Pathogenicity of Meloidogyne incognita Race 1 on Lima bean cultivars. Der Tropenlandwirt-J. Agric. Tropics Subtropics, 82: 19-23.
Direct Link - Olabiyi, T.I., I.O. Adepoju , S.A. Abolusoro and E.E.A. Oyedunmade, 2011. Suppression of nematode pests of tomato with aqueous leaf extracts of nitta, tobacco and pawpaw. Am.-Eurasian J. Agron., 4: 23-27.
Direct Link - Onyeke, C.C. and C.O. Akueshi, 2012. Infectivity and reproduction of Meloidogyne incognita (Kofoid and White) Chitwood on African yam bean, Sphenostylis stenocarpa (Hochst Ex. A. Rich) Harms accessions as influenced by botanical soil amendments. Afr. J. Biotechnol., 11: 13095-13103.
Direct Link - Saravanapriya, B. and M. Sivakumar, 2005. Management of root knot nematode, Meloidogyne incognita on tomato with botanicals. Nat. Prod. Radiance, 4: 158-161.
Direct Link - Udo, I.A. and K.I. Ugwuoke, 2010. Pathogenicity of Meloidogyne incognita race 1 on turmeric (Curcuma longa L.) as influenced by inoculum density and poultry manure amendment. Plant Pathol. J., 9: 162-168.
CrossRefDirect Link - Wang, K., C. Luo, H. Liu, J. Xu, W. Sun and L. Zhou, 2012. Nematicidal activity of the alkaloids from Macleaya cordata against certain nematodes. Afr. J. Agric. Res., 7: 5925-5929.
Direct Link