
Pratibha Sharma
Division of Plant Pathology, Indian Agricultural Research Institute, New Delhi, 110012, India
Swati Deep
Division of Plant Pathology, Indian Agricultural Research Institute, New Delhi, 110012, India
Dinesh Singh Bhati
Division of Plant Pathology, Indian Agricultural Research Institute, New Delhi, 110012, India
Manika Sharma
Division of Plant Pathology, Indian Agricultural Research Institute, New Delhi, 110012, India
P. Chowdappa
Indian Institute of Horticultural Research, Bengaluru, 560089, India
Plant Pathology Journal
Year: 2014 | Volume: 13 | Issue: 2 | Page No.: 100-111







ABSTRACT
Alternaria blight caused by Alternaria brassicicola (Schwein) Wiltshire and Alternaria brassicae (Berk) Sacc. is one of the most serious diseases of Cauliflower [Brassica oleracea (L.) var. botrytis] and Mustard [Brassica juncea L.] grown as edible vegetable and oilseed crop, respectively in India. As resistance against these pathogens is not found till now therefore, the exploration of the host-pathogen interaction is much needed. In the present study, the cauliflower-A. brassicicola and mustard-A. brassicae interactions were studied at microscopic level using light and electron microscope. The initial infection processes involve conidia germination, penetration and colonization on the plant surfaces. Several germ-tubes developed from conidia and colonize extensively across the leaf surfaces. Penetration in the plant surface, either directly through the epidermis or via stomata occurred with occasional appressoria formation. Least aggressive A. brassicae isolate was found to infect mustard leaf by spreading over epidermal cells only. Finally, hyphae colonize the leaf cells forming network collapsing epidermal cells to form necrotic lesions which may be due to release of toxin.
PDF Abstract XML References Citation
Received: December 18, 2013;
Accepted: March 17, 2014;
Published: April 19, 2014
How to cite this article
Pratibha Sharma, Swati Deep, Dinesh Singh Bhati, Manika Sharma and P. Chowdappa, 2014. Penetration and Infection Processes of Alternaria brassicicola on Cauliflower Leaf
and Alternaria brassicae on Mustard Leaf: A Histopathological Study. Plant Pathology Journal, 13: 100-111.
DOI: 10.3923/ppj.2014.100.111
URL: https://scialert.net/abstract/?doi=ppj.2014.100.111
DOI: 10.3923/ppj.2014.100.111
URL: https://scialert.net/abstract/?doi=ppj.2014.100.111
INTRODUCTION
Leaf spot is a common term applied to a disease that normally affects the foliage of annual, perennial crops and forest trees, the majority of which are caused by a variety of fungal pathogens and also by bacteria. Among various leaf spot fungal pathogens, Alternaria species is one of the major production constraints in most of the field and horticultural crops. In vegetable Brassica seeds, especially white cabbage and cauliflower [Brassica oleracea (L.) var. botrytis], Alternaria brassicicola (Schwein) Wiltshire (Maude and Humpherson-Jones, 1980; Humpherson-Jones, 1985; Maude et al., 1984; Kubota et al., 2006; Deep and Sharma, 2012; Sharma et al., 2013a) is the dominant pathogen whereas in oilseed rape, especially mustard [Brassica juncea (L.)], A. brassicae (Berk) Sacc. is dominant (Sharma et al., 2013b). These species can survive in seeds for several months at different temperatures and relative humidity (Kumar and Gupta, 1994; Abul-Fazal et al., 1994) and the disease is spread during the growing season having cool and moist weather by wind-blown or rain-splashed spores (Rotem, 1994; MacKinnon et al., 1999; Oliver et al., 2001). The pathogen attacks most parts of the plant like seed, stem, leaf, inflorescence and fruits and it is thought to induce its chlorotic effect by release of phytotoxins (Jung et al., 2002). Alternaria leaf spots usually appear on the oldest leaves first and later spread to the newer leaves towards the tips (Chattopadhyay, 1999; Meena et al., 2010; Deep and Sharma, 2012). Leaf spots appear as bright to pale yellow or tan flecks on the upper leaf surface which may be surrounded by light green or yellow halos. The older spots are somewhat circular to irregularly lobed and are light brown-black in colour. These old spots may or may not have characteristic concentric rings. Lesions on the petioles and stems are dark brown and often coalesce and girdle the stems. The leaf spots increase in size and number and coalesce and cause leaf blights.
The disease cycle is simple as no teleomorph found till now (Aveling et al., 1994). Conidia are produced on leaf surface and dispersed by wind. Conidia germinate in availing suitable condition getting moisture and produce toxin before penetrating host tissue. Penetration on the leaf surface can occur directly or through stomata (Dehpour et al., 2007). To study the penetration and infection process of the A. brassicicola and A. brassicae on economically important cauliflower and mustard leaves, respectively, the present study has been carried out.
MATERIALS AND METHODS
Inoculum preparation: Seven day old Alternaria brassicicola (CaAbcT3 isolate), A. brassicae (CaAbD5, highly aggressive and CaAbR3, least aggressive isolates) grown on Potato Dextrose Agar (PDA) plates at 25°C were flooded with distilled water and spores were released by agitation with a sterile brush. The resulting spore suspension density was adjusted to 4x104 spores mL-1 by haemocytometer. One drop of Tween-20 1 mL-1 suspension was added as a wetting agent.
Plant material: The cauliflower cultivar namely, Pusa Deepali (early-maturing) and mustard cultivar namely, Pusa Jagganath seeds were obtained from Division of Vegetable Sciences and Division of Plant Pathology, Indian Agricultural Research Institute (IARI), New Delhi, respectively. The cauliflower seeds were sown in nursery beds during October, 2011-12 seasons. Seedlings were transplanted after two weeks in fields with a spacing of 30x40 cm. Similarly mustard seeds were also sown in field. 45-60 days old leaves were taken for the study further.
Detached leaf inoculation: Leaves were properly washed under running tap water and then surface wiped off with 70% alcohol and 2 μL of 4x104 spores mL-1 spores were inoculated with a fine needle (Dispovan, 2.5 mL) while sterile distilled water was applied on control (Sharma et al., 2004). The leaves were placed inside moist chambers in green house conditions and were observed for appearance of disease symptoms on 3rd day after inoculation. Development of disease symptoms was observed on third day after inoculation in the inoculated leaves while controls remained free from symptoms.
Light microscopy: To observe the infection process of Alternaria brassicicola and A. brassicae on leaves of cauliflower and mustard, infected and uninfected leaf samples were properly washed with distilled water twice and then dried. Thin section (1”) crosswise out was cut into transparent slices with the help of a sterilized blade or scalpel. Thin cross sections of leaf samples were decolorized with 0.15% (w/v) TCA in Ethanol: Chloroform (3:1) then wet mounted with a drop of 0.01% lactophenol cotton blue on a glass slide and covered with a cover slip. The slides were observed under light microscope (Zeiss AX10Star Plus) at 50-60 Hz and 65 V. More than fifty samples were examined to confirm the results.
Scanning electron microscopy: Samples of leaf pieces (size: ~2 mm) after infection collected were washed in 0.1 M phosphate buffer three times at an interval of 15 min at a temperature of 4°C. Samples were post fixed in 1% Osmium tetraoxide in the same buffer for 2 h at 4°C and again washed in 0.1 M phosphate buffer three times as above. Dehydration with different concentrations of acetone (30, 50, 70, 80, 90, 95% once each and 100% twice) was carried out at 15 min interval at 4°C. Critical point drying (optional) with liquid CO2 (31.5°C at 100 PSI) was done. The dried materials were then adhered on to the aluminium specimen mounts with colloidal silver paste and then sputter coated (Emitech CA7625 Carbon Accessory) with gold palladium (~24 nm thickness). The specimens were examined and photographed on a scanning electron microscope (Zeiss EVOMA10) at 20 kV/EHT and 10 Pa between 2-100 X zoom. At least 30 leaf disks were observed to verify the result.
All the three isolates of A.brassicicola (CaAbcT3) and A. brassicae (CaAbD5 and CaAbR3) grown on PDA, were also observed under light microscope and scanning electron microscope as above to observe the morphology microscopically.
RESULTS AND DISCUSSION
Conidia of Alternaraia brassicicola isolate CaAbcT3 was found to be small and septate ranging from 7-35 μm long with short beak (Fig. 1). Chain of conidia were found (Fig. 1b) having 2-8 conidia forming germ tubes which were branched infrequently with average size of 8-200 μm in length. Similarly conidia of A. brassicae CaAbD5 (Fig. 1c) and CaAbR3 (Fig. 1d) were also found to be small and septate having 3-5 septa and 10-45 μm long with long beak. This was morphologically similar to leaf spot causing pathogen A. brassicae in vegetable and oil seed crops (Sharma et al., 2013c). As a dominant pathogen in cauliflower (Deep and Sharma, 2012; Sharma et al., 2013a), A. brassicicola infected cauliflower leaf tissue revealed damage to cell and cell wall as compared to the healthy tissue. The damaged portions were found to be discoloured (Fig. 2a) as a result of interaction with the fungus. It may be due to production of lytic enzymes like polygalacturonase, pectin lyase, pectin methylestarase, cellulase or fungal toxins either Host Specific Toxin (HST) or Non-Host Specific Toxin (NHST) (Nozaki et al., 1997; Berto et al., 1997; Gautam et al., 2012; Jahangeer et al., 2005). In some infected cell, deposition of lignin to the cell wall was found (Ramm, 1962). A. brassicicola conidia both germinated and ungerminated were adhered strongly to the leaf surface. Light microscopy results showed profusely growing germtubes from the conidia dispersed throughout the infected leaf surface lesion (Fig. 2b).
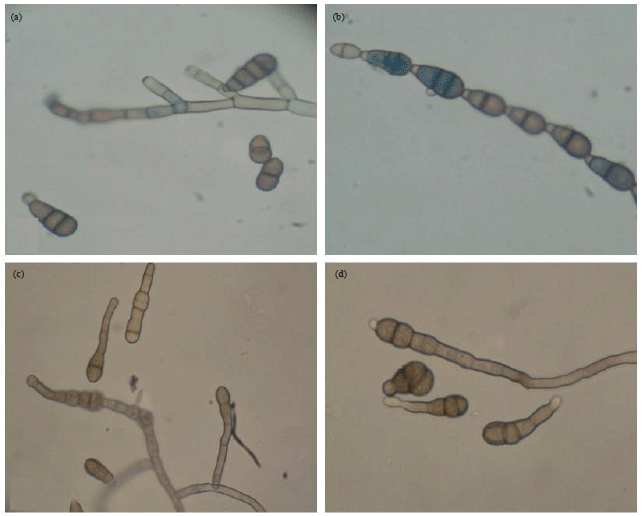 | |
| Fig. 1(a-d): | Light micrograph (40X) of conidia and mycelia formed by Alternaria brassicicola and Alternaria brassicae (a) Mature small beaked conidia of A. brassicicola with multiple septa, (b) A. brassicicola conidia arranged in chain form, (c) Mature conidia of A. brassicae isolate; CaAbD5 and (d) A. brassicae isolate; CaAbR3 with multiple septa and long beak |
Germtube was also found to penetrate the leaf cell through stomata (Fig. 2c, d) forming appresoria. The formation of several germ tubes and appressoria from a conidium is a common feature of this genus, as reported in A. porri (Giri et al., 2013; Aveling et al., 1994; Everts and Lacy, 1996), A. linicola (Vloutoglou et al., 1996), A. alternata (Gupta et al., 1997) and A. cassia (Mims et al., 1997).
Scanning electron microscopy also resulted into similar observations as that of light microscopy. Healthy cauliflower leaf (Fig. 3a) was found to have normal epidermal cells without any damage while A. brassicicola (CaAbcT3) infected leaf surface (Fig. 3b) was spreaded over by the hypahal growth intercellularly entering with or without forming appresoria near and through the stomatal opening. Extensive growth of germ tubes formed a hyphal network on the infected leaf tissue (Fig. 3c). Some hyphal structures of A. brassicae (highly aggressive; CaAbD5) were found to spread around the intercellular space of the mesophyll and parenchyma tissue through the wounds of the mustard leaf (Fig. 3d). Most of the A. brassicicola conidia and germ tube penetrate inside the epidermal cell directly through stomata while other passed near the stomata without forming appresoria (Fig. 4a, b). Similarly young conidia of A. brassicae were found to spread over the mustard leaf tissue (Fig. 4c) and movement of germtube was found through epidermal cells of mustard leaf not through stomata (Fig. 4d).
A distinct difference between highly aggressive and least aggressive isolate of A. brassicae was found over the infection process on mustard leaf. The highly aggressive isolate of A. brassicae (CaAbD5) was found to invade through stomatal opening of the host mustard leaf (Fig. 5a) as that of A. brassicicola infection on cauliflower leaf while the least aggressive isolate of A. brassicae (CaAbR3) was found to infect the mustard leaf by spreading its hyphae over the epidermal cells moving adjascent to stomata (Fig. 5b).
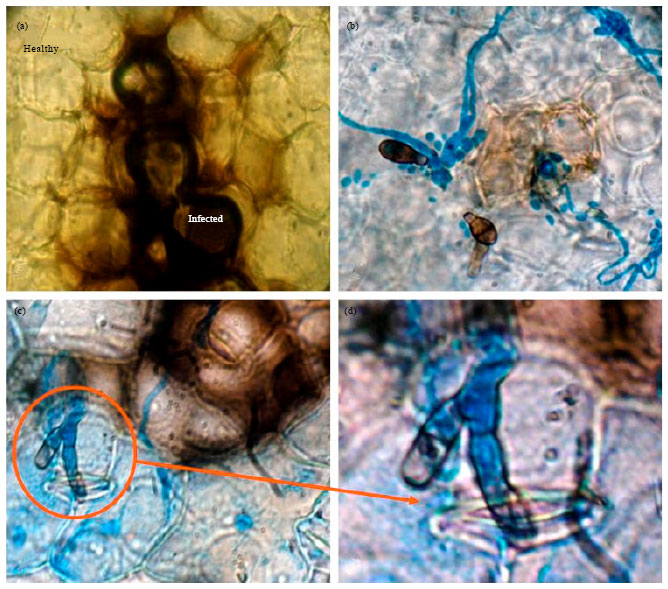 | |
| Fig. 2(a-d): | Light micrograph of cauliflower leaf infected with Alternaria brassicicola (40X) (a) Cauliflower leaf tissue showing difference between healthy and infected portion due to Alternaria toxin, (b) A. brassicicola conidia and mycelia spread on cauliflower leaf cells and (c, d) Germtube of A. brassicicola penetrating the cauliflower leaf tissue through stomata opening |
The microscopic infection process of A. brassicicola on cauliflower and A. brassicae on mustard observed in the present study was similar to that of infection by wild type A. brassicicola on cabbage leaf surface where appresorium was developed from the conidia to penetrate plant surface through stomata and wounds to colonize its hyphae (Scott, 2008). Another microscopic observations also revealed that the fungus, A. brassicicola penetrate Arabidopsis thaliana siliques through cellular junctions, replum and stomata and into seed coats either directly or through cracks (Pochon et al., 2012). Giri et al. (2013) also found similar mode of infection and penetration by A. brassicae on mustard leaf tissue. Similar results were found by some other Alternaria species on a range of hosts (Dehpour et al., 2007; Isshiki et al., 2001; Kiely, 1964; Mims et al., 1997; Pegg, 1966; Ruehle, 1937; Saad and Hagedorn, 1969). Van Dyke and Trigino (1987) reported that the cells in the substomatal area beneath appressoria were necrotic with no evidence of fungal invasion in the tissue. Hyphal penetration was seldom observed prior to necrosis of mesophyll cells and that the death of these cells in advance of fungal penetration the action of diffusible toxins.
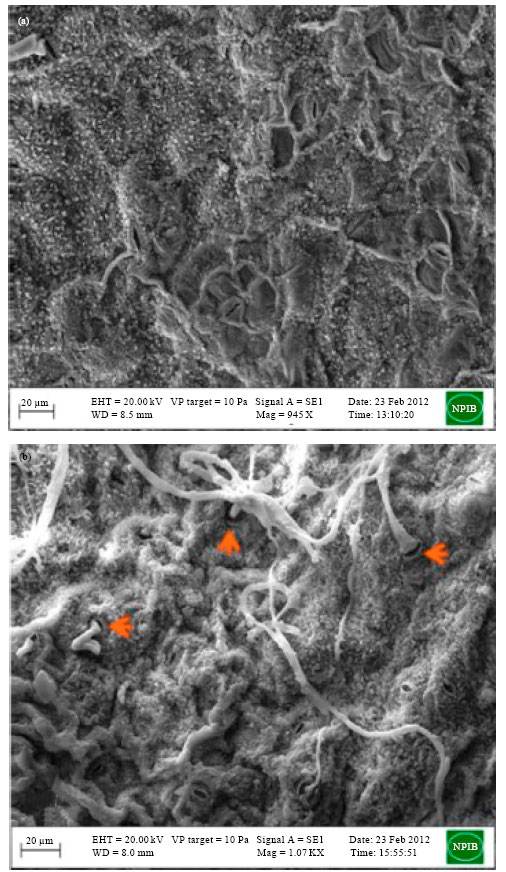  | |
| Fig. 3(a-d): | Scanning electron micrograph of cauliflower and mustard leaves infected with Alternaria brassicicola and Alternaria brassicae (a) Healthy cauliflower leaf (Bar = 20 µm). (b) Infected cauliflower leaf with movement of A. brassicicola mycelia through stomatal opening, (c) Spreading of hyphal network of A. brassicicola (CaAbcT3) on infected cauliflower leaf surface and (d) A. brassicae (Highly aggressive; CaAbD5) hyphae growing intercellularly passing through damaged epidermal cells of the necrotic lession |
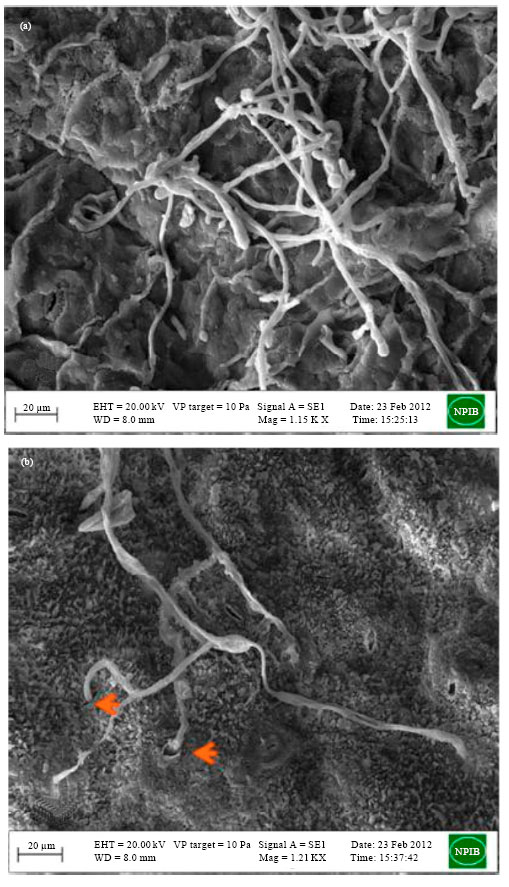  | |
| Fig. 4(a-d): | Scanning electron micrograph of penetration of conidia and germtube of Alternaria brassicicola and Alternaria brassicae on cauliflower and mustard leaf, respectively, (a, b) Direct penetration of A. brassicicola (CaAbcT3) hyphae through the epidermis and stomata (Bar = 20 µm), (c) Spreading of A. brassicae (CaAbD5) hyphae and conidia over infected mustard leaf (Bar = 20 µm) and (d) Movement of A. brassicae (CaAbD5) hyphae and germtube through epidermal cells of mustard leaf not through stomata (Bar = 10 µm) |
 | |
| Fig. 5(a-b): | Scanning electron micrograph of movement of germtube and hypahe of Alternaria brassicae on mustard (Pusa Jagganath) leaf (a) Penetration of germtube of A. brassicae (highly aggressive; CaAbD5) through stomatal opening and (b) Spreading of A. brassicae (least aggressive; CaAbR3) hyphae over the epidermal cells of leaf, passing near the stomata (Bar = 10 µm) |
Dehpour et al. (2007) found that secondary hyphae developed from primary hyphae and grew in the intercellular spaces and also penetrated and grew intercellularly with in the epidermis.
Both A. brassicicola and A. brassicae enter the leaf tissue of B. oleracea var. botrytis and B. juncea, respectively both by direct (epidermal) and stomatal penetration which was confirmed by same penetrations process in A. ricini on castor leaf and A. porri on onion leaf (McKenzie et al., 1993; Aveling et al., 1994; Suheri and Price, 2000; Babu et al., 2009). The least aggressive A. brassicae isolate showed somewhat deviation in the present study by moving only over epidermal cells of B. juncea. Similar results were found by A. linicola on water agar, linseed leaves (Vloutoglou et al., 1996) and in A. alternate on mulberry leaves (Gupta et al., 1997) whereas, penetration only through stomata was observed in case of A. eichhorniae on water hyacinth leaves (Shabana et al., 1997) and in A. alternata on grape fruits (Swart et al., 1995).
CONCLUSION
In the present study, only the process of infection and penetration of A. brassicicola through cauliflower leaf tissue and A. brassicae through mustard leaf tissue were studied and confirmed with previous studies done on interaction of other Alternaria spp. on wide range of hosts. Penetration and infection directly by epidermal cella and/or stomatal opening was found. Hyphae of the both the Alternaria spp. on both Brassica host was found to spread like net formation which lead to damage the leaf cell forming necrotic lesions. This is initial study of infection process of the important leaf spot causing pathogen in crucifer. The mechanism behind the host and pathogen interactions can be studied further.
ACKNOWLEDGMENTS
Director, IARI, Head, Plant Pathology and Financial support by Indian Council of Agricultural Research (ICAR), through Outreach program entitled “Diagnosis and management of leaf spot diseases of field and horticultural crops” is duly acknowledged.
REFERENCES
- Abul-Fazal, M., M.I. Khan and S.K. Saxena, 1994. The incidence of Alternaria species ID different cultivars of cabbage and cauliflower seeds. Indian Phytopathol., 47: 419-421.
Direct Link - Aveling, T.A.S., H.G. Snyman and F.H.J. Rijkenberg, 1994. Morphology of infection of onion leaves by Alternaria porri. Can. J. Bot., 72: 1164-1170.
CrossRefDirect Link - Babu, A.M., T. Philip, V. Kumar and B.K. Kariappa, 2009. Germination, penetration and sporulation of Alternaria ricini (Yoshii) Hansf. on castor leaf. Arch. Phytopathol. Plant Protect., 42: 915-921.
CrossRef - Berto, P., L. Belingheri and B. Dehorter, 1997. Production and purification of a novel extracellular lipase from Alternaria brassicicola. Biotechnol. Lett., 19: 533-536.
CrossRefDirect Link - Chattopadhyay, C., 1999. Yield loss attributable to Alternaria blight of sunflower (Helianthus annuus L.) in India and some potentially effective control measures. Int. J. Pest Manage., 45: 15-21.
CrossRef - Deep, S. and P. Sharma, 2012. Host age as predisposing factor for incidence of black leaf spot of cauliflower caused by Alternaria brassicae and Alternaria brassicicola. Indian Phytopathol., 65: 71-75.
Direct Link - Dehpour, A.A., S.V. Alavi and A. Majad, 2007. Light and scanning electron microscopy studies on the penetration and infection processes of Alternaria alternata, causing brown spot on Minneola Tangelo in the West Mazandaran-Iran. World Applied Sci. J., 2: 68-72.
Direct Link - Everts, K.L. and M.L. Lacy, 1996. Factors influencing infection of onion leaves by Alternaria porri and subsequent lesion expansion. Plant Dis., 80: 276-280.
Direct Link - Gautam, S.P., P.S. Bundela, A.K. Pandey, Jamaluddin, M.K. Awasthi and S. Sarsaiya, 2012. Diversity of cellulolytic microbes and the biodegradation of municipal solid waste by a potential strain. Int. J. Microbiol.
CrossRef - Giri, P., G. Taj, P.D. Meena and A. Kumar, 2013. Microscopic study of Alternaria brassicae infection processes in Brassica juncea cultivars by drop plus agarose method. Afr. J. Microbiol. Res., 7: 4284-4290.
Direct Link - Gupta, V.P., H.V. Raju, V. Kumar and Govindaiah, 1997. Surface ultrastructure of infection process of Alternaria alternata and Fusarium pallidoroseum on mulberry. Arch. Phytopathol. Plant Protect., 31: 429-434.
CrossRef - Humpherson-Jones, F.M., 1985. The incidence of Alternaria spp. and Leptosphaeria maculans in commercial brassica seed in the United Kingdom. Plant Pathol., 34: 385-390.
CrossRefDirect Link - Isshiki, A., M. Akimitsu and H. Yamamoto, 2001. Endopolygalacturonase is essential for citrus black rot caused by Alternaria citri but not brown spot caused by Alternaria alternate. Mol. Plant Microb. Interact., 14: 749-757.
Direct Link - Jahangeer, S., N. Khan, S. Jahangeer, M. Sohail, S. Shahzad, A. Ahmad and S.A. Khan, 2005. Screening and characterization of fungal cellulases isolated from the native environmental source. Pak. J. Bot., 37: 739-748.
Direct Link - Jung, D.S., Y.J. Na and K.H. Ryu, 2002. Phyolgenic analysis of Alternaria brassicicola producing bioactive metabolites. J. Microbiol., 40: 289-294.
Direct Link - Kubota, M., K. Abiko, Y. Yanagisawa and K. Nishi, 2006. Frequency of Alternaria brassicicola in commercial cabbage seeds in Japan. J. Gen. Plant Pathol., 72: 197-204.
CrossRefDirect Link - Kumar, R. and P.P. Gupta, 1994. Survival of Alternaria brassicae, A. brassicicola and A. alternata in the seeds of mustard (B. juncea) at different temperatures and relative humidities. Ann. Biol., 10: 55-58.
Direct Link - MacKinnon, S.L., P. Keifer and W.A. Ayer, 1999. Components from the phytotoxic extract of Alternaria brassicicola, a black spot pathogen of canola. Phytochemistry, 51: 215-221.
CrossRefDirect Link - Maude, R.B. and F.M. Humpherson-Jones, 1980. Studies on the seed-borne phases of dark leaf spot Alternaria brassicicola and grey leaf spot Alternaria brassicae of brassicas. Ann. Applied Biol., 95: 311-319.
CrossRefDirect Link - Maude, R.B., F.M. Humpherson-Jones and C.G. Shuring, 1984. Treatments to control Phoma and Alternaria infections of brassica seeds. Plant Pathol., 33: 525-535.
CrossRefDirect Link - McKenzie, C.L., B. Cartwright, M.E. Miller and J.V. Edelson, 1993. Injury to onions by Thrips tabaci (Thysanoptera: Thripidae) and its role in the development of purple blotch. Environ. Entomol., 22: 1266-1277.
Direct Link - Meena, P.D., R.P. Awasthi, C. Chattopadhyay, S.J. Kolte and A. Kumar, 2010. Alternaria blight: A chronic disease in rapeseed-mustard. J. Oilseed Brassica, 1: 1-11.
Direct Link - Mims, C.W., M.A. Rogers and C.G. Van Dyke, 1997. Ultrastructure of conidia and conidium germination in the plant pathogenic fungus Alternaria cassiae. Can. J. Bot., 75: 252-260.
CrossRef - Oliver, E.J., P.H. Thrall, J.J. Burdon and J.E. Ash, 2001. Vertical disease transmission in the Cakile-Alternaria host-pathogen interaction. Aust. J. Bot., 49: 561-569.
CrossRefDirect Link - Pochon, S., E. Terrasson, T. Guillemette, B. Iacomi-Vasilescu and S. Georgeault et al., 2012. The Arabidopsis thaliana-Alternaria brassicicola pathosystem: A model interaction for investigating seed transmission of necrotrophic fungi. Plant Methods, Vol. 8.
CrossRef - Rotem, J., 1994. The Genus Alternaria, Biology, Epidemiology and Pathogenicity. APS Press, USA., Pages: 326.
Direct Link - Shabana, Y.M., Z.A.A. Baka and G.M. Abdel-Fattah, 1997. Alternaria eichhorniae, a biological control agent for waterhyacinth: Mycoherbicidal formulation and physiological and ultrastructural host responses. Eur. J. Plant Pathol., 103: 99-111.
CrossRef - Sharma, P., S.R. Sharma and M. Sindhu, 2004. A detached leaf technique for evaluation of resistance in cabbage and cauliflower against three major pathogens. Indian Phytopathol., 57: 315-318.
Direct Link - Sharma, M., S. Deep, D.S. Bhati, P. Chowdappa, R. Selvamani and P. Sharma, 2013. Morphological, cultural, pathogenic and molecular studies of Alternaria brassicae infecting cauliflower and mustard in India. Afr. J. Microbiol. Res., 7: 3351-3363.
Direct Link - Sharma, P., S. Deep, M. Sharma and D.S. Bhati, 2013. Genetic variation of Alternaria brassicae (Berk.) Sacc., causal agent of dark leaf spot of cauliflower and mustard in India. J. Gen. Plant Pathol., 79: 41-45.
CrossRef - Sharma, P., S. Deep, M. Sharma, D. Singh and P. Chowdappa, 2013. PCR based assay for the detection of Alternaria brassicicola in Crucifers. Indian Phytopathol., 66: 263-268.
Direct Link - Suheri, H. and T.V. Price, 2000. Infection of onion leaves by Alternaria porri and Stemphylium vesicarium and disease development in controlled environments. Plant Pathol., 49: 375-382.
CrossRef - Swart, A.E., C.L. Lennox and G. Holz, 1995. Infection of table grape bunches by Alternaria alternata. S. Afr. J. Enol. Viticult., 16: 3-6.
Direct Link - Van Dyke, C.G. and R.N. Trigiano, 1987. Light and scanning electron microscopy of the interaction of the biocontrol fungus Alternaria cassiae with sicklepod (Cassia obtusifolia). Can. J. Plant Pathol., 9: 230-235.
CrossRef - Vloutoglou, I., B.D.L. Fitt and J.A. Lucas, 1996. Germination of Alternaria linicola conidia on linseed: Effects of temperature, incubation time, leaf wetness and light regime. Plant Pathol., 45: 529-539.
CrossRef - Ramm, C.V., 1962. Histological studies of infection by Alternaria longipes on tobacco. J. Phytopathol., 45: 391-398.
CrossRef