
Balasubramanian Mythili Gnanamangai
Department of Biotechnology, K.S. Rangasamy College of Technology, Tiruchengode, Tamil Nadu, 637 215, India
Ponnusamy Ponmurugan
Department of Biotechnology, K.S. Rangasamy College of Technology, Tiruchengode, Tamil Nadu, 637 215, India
Rajendran Yazhini
Department of Biotechnology, Karunya University, Coimbatore, Tamil Nadu, 641 114, India
Sekar K. Pragadeesh
Department of Biotechnology, Arunai Engineering College, Tiruvannamalai, Tamil Nadu, 606 603, India
Plant Pathology Journal
Year: 2011 | Volume: 10 | Issue: 1 | Page No.: 13-21







ABSTRACT
A total of three isolates of Cercospora theae namely KC10, MC24 and VC38 obtained from birds eye spot disease were used for the present study and subjected to various Pathogen Related (PR) enzymatic secretion by the pathogen and the plant as a response to the disease attack. The enzyme secreted from the pathogen includes CMCase, amylase, invertase, pectinase and protease, its activity was studied at various incubation periods ranging from 15, 30, 60, 90 and 120 min. All enzymes except amylase, exhibited its maximal activity at 30 min incubation period whereas amylase exhibited its optimal activity at 60 min of incubation. The study revealed that C. theae was capable of producing the range of pathogenic enzymes and exhibited its maximal activity at 10th day harvest. The said enzyme activities was also studied in healthy and C. theae infected tea leaves including chitinase, DNase, Phenylalanine Ammonia Lyase (PAL), Tyrosine Ammonia Lyase (TAL), polyphenol oxidase (PPO), peroxidase (PO) and catalase in order to detect the level of defensive response. The significance of the specific enzymatic activity detected in C. theae isolates and tea leaves were discussed by correlating to the plant pathogenesis. This study showed that phyopathogenic isolates of C. theae is capable of producing various carbohydrases, pectinases and protease and defensive enzymes along with phenolic and oxidative enzymes. The significance of correlating the enzymatic activity with the virulence of the isolates was ascertained and discussed. The enzyme profile studies of the pathogenic isolates assist in understanding and revealing of host-pathogen interaction.
PDF Abstract XML References Citation
Received: February 04, 2011;
Accepted: February 24, 2011;
Published: May 06, 2011
How to cite this article
Balasubramanian Mythili Gnanamangai, Ponnusamy Ponmurugan, Rajendran Yazhini and Sekar K. Pragadeesh, 2011. PR Enzyme Activities of Cercospora theae Causing Bird’s Eye Spot Disease in Tea Plants (Camellia sinensis (L.) O.kuntze). Plant Pathology Journal, 10: 13-21.
DOI: 10.3923/ppj.2011.13.21
URL: https://scialert.net/abstract/?doi=ppj.2011.13.21
DOI: 10.3923/ppj.2011.13.21
URL: https://scialert.net/abstract/?doi=ppj.2011.13.21
INTRODUCTION
Plant pathogens produce a wide range of enzymes that are capable of degrading plant cell wall components during plant pathogenesis these enzymes are called as pathogenic enzymes (PR) and are mostly cellulolytic and pectinolytic (Valueva and Mosolov, 2004). Extracellular proteins secreted by the fungal pathogen are able to macerate the tissues and degrade cell wall components at the time of host-pathogen interaction (Federici et al., 2006). Proteins induced in the plant by pathogen attack are defensive compounds that are elicited due to hypersensitive response of the plant immune system (Falahian et al., 2007). When the compound is an enzyme it is called as defensive enzymes.
Tea is the most popular and inexpensive beverage produced from the young leaves of commercially cultivated tea plant. Tea has become one of an important revenue source for tea producing countries in the world (Bandyopadhyay, 2011). Tea is grown in more than 50 countries. India with around 4,40,000 ha under tea cultivation is the largest producer and consumer of tea in the world (Agnihothrudu, 1999). Tea production is greatly hindered due to a number of pests and diseases. Perennial habit of tea plant, peculiar culture conditions and warm humid climate of the tea growing areas are highly conducive for disease development (Ponmurugan and Baby, 2007). Majority of the diseases is of fungal origin (Muraleedharan and Chen, 1997). In this aspect, fungal attack is very important in terms of direct crop loss as tea dhool is manufactured from leaves. Among the fungal diseases, bird’s eye spot disease is very important and it is caused by Cercospora theae Petch, an Ascomycetes that attacks the younger leaves initially, later confined to mature leaves and some extent to bare stalk also which in turn affect the yield potential (Arulpragasam, 1992).
It forms typical necrotic spots of brown to black colour with a red colored margin upon the infected leaves. The leaf spot disease of tea plant is by far the most serious disease of tea creating a huge capital loss. Distinct relationship between the extent of the disease incidence and heavy crop loss due to bird’s eye spot disease is being recorded every year by UPASI Tea Research Foundation, Valparai, Pollachi District, Tamil Nadu, India (Muraleedharan, 2007).
The enzymes produced by the phytopathogen are reported to be present in the host plant tissues. Plant pathogen causing necrotic symptoms like leaf spots, blotches, canker etc., is due to the secretion of cell wall degrading enzymes. During fungal infection, ranges of enzymes are secreted to help promoting host colonization. Hence, this work deals with the enzymatic profile of isolates of Cercospora theae and its activity in the infected tea plants comparing to that of in healthy plants along with some defense related enzymes (chitinase, protease and DNase), phenolic enzymes (Phenylalanine Ammonia Lyase (PAL) and Tyrosine Ammonia Lyase (TAL)) and oxidative enzymes (peroxidase (PO), polyphenol oxidase (PPO) and catalase).
MATERIALS AND METHODS
Cercospora theae isolates: Cercospora theae isolates namely KC10, MC24 and VC38 used for the present study were obtained from infected tea plants of Koppa, Munnar and Valparai estates belongs to Karnataka, Kerala and Tamil Nadu states respectively of Southern India during June 2010. The isolates were sub-cultured periodically on Potato Dextrose Agar (PDA) and three-day-old culture was used through out the commenced study conducted from August 2010, in assistance and the facilities provided by of K.S. Rangasamy College of Technology, Tiruchengode, Namakkal District, Tamil Nadu.
Preparation of enzyme extract from the pathogen: The isolates were cultivated in Czapeck Dox (CD) medium by replacing the indigenous source of the medium and were harvested at 5th, 10th and 15th day intervals. In order to elicit the secretion of specific enzymes, related substrates like 2% of carboxmethyl cellulose (CMC), starch, sucrose and pectin were replaced for carbon source in the CD medium for inducing CMCase, amylase, invertase and pectinase. In case of enzyme preparation for protease, sodium nitrate in CD medium was replaced with 1% casein. The mycelium was harvested twice using Whatmann No: 1 filtration, the filtrate was subjected to centrifugation at 12000 rpm for 5 min to obtain spore free enzyme extract. The supernatant was then filter sterilized using Rivera vacuum filtration unit model no MV3500 with 0.2 μm pore filter membrane. The filtered extract was then used as the respective enzyme source.
Preparation of enzyme extract from the healthy and infected leaves: The healthy and infected tea leaves harvested from the widely used moderately susceptible south Indian clone UPASI 9, as we concluded from earlier study (Gnanamangai and Ponmurugan, 2010) were collected and ground separately with chilled 0.1 N phosphate buffer in 1:1 ratio. The pulp was removed carefully. The leaf extract was filtered using Whatman No: 1 filter paper and the filtrate were centrifuged at 2000 rpm for 30 min to remove the cellular debris. The supernatant was then filter sterilized and used as the respective enzyme source.
Enzyme extract for assay of phenolic and oxidative enzymes: Healthy and infected leaves collected separately were cut into 1 cm2 and subjected to acetone powder preparation (Neish, 1961) for phenolic enzyme assay. Acetone powder (0.5 g) of each healthy and infected leaves were suspended in tris HCl buffer (0.05M), pH8.8, stirred for 30 min and filtered through cheese cloth. The filtrate was centrifuged at 5000 rpm for 10 min; the resulted supernatant was used as the enzyme source for the assay of Phenylalanine Ammonia Lyase (PAL) and TAL. On the other hand for the assay of oxidative enzymes, enzyme sources used were prepared from the freshly harvested leaves (Kar and Mishra, 1976).
Carbohydrases: The CMCase or β-1-4-endoglucanase activity was determined for all the three isolates of C. theae. According to Zaldivar et al. (2001), using carboxy methyl cellulose as substrate, the formation of reducing sugars was measured by reaction with dinitrosalicyclic acid (DNS). The reaction mixture was prepared as per (Okunowo et al., 2010) and the activity was determined at various time intervals ranging from 15, 30, 60, 90 and 120 min. Absorbance was read at 540 nm using (U-2900E, Model No. 2JI-0003, UV-visible spectrophotometer, Hitachi, Tokyo, Japan). Release of reducing sugar concentration was calculated using glucose as the standard.
Similarly, amylase and invertase activities were determined using starch and sucrose as their substrate respectively and absorbance was measured at 540 nm for estimation of reducing sugars according to Narayanan and Shanmugasundaram (1967). The pectinase activity was estimated using D-Polygalacturonic acid as the standard for the enzyme source obtained from the pathogen (Miller, 1959).
Protease: The protease activity was assayed by micromole release of aromatic residues with Folin phenol reagent, at the absorbance of 650 nm according to the methods of Srivastava and Prasad (1989) and Lowry et al. (1951).
Defense related enzymes: In the same way, all the above enzyme activities were also estimated in the enzymes obtained from healthy and diseased tea leaves. The chitinase activity (Miller, 1959) was determined in enzyme source obtained from healthy and infected leaves using colloidal chitin as the substrate prepared using the methods of Berger and Reynolds (1958). Chitinase was not detected in pathogen due to the fact that chitin is its own cell wall component.
The DNase activity was measured in enzyme extracts obtained from healthy and infected leaves. Calf thymus DNA (0.05%) in phosphate buffer (0.05 M) pH6.5 was used as substrate. The enzyme-substrate reaction mixture was added in the ratio of 1:2 along with 2 mL of 0.2% MgCl2. Absorbance was measured every 30 min by stopping the reaction with 0.5 mL of 2.5% chilled perchloric acid at 260 nm.
Phenolic enzymes (PAL and TAL): Phenylalanine Ammonia Lyase (PAL) and Tyrosine Ammonia Lyase (TAL) activities were measured using cinnamic acid and p-coumaric acid as their respective standards (Koukol and Conn, 1961), the enzyme activity for PAL was measured at 268 nm and for TAL it was measured at 330 nm.
Oxidative enzymes: PO and PPO were assayed by using the method reported by Patra and Mishra (1979). Catalase activities in healthy and infected leaves were assayed using the modified method of Kar and Mishra (1976).
Enzyme activity: Enzyme activity for carbohydrases was measured using micromole of sugar released per min against glucose as a standard. One unit activity is the amount of enzyme releasing 1 μmole of reducing sugar from substrate (CMC, Starch, Sucrose, Pectin and Chitin) where 1 Kat is expressed as 6.107 enzyme unit/mL/min. Enzyme activity for DNase was measured by increasing in 1.0 absorbance per hour as one unit of DNase. Enzyme activity for phenolic enzymes is defined, as the one unit activity is the amount of enzymes (PAL or TAL) released 1 μmole of simple compound (Cinnamic acid or p-Coumaric acid) from the substrate (Phenylalanine or Tyrosine). One unit of peroxidase or polyphenol oxidase activity was defined as the amount of purpurogallin formed which increased the absorbance by 0.1 min-1 under assay condition. One unit of catalase activity is defined as the enzyme which breaks down one μmole of H2O2 min-1 under the assay conditions. Zero time control was used for the entire enzyme assay performed throughout this study.
Specific activity of enzymes: All the enzyme assays was carried out at different incubation time intervals ranging from 15, 30, 60, 90 and 120 min so as to find the optimal incubation period of the specific enzyme and only the activity of the enzyme at its optimal incubation period was denoted in the present study. The specific activity of all the enzymes at the optimal time period by using all three isolates of C. theae along with healthy and disease infected tea leaves were determined and expressed as Kat mg-1. It is the ratio of enzyme unit to the protein content of the enzyme solution.
Statistical analysis: All the grouped data were statistically evaluated using SPSSS 14.0 statistical package (SPSS, Inc. Chicago, IL). Hypothesis testing methods included are one way analysis of variance (ANOVA) followed by Least Significant Difference (LSD) test (Snedecor and Cochran, 1980). Differences with p<0.05 were considered to be statistically significant (Snedecor and Cochran, 1980). All the results were expressed as the Mean±SD for six replicated values.
RESULTS
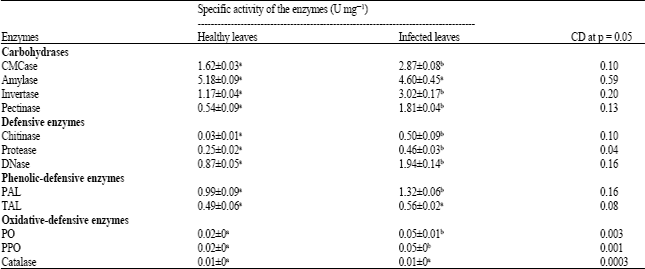
Determination of carbohydrases in C. theae isolates: The results of CMCase activity showed that all the isolates exhibited its activity during 5th, 10th and 15th day of enzyme harvest. The maximal activity was obtained at 10th day enzyme harvest as shown in Table 2 which exhibited significant variation at (p<0.05). In healthy and infected leaves, CMCase activity was determined and found to be high in infected leaves at (p<0.05) (Fig. 1) and exhibits significant variation on comparing with healthy leaves.
Isolates of C. theae exhibiting amylase activity at various intervals of enzyme harvest were determined. The amylase activity was high at p<0.05 on 10th day enzyme harvest (Fig. 1).
Invertase activity determined from the C. theae isolates was maximal at 10th day enzyme harvest. This was the only enzyme that exhibited significantly high enzyme activity at p<0.05 as compared to other enzymes secreted by the C. theae (Table 2). Invertase activity was found to be two fold increase in leaf spot infected leaves (Fig. 1), indicating that invertase may be imparted to leaves due to infection. Carbohydrate metabolism in leaves is altered due to invasion of pathogen.
| Table 1: | Activity of Pathogen related enzymes in healthy and infected tea leaves |
 | |
| Similar alphabets followed in a row denote they are statistically insignificant and vice versa at p<0.05 | |
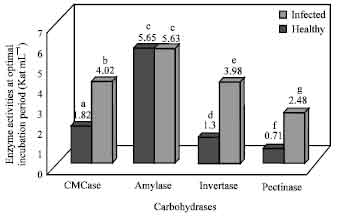 | |
| Fig. 1: | Carbohydrases activities in healthy and C. theae infected tea leaves. Enzyme activity between healthy and infected tissues followed by same alphabetical letters are not statistically significant and followed by different alphabetical letters are statistically significant at p<0.05 |
Pectinase activity in this study was determined and exhibited the maximal activity at the 10th day enzyme harvest compared to 5th and 15th days of harvest (Table 2). Very low pectolytic activity was detected in healthy plants when compared to that of disease infected leaves. Pectinase activity determined in healthy and infected tea leaves is depicted in Fig. 1 which revealed significantly high at p<0.05 in infected leaves.
Determination of chitinase activity: Chitinase is a major defense related protein found to be expressed at higher levels in the susceptible cultivars as a result of infection. The activity of chitinase was more in infected leaves which might be due to infection in order to break the major component of fungal cell wall, the chitin. Chitinase induced from host tea plants showed greater activity with several fold and was observed to be significantly higher in leaf spot affected tissues due to pathogen infection when compared to the enzyme elicitation by healthy tissues (Table 1).
Determination of protease activity: Table 2 shows that the enzyme activity was significantly high at p<0.05 on 10th day enzyme harvest compared to 5th and 15th day. The protease was found to be more in infected leaves comparing to that of the healthy leaves and the differences were significatly high at p<0.05 (Table 1).
Activity of DNase: The healthy and infected leaves examined for DNase activity exhibited two the fold increase in infected tissues to that of healthy tissues (Table 1). Increased level of DNA results in higher DNase activity which may be due to pathogenesis in host plants.
Activity of phenolic defensive enzymes: C. theae infected tissues exhibited an increase in PAL activity with respect to healthy tissues. Significantly higher activities of PAL were observed in leaf samples obtained from infected plants (Fig. 2, Table 1). The present findings confirm the reports on involvement of PAL in the incompatible reactions. TAL was observed to be more in infected tissues compared to that of the activities exhibited by healthy tissues. The difference in TAL activities among the healthy and infected tissues is not significant as it is a subsidiary enzyme in the phenylpropanoid pathway that forms a phenolic precursor.
Oxidative defensive enzyme activities: In the present study, activities of all oxidative enzymes such as peroxidase, polyphenol oxidase and catalase increased to two fold when compared to the activities of healthy tissues (Fig. 3).
| Table 2: | Effect on specific activity of enzymes from C. theae isolates at various intervals |
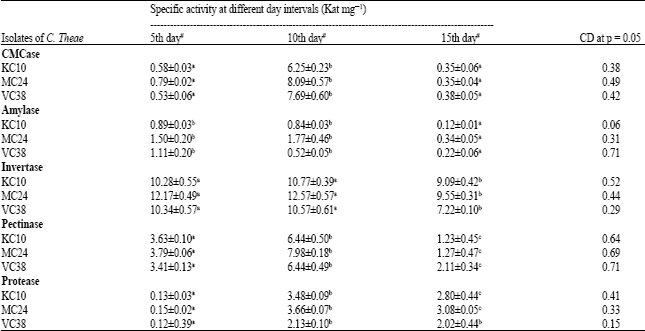 | |
| KC10, MC24 and VC38 are the isolates of C. theae. #Specific activities determined by the isolates were at its optimal time of incubation. Similar alphabets followed in a row denote the differences are statistically insignificant and vice versa at p<0.05 | |
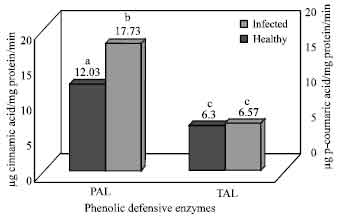 | |
| Fig. 2: | Activity of phenolic defensive enzymes in healthy and infected tea leaves at optimal incubation period. Differences exhibited by each enzyme activity between healthy and infected tissues followed by same alphabetical letters are not statistically significant and followed by different alphabetical letters are statistically significant at p<0.05 |
The observed differences were significantly higher in diseased tissues. Necrotization of the plant cells in C. theae infected tea leaves may have been one of the primary inducers of the oxidative enzymes in the present study.
Specific activity of the enzymes exhibited by C. theae isolates, healthy and infected leaves: The specific activity of CMCase, amylase, invertase, pectinase and protease exhibited by the isolates of C. theae at three different days of harvest was shown in Table 2.
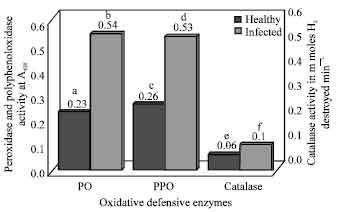 | |
| Fig. 3: | Activity of oxidative defensive enzymes in healthy and infected tea leaves. Differences exhibited by each enzyme activity between healthy and infected tissues followed by same alphabetical letters are not statistically significant and followed by different alphabetical letters are statistically significant at p<0.05 |
CMCase, pectinase and protease exhibited significantly high specific activity at p<0.05 in 10th day enzyme source. Invertase and amylase exhibited high activity at p<0.05 in both 5th and 10th day harvest. The specific activity of all the enzymes was determined for the enzyme activity obtained at its optimal incubation time.
Infected leaves exhibited high specific activity at p<0.05 for invertase, pectinase, chitinase, protease, cellulase and Dnase on comparing with enzymes from healthy leaves. On the other hand amylase was the only enzyme exhibiting high specific activity and catalase was the only enzyme showing very low specific activity in healthy and infected leaves which doesn’t show any significant difference at p<0.05 (Table 1).
DISCUSSION
Studies of Banerjee (1990), Nitharwal et al. (1991) and Moreira et al. (2005) have shown that most of the phytopathogen like Curvularia sp., Fusarium solani and Myrothecium sp. are capable of producing CMCase suggesting CMC as the substrate for the induction of the enzymes at higher levels that induce pathogenesis to plants by penetration of plant material. In well agreement to these studies, C. theae was able to grow in carboxy methyl cellulose indicating that cellulolytic enzymes were secreted by this phytopathogen to depolymerize the carbon sources to simple sugars for its growth.
The activity of the CMCase on comparing between healthy and infected leaves were found to be significantly high which were contrast to work of Moreira et al. (2005) in which plant proteinases had been implicated in the inhibition of CMCase, other cellulases and hemicellulase production by a phtopathogens.
The obtained results for amylase activity were in agreement to the study of Singh et al. (2009) who reported that the activity of amylase was irrespective to the cultivars and stage of infection in leaf blight disease caused Horedeum vulgare in barley plants, indicating that amylase had no significant role in the mechanism of pathogenesis. Similar results were observed in the present study on determining the amylase activity in healthy and infected leaves which doesn’t exhibit any statistically significant variation.
Interesting results on observation of invertase activity were reported, by Mandels (1956) who reported relationship of invertase synthesis in relation to growth of the fungus Myrothecium verrucaria which is due to the surface location of the enzyme in the spores of the pathogen. Increase in invertase activities in response to fungal infection has earlier been reported in Arabidopsis and in wheat leaves by Fotopoulos et al. (2003) and Sutton et al. (2007). It has been reported that both defense and invertase gene expressions were induced by pathogen attack in tomato (Berger et al., 2004). The upregulation of invertase activity appears to be a common response to various biotic and abiotic stresses (Roitsch et al., 2003). They further indicated that an increase in extracellular invertase activity leads to inverse regulation of photosynthesis and carbohydrates during pathogen infection. Role for invertase in the generation of hexoses which supply energy for defense reactions and/or may act as signals inducing defense gene expression was well portrayed by Singh et al. (2009). In general, during leaf infection, invertase activity was predominant. Present results are consistent with the above stated works.
Significant correlation was well demonstrated earlier between pathogenicity causing severe tissue maceration and the extracellular pectinase activities of the indigenous Sclerotinia sclerotiorum isolates from Jordan valley (Asoufi et al., 2007). Obi and Umezurike (1981) reported the elaboration of a pectic enzyme activity by Burkholderia (Pseudomonas) cepacia in diseased onions. In addition, such enzymes are reported to be produced by a variety of phytopathogens (Doyle et al., 2001). Pectic enzymes that have the ability to split the 1, 4 bonds between the galacturonic acid moieties in the pectic fraction of the cell wall remain the only enzyme confirmed to cause plant tissue maceration.
The above results on pectinase activity in C. theae were in agreement with Heale and Gupta (1972) and Bahkali et al. (1997) who reported similar findings in the in vitro and in vivo of some fungi causing leaf spot and soft rot diseases like Fusarium oxysporum, Rizhoctonia Solani, A. macrospora and P. debaryanum.
The lytic enzymes like chitinases reported in the present study were suggested to have hydrolytic action and hence degrades the cell wall of many pathogenic fungi (Mathivanan et al., 1998). Chitinase are known to be induced in many plants in responses to infection with fungal pathogens and were correlated to induced resistance by Ji and Kuc (1996).
Activity of extracellular pumAe protease from U. maydis by growing in YNB medium containing ammonium sulfate showed specific activity that began to increase after 12 h of growth and reached its maximum activity after 24 h was reported by Mercado-Flores et al. (2003). Proteolytic enzymes have a potential importance in plant pathogenesis by disrupting the host plant metabolism and initiating disease process. Some proteases are important in the formation of spores, germination of spores, in pathogenesis of several microorganisms and in post-translational regulation were reported by Rendueles and Wolf (1988) and Monod et al. (1998). In agreement to the above studies our results showed higher protease activity by the pathogen and diseased plants.
Cheema et al. (1981) recorded similar observations on DNase activity in tristeza infected leaves of Kagzilime. Similarly, Verma and Singh (1994) reported decreased levels of DNA and RNA in mango leaves infected by Pestalopsis mangiferae due to increase in nuclease activity that were compared over healthy leaves of mango cultivars.
Significantly high PAL activity in the incompatible interactions was also demonstrated in woody kiwifruit vines (Reglinski et al., 2001) and tea plants (Chakraborty and Chakraborty, 2005). The activity of TAL is not well demonstrated in dicots (Reglinski et al., 2001). TAL activity was definitely lower than PAL in the present findings similar to the results obtained in wheat tissues by Guerra et al. (1985). Many studies have demonstrated the importance of phenolic compounds in plant defense mechanism. In general, plant phenolics have a diverse range of biological activity, depending on their structure, degree of polymerization, stereoisomeric differences, etc. Given all the above properties, it is obvious that plant phenolics play an important role in defense. In the present study, increase in phenolic enzyme activity was observed during host-parasite interaction disease attack which in turn affects the contents of defensive phenolics.
Similar to the present report on oxidative enzymes, previous works also indicated that these oxidative enzymes are involved in the defense role (Chakraborty et al., 2002) with resistance in many plants (Chitra and Rajamani, 2010). An immediate response of plants to injury, in most cases, is the accelerated accumulation of oxidative enzymes required for scavenging toxic radicals (Kessler and Baldwin, 2002). Plants were reported to possess efficient antioxidant defense system due to the presence of pathway for catalase and peroxidase (Saffar et al., 2009). On the other hand, Bi et al. (1997) reported insect pest damage in cotton cultivars caused activation of several oxidative enzymes, including peroxidase, diamine oxidase and lipoxygenase within the tissue. Results of the present study strongly support the above observation of Bi et al. (1997).
Chatterjee and Ghosh (2008) studied the biochemical changes in mesta plants infected with yellow vein disease. It has been reported that different enzymes like catalase, acid phosphatase, peroxidase, esterase, polyphenol oxidase and superoxide dismutase which revealed lower activities of catalase, acid phosphatase and peroxidase enzymes, enhanced activities of esterase, polyphenol oxidase and superoxide dismutase in diseased plants as compared to healthy ones.
CONCLSION
Thus in the present work we have shown a wide range of enzymes secreted by C. theae that has the tendency to depolymerase the important component involved in structural integrity of cell wall. This study also outlines about the significance of enzymatic changes occurred in the tea host as a defensive role, during the infection caused by bird’s eye spot disease. It can be concluded from the present study that, enzymes like invertase, pectinase, CMCase followed by protase are involved in inducing pathogenesis and DNase PAL, chitinase, protease followed by PO, PPO, are involved in imparting defense in tea plants against C. theae during infection. The work has led to the knowledge and growth conditions of this disease causing fungus and the level of the enzyme responses by the healthy and susceptible tea plant that assisted in understanding of host pathogen interaction.
ACKNOWLEDGMENT
The authors wish to express their sincere thanks to the Chairman and Principal, K.S.R. College of Technology, Tiruchengode for their support and constant encouragement.
REFERENCES
- Asoufi, H., K.M. Hameed and A. Mahasneh, 2007. The cellulase and pectinase activities associated with the virulence of indigenous Sclerotinia sclerotiorum Isolates in Jordan Valley. Plant Pathol. J., 23: 233-238.
Direct Link - Bahkali, A.H., A.S. Al-Khaliel and K.A. Elkhider, 1997. In vitro and in vivo production of pectolytic enzymes by some phytopathogenic fungi isolated from Southwest Saudi Arabia. J. King Saud Univ., 9: 125-137.
Direct Link - Bandyopadhyay, T., 2011. Molecular marker technology in genetic improvement of tea. Int. J. Plant Breed. Genet., 5: 23-33.
CrossRefDirect Link - Banerjee, U.C., 1990. Production of β-glucosidase (Cellobiase) by Curvularia sp. Lett. Applied Microbiol., 10: 197-199.
CrossRef - Berger, L.R. and D.M. Reynolds, 1958. The chitinase system of a strain Streptomyces griseus. Biochem. Biophys. Acta, 29: 522-534.
CrossRef - Bi, J.L., J.B. Murphy and G.W. Felton, 1997. Antinutritive and oxidative components of induced resistance in cotton to Helicoverpa zea. J. Chem. Ecol., 23: 97-118.
CrossRef - Chakraborty, B.N., S. Dutta and U. Chakraborty, 2002. Biochemical responses of tea plants induced by foliar infection with Exobasidium vexans. Indian Phytopathol., 55: 8-13.
Direct Link - Chakraborty, U. and N. Chakraborty, 2005. Impact of environmental factors on infestation of tea leaves by Helopeltis theivora and associated changes in flavonoid flavor components and enzyme activities. Phytoparasitica, 33: 88-96.
CrossRef - Chatterjee, A. and S.K. Ghosh, 2008. Alterations in biochemical components in mesta plants infected with yellow vein mosaic disease. Braz. J. Plant Physiol., 20: 267-275.
CrossRefDirect Link - Falahian, F., Z.O. Ardebili, F. Fahimi and R. Khavarinejad, 2007. Effect of mycorrhizal fungi on some defense enzymes against Gaeumannomyces gaminis in wheat. Pak. J. Biol. Sci., 10: 2418-2422.
CrossRefPubMedDirect Link - Mercado-Flores, Y., C. Hernandez-Rodriguez, J. Ruiz-Herrera and L. Villa-Tanaca, 2003. Proteinases and exopeptidases from the phytopathogenic fungus Ustilago maydis. Mycologia, 95: 327-339.
Direct Link - Fotopoulos, V., M.J. Gilbert, J.K. Pittman, A.C. Marvier and A.J. Buchanan et al., 2003. The monosaccharide transporter gene, AtSTP4 and the cell-wall invertase, at beta fruct1, are induced in Arabidopsis during infection with the fungal biotroph Erysiphe cichoracearum. Plant Physiol., 132: 821-829.
CrossRef - Guerra, D., A.J. Anderson and F.B. Salisbury, 1985. Reduced phenylalanine ammonia lyase and tyrosine ammonia lyase activities and liginin synthesis in wheat grown under low pressure sodium lamps. Plant Physiol., 78: 126-130.
Direct Link - Heale, J.B. and D.P. Gupta, 1972. Mechanism of vascular wilting induced by Verticillium albontram. Trans. Br. My. Soc., 58: 19-28.
CrossRef - Ji, C. and J. Kuc, 1996. Antifungal activity of cucumber β 1-3 glucanase and chitinase. Physiol. Mol. Plant Pathol., 49: 257-265.
Direct Link - Kar, M. and D. Mishra, 1976. Catalase, peroxidase and polyphenoloxidase activities during rice leaf senescence. Plant Physiol., 57: 315-319.
CrossRefDirect Link - Kessler, A. and I.T. Baldwin, 2002. Plant responses to insect herbivory: The emerging molecular analysis. Annu. Rev. Plant Biol., 53: 299-328.
CrossRefDirect Link - Koukol, J. and E.E. Conn, 1961. The metabolism of aromatic compounds in higher plants IV Purification and properties of the phenylalanine deaminase of Hordeum vulgare. J. Biol. Chem., 236: 2692-2698.
Direct Link - Lowry, O.H., N.J. Rosebrough, A.L. Farr and R.J. Randall, 1951. Protein measurement with the folin phenol reagent. J. Biol. Chem., 193: 265-275.
CrossRefPubMedDirect Link - Mandels, G.R., 1956. Synthesis and secretion of invertase in relation to the growth of Myrothecium verrucaria. J. Bacteriol., 71: 684-688.
Direct Link - Mathivanan, N., V. Kabilan and K. Murugesan, 1998. Purification, characterization and antifungal activity of chitinase from Fusarium chlamydosporum, a mycoparasiteto groundnut rust, Puccinia arachidis. Can. J. Microbiol., 44: 646-651.
CrossRefPubMedDirect Link - Miller, G.L., 1959. Use of dinitrosalicylic acid reagent for determination of reducing sugar. Anal. Chem., 31: 426-428.
CrossRefDirect Link - Monod, M., B. Hube, D. Hess and D. Sanglard, 1998. Differential regulation of SAP8 and SAP9, which encode two new members of the secreted aspartic proteinases family in Candida albicans. Microbiology, 144: 2731-2737.
PubMed - Moreira, F.G., S. dos Reis, M.A.F. Costa, C.G.M. de Souza and R.M. Peralta, 2005. Production of hydrolytic enzymes by the plant pathogenic fungus Myrothecium verrucaria in submerged cultures. Brazil. J. Microbiol., 36: 7-11.
CrossRefDirect Link - Muraleedharan, N. and Z.M. Chen, 1997. Pests and diseases of tea and their management. J. Plantn. Crops, 25: 15-43.
Direct Link - Narayanan, A.S. and E.R.B. Shanmugasundaram, 1967. Studies on amylase of Fusarium vasinfectum. Arch. Biochem. Biophys., 118: 317-322.
CrossRef - Neish, A.C., 1961. Formation of m-coumaric and p-coumaric acids by enzymatic deamination of the corresponding isomers of tyrosine. Phytochemistry, 1: 1-24.
CrossRef - Nitharwal, P.D., H.N. Gour and S. Agarwal, 1991. Effects of different factors on the production of cellulose by Curvularia lunata. Folia Microbiol., 36: 357-361.
CrossRef - Obi, S.K.C. and G.M. Umezurike, 1981. Pectic enzyme activities of bacteria associated with rotted onions (Allium cepa). Applide Environ. Microbiol., 42: 585-589.
Direct Link - Okunowo, W.O., G.O. Gbenle, A.A. Osuntoki, A.A. Adekunle and S.A. Ojokuku, 2010. Production of cellulolytic and xylanolytic enzymes by a phytopathogenic Myrothecium roridum and some a virulent fungal isolates from water hyacinth. Afr. J. Biotechnol., 9: 1074-1078.
Direct Link - Patra, H.K. and D. Mishra, 1979. Pyrophosphatase, peroxidase and polyphenoloxidase activities during leaf development and senescence. Plant Physiol., 63: 318-323.
Direct Link - Ponmurugan, P. and U.I. Baby, 2007. Morphological, physiological and biochemical changes in resistant and susceptible cultivars of tea in relation to phomopsis disease. Plant Pathol. J., 6: 91-94.
CrossRefDirect Link - Reglinski, T., G. Whitaker, J.M. Cooney, J.T. Taylor, P.R. Poole, P.B. Roberts and K.K. Kim, 2001. Systemic acquired resistance to Sclerotinia sclerotiorum in kiwifruit vines. Physiol. Mol. Plant. Pathol., 58: 111-118.
CrossRefDirect Link - Rendueles, P.S. and D.H. Wolf, 1988. Proteinase function in yeast: Biochemical and genetic approaches to a central mechanism of post-translational control in the eukaryotic cell. FEMS Microbiol. Lett., 54: 17-45.
CrossRef - Roitsch, T., M.E. Balibrea, M. Hofmann, R. Proels and A.K. Sinha, 2003. Extracellular invertase: Key metabolic enzyme and PR protein. J. Exp. Bot., 54: 513-524.
CrossRefDirect Link - Sutton, P.N., M.J. Gilbert, M.J. Williams and. J.L. Hall, 2007. Powdery mildew infection of wheat leaves changes host solute transport and invertase activity. Physiol. Plant, 129: 787-795.
Direct Link - Valueva, T.A. and V.V. Mosolov, 2004. Role of inhibitors of proteolytic enzymes in plant defense against phytopathogenic microorganisms. Biochem. (Moscow), 69: 1305-1309.
PubMedDirect Link - Zaldivar, M., J.C. Velasquez, I. Contreras and L.M. Perez, 2001. Trichoderma aureoviride 7-121, a mutant with enhanced production of lytic enzymes: Its potential use in waste cellulose degradation and/or biocontrol. Electron. J. Biotechnol., 4: 1-9.
Direct Link - Saffar, A., M.B. Bagherieh Najjar and M. Mianabadi, 2009. Activity of antioxidant enzymes in response to cadmium in Arabidopsis thaliana. J. Biol. Sci., 9: 44-50.
CrossRefDirect Link - Chitra, R. and K. Rajamani, 2010. Assessment of different glory lily (Gloriosa superba L.) accessions for biochemical traits. Res. J. Med. Plant, 4: 21-25.
CrossRefDirect Link