
S.F. Hwang
Crop Diversification Centre North, Alberta Agriculture and Rural Development, Edmonton, AB, T5Y 6H3, Canada
H.U. Ahmed
Crop Diversification Centre North, Alberta Agriculture and Rural Development, Edmonton, AB, T5Y 6H3, Canada
K. Ampong-Nyarko
Crop Diversification Centre North, Alberta Agriculture and Rural Development, Edmonton, AB, T5Y 6H3, Canada
S.E. Strelkov
Department of Agricultural, Food and Nutritional Science, University of Alberta, Edmonton, AB, T6G 2P5, Canada
R.J. Howard
Crop Diversification Centre South, Alberta Agriculture and Rural Development, Brooks, AB, T1R 1E6, Canada
G.D. Turnbull
Crop Diversification Centre North, Alberta Agriculture and Rural Development, Edmonton, AB, T5Y 6H3, Canada
Plant Pathology Journal
Year: 2009 | Volume: 8 | Issue: 3 | Page No.: 120-126







ABSTRACT
Rhodiola (Rhodiola rosea) is a plant with adaptogenic properties and is suitable for cultivation in Alberta, Canada. Disease surveys indicated the occurrence of root rots in rhodiola plantations in the Province. A total of 74 fungal isolates were associated with discoloration and rotting in the crown and root regions of the plants. Among these, 15 isolates were identified as Fusarium sp., three as Pythium sp. and eight as Rhizoctonia sp. This is the first report of root rot in rhodiola in Alberta, Canada. These soil-borne pathogens are the potential threat to the quality and quantity of rhodiola production. Experiments were conducted to determine the effect of vesicular-arbuscular mycorrhizal fungi and these soil pathogens on rhodiola growth and development under greenhouse conditions. Overall results indicated that Fusarium, Pythium and Rhizoctonia sp. are all capable of reducing rhodiola biomass. However, biomass was significantly higher when vesicular-arbuscular mycorrhizal fungi were applied in conjunction with these pathogens or in non-inoculated controls. This suggests that vesicular-arbuscular mycorrhizal fungi could be used as a management tool for the control of seedling root rot diseases of rhodiola.
PDF Abstract XML References Citation
How to cite this article
S.F. Hwang, H.U. Ahmed, K. Ampong-Nyarko, S.E. Strelkov, R.J. Howard and G.D. Turnbull, 2009. Causal Agents of Root Rot and the Effect of Vesicular-Arbuscular Mycorrhizal Fungi in Seedlings of Rhodiola rosea in Alberta, Canada. Plant Pathology Journal, 8: 120-126.
DOI: 10.3923/ppj.2009.120.126
URL: https://scialert.net/abstract/?doi=ppj.2009.120.126
DOI: 10.3923/ppj.2009.120.126
URL: https://scialert.net/abstract/?doi=ppj.2009.120.126
INTRODUCTION
Rhodiola rosea L. (roseroot, golden root or arctic root) is a yellow-flowered herbaceous perennial plant of the Crassulaceae family. The plant is a circumpolar species of cool temperate and sub-arctic areas of the Northern hemisphere, including North America, Greenland, Iceland and the Altai, Tien-Shan and Himalaya mountains in Asia. The European distribution includes Scandinavia and most of the mountains of Central Europe.
Rhodiola is a multipurpose medicinal herb with adaptogenic properties that increase the body’s nonspecific resistance and normalize body functions (Brown et al., 2002; Galambosi, 2005).
Based on the documented beneficial pharmacological effects and product safety, commercial interest in roseroot-based products has increased worldwide. Commercial development, however, has been hindered by insufficient quantities of rhodiola to meet the growing industrial demand. Rhodiola cultivation is the only solution to this lack of raw materials for the production of roseroot-based products. Rhodiola cultivation experiments are being conducted at various locations in Alberta, since the province has an environment that is ideal for the growth of this crop (Ampong-Nyarko, 2004). To cultivate rhodiola, the seedlings need to be established in a greenhouse and transplanted to the field. Therefore, the crop requires a 4-5 year commitment of time, land and labour before it reaches harvestable maturity.
As with other plant species, R. rosea is susceptible to attack by various pathogenic microorganisms. However, there have been few plant pathological studies on rhodiola (Hwang et al., 2007) because, until recently, rhodiola has not been extensively grown as a crop. Losses due to disease may be a critical factor in adoption of the crop by Alberta producers. Root rot has caused particular concern because the crop derives its economic value from its roots. Since, rhodiola needs a long time from planting to harvesting, soil-borne pathogens are of prime importance and monitoring their prevalence and identification of the pathogens are necessary for successful disease management.
Vesicular-Arbuscular Mycorrhizal (VAM) fungi are reported to be effective biological control agents for the control of many soil-borne fungal root diseases, including those caused by Fusarium, Pythium and Rhizoctonia sp. (Chang et al., 2006; Hwang, 1988; Hwang et al., 1992; Yao et al., 2002). Since, no research results are available on the interaction of VAM, root rot pathogens and rhodiola, the objectives of this study were to: (1) isolate and identify mycoflora associated with discoloured and diseased rhodiola roots, (2) evaluate the effect of root-associated pathogens on rhodiola seedlings and (3) understand the influence of VAM and root pathogens on rhodiola seedlings.
MATERIALS AND METHODS
Collection of diseased plant samples and fungal isolation: Plantations of rhodiola were surveyed for the presence of diseased plants in the greenhouse and experimental plots at Crop Diversification Center (CDC) North, Edmonton, Alberta and in farmers’ fields in the spring of 2005, 2006 and 2007. The plots at each site were checked for the presence of diseased or dead plants. A total of 50 rhodiola seedlings showing disease lesions on the roots were collected from several locations in central Alberta. Roots and stem bases bearing distinct lesions were washed thoroughly in running tap water and cut into 2x2 mm pieces. Ten pieces of discoloured root tissue from each of the 50 plant samples were surface-disinfested in 0.6% sodium hypochlorite (v/v) for 1 min, rinsed in sterile distilled water and plated on pentachloronitrobenzene (PCNB) medium (Nash and Snyder, 1962) and potato dextrose agar acidified with lactic acid to pH 5.0 (APDA). After seven to 10 days, hyphal tips of representative isolates (based on colony characteristics) were transferred onto fresh Potato Dextrose Agar (PDA) tubes. Fusarium isolates were transferred from the PDA slants onto Carnation Leaf Agar (CLA) (Leslie and Summerell, 2006) and PDA (BD/Difco, Sparks, MD) for identification. The isolates were incubated at room temperature under fluorescent light with a 16/8 h photoperiod. Observations of the cultural and microscopic characteristics of the isolates were recorded 10 to 14 days after plating. Isolates were identified using the taxonomic key of Nelson et al. (1983) and the additional species descriptions of Leslie and Summerell (2006). Pythium isolates were identified to the genus level, by colony morphology on APDA and Rhizoctonia isolates were identified to the species level based on colony morphology on PDA and anastomosis grouping as described below.
Anastomosis grouping of Rhizoctonia isolates: Anastomosis grouping of Rhizoctonia isolates was conducted using tester isolates representing AG-1, AG-2-1, AG-2-2, AG-3, AG-4, AG-5, AG-6, AG-7, AG-8 and AG-9 of R. solani. Tests were conducted on a clean slide in Petri dishes, which served as moisture chambers. Slides were dipped into 70% ethanol, flamed and placed on top of two pieces of water agar. Agar plugs of the tester strains and the isolate to be tested were placed on the slide, 2 cm apart. Five milliliters of sterilized distilled water were added to each dish to provide high humidity. At least two slides were prepared for each isolate. Petri dishes were incubated at 25°C and growth was monitored every 24 h. When hyphae of the two isolates touched, slides were removed from the Petri dishes and examined under a microscope to determine whether anastomosis had occurred.
Effect of Fusarium, Pythium and Rhizoctonia inoculum on rhodiola
Rhodiola seedlings and growth conditions: Rhodiola seedlings were grown in a greenhouse on Metro-Mix® (Sun-Gro Horticulture, Vancouver, BC) in 96-cell (3x3 cm) plastic trays (ITML Horticultural Products, Brantford, ON). Surface-sterilized rhodiola seeds were sown and covered with a thin layer of the soil mixture. The seedlings were grown for 30 days and then transplanted into 450 mL plastic cups (Georgia-Pacific Canada Ltd., Edmonton, AB) filled with pasteurized Metro-mix, at a rate of three seedlings per cup during inoculation. The experiment was conducted under greenhouse conditions at 20±5°C temperature and a 16/8 h photoperiod.
Inoculum preparation: Mycelial inoculum of 15 Fusarium isolates, three Pythium isolates and eight Rhizoctonia isolates was prepared in 250 mL Erlenmeyer flasks containing 100 mL of sterile Potato Dextrose (PD) broth. The PD broth was inoculated with a 9 mm diameter mycelial agar disc from a 7 day old culture and shaken continuously at 200 rpm for seven days at room temperature (20°C) under ambient light. The culture in each flask was comminuted for 15 sec in a waring blender. For each isolate, 20 mL of the inoculum was poured into pots containing a steam-pasteurized soil mixture during seedling transplanting. Non-inoculated PD broth was used as a control. Three separate experiments were conducted for the three pathogens. For each experiment ten replicate pots were used for each treatment (isolate) in every repetition of the experiments. The experiments were repeated once. All pots in an experiment were randomly arranged on a greenhouse bench in a Complete Randomized Design (CRD). Two months after inoculation data on plant height, shoot vigour and shoot dry mass were recorded. Shoot vigour was assessed subjectively on a scale of 1-4 where, 1 = poor, 2 = fair, 3 = fairly good and 4 = good. Re-isolations were made from the roots of diseased plants to confirm the pathogenicity of the isolates.
Data analysis: Prior to analysis of variance, data were tested for homogeneity using a normal probability plot. The data distributions for the three pathogens were normal. Analysis of Variance (ANOVA) of data was performed following the General Linear Model procedure using SAS Statistical software (SAS Institute Inc., 2005). The trialxtreatment (isolate) interaction was not significant for any of the pathogens at p≤0.05. Therefore, the data were pooled over the trials before analysis. The means separation was performed using Duncan’s new multiple range test (p≤0.05).
Influence of VAM and root pathogens on rhodiola seedlings: To test the interaction of Fusarium avenaceum (isolate RD18), Pythium (isolate RD50) and Rhizoctonia solani (RD12) (one isolate of each pathogen) with VAM (Glomus intraradices) (Myke Pro®; Premier Tech Biotechnologies, Rivière-du-Loup QC), three separate experiments (one for each pathogen) were conducted under greenhouse conditions. This VAM strain has been found to provide effective control of Fusarium, Pythium and Rhizoctonia root disease in other crops (Chang et al., 2006; Hwang, 1988; Hwang et al., 1992). The rhodiola plants used for these experiments were grown and transplanted as described above. The treatments in each experiment included: (1) pathogen+VAM, (2) no pathogen +VAM, (3) pathogen+no VAM and (4) no pathogen+no VAM. The treatment that included pathogen and VAM was inoculated with 5 mL cup-1 of each. The pathogen was grown on sterilized wheat grains for 15 days and then dried at 29°C and ground to powder. The VAM or pathogen inoculum was applied in a 5 mL volume to the top of the soil when the seedlings were growing in the 450 mL plastic cups, as described above. Two months after inoculation, the plants were harvested, the roots washed under tap water and data on shoot dry mass, root dry mass and shoot length and shoot dry mass were recorded and analyzed. Shoot vigour was assessed following the same scale (1-4) as described above.
Data analysis: Exploratory analysis using a normal probability plot indicated heterogeneity in the original distribution of the three sets of data obtained with the three pathogens to determine their effect on rhodiola. The data were square root transformed to provide the best fit for a normal distribution. Analysis of variance (ANOVA) was performed following the General Linear Model (SAS Institute Inc., 2005) procedure. The class variables included trial, treatment and trial x treatment interactions; the response variables included plant number, shoot length, shoot dry mass and root dry mass. Since, there were significant trialxtreatment interactions (although not for all response variables) in the data sets of the three pathogens, the data were analyzed separately by trial. The means separation was performed using Fisher’s Protected Least Significant Difference (LSD) at p≤0.05.
RESULTS
Isolation and identification of root-associated mycoflora: A total of 74 fungal isolates were isolated from the roots and crowns of plant samples exhibiting necrosis or discoloration. Among the isolates, 15 were identified as Fusarium sp., three were Pythium sp. and eight isolates were Rhizoctonia solani. The remaining isolates were not identified and not included in the study. Among the Fusarium isolates, three were classified as F. avenaceum, two as F. culmorum and two as F. equiseti (Fig. 1a-c), whilst the other Fusarium isolates could not be identified to the species level because of an absence of identifying characteristics. Anastomosis testing indicated that all eight R. solani isolates belonged to AG-4. The Pythium isolates were not identified at the species level.
Effect of Fusarium, Rhizoctonia and Pythium on rhodiola: Analysis of variance indicated that plant height, shoot vigour and shoot dry mass were significantly reduced in inoculated plants relative to the non-inoculated control. There also were differences among individual isolates of Fusarium and Rhizoctonia (Table 1, 2), but no significant differences were observed among Pythium isolates (Table 3). In the case of seedlings inoculated with Fusarium sp., plant heights ranged from 1.8-3.3 cm, shoot vigour from 1.5-2.8 and shoot dry mass from 0.19 -0.57 g, two months after inoculation (Table 1). In the case of R. solani-inoculated seedlings, the plant heights ranged from 1.9-3.3 cm, shoot vigour from 1.5-2.5 and shoot dry mass from 0.15-0.42 g (Table 2). In the case of seedlings inoculated with Pythium, plant height ranged from 2.0-2.4 cm, shoot vigour from 1.5-1.8 and shoot dry mass from 0.21-0.33 g (Table 3).
Response of VAM and root pathogens on rhodiola seedlings: With few exceptions, there were significant differences among the variables for all pathogens in the trial (p≤0.05).
 | |
| Fig. 1: | Fusarium sp. associated with Rhodiola rosea root rot in Alberta. (a), Fusarium avenaceum, (b) Fusarium culmorum and (c) Fusarium equiseti). Scale bar = 25 μm |
| Table 1: | Effect of Fusarium sp. on rhodiola height, shoot vigour and shoot dry mass when grown under greenhouse conditions |
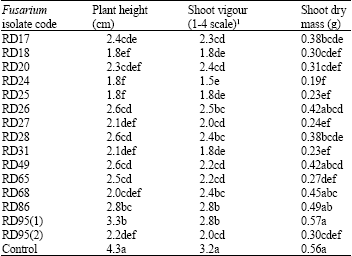 | |
| Data are the means of 10 replications. Means followed by the same letter(s) are not significantly different according to Duncan’s new multiple range test (p≤0.05). 1Vigour was assessed using 1-4 scale, where, 1 = Poor; 2 = Fair; 3 = Fairly good and 4 = Good growth of shoots and roots | |
| Table 2: | Effect of Rhizoctonia solani on rhodiola height, shoot vigour and shoot dry mass when grown under greenhouse conditions |
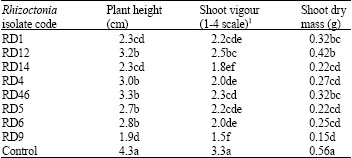 | |
| Data are the means of 10 replications. Means followed by the same letter(s) are not significantly different according to Duncan’s new multiple range test (p≤0.05). 1Vigour was assessed using 1-4 scale, where, 1 = Poor; 2 = Fair; 3 = Fairly good and 4 = Good growth of shoots and roots | |
In the case of seedlings inoculated with F. avenaceum, the shoot dry mass was significantly higher in the control (no pathogen; no VAM) followed by the treatment that included both the pathogen and VAM (Table 4).
| Table 3: | Effect of Pythium sp. on rhodiola height, shoot vigour and shoot dry mass when grown under greenhouse conditions |
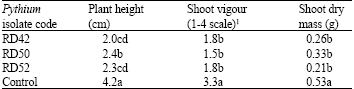 | |
| Data are the means of 10 replications. Means followed by the same letter(s) are not significantly different according to Fisher’s protected least significant difference test (p≤0.05). 1Vigour was assessed using 1-4 scale, where, 1 = poor; 2 = fair; 3 = fairly good and 4 = good growth of shoots and roots | |
| Table 4: | Effects of the interaction of Fusarium avenaceum (isolate RD18) and vesicular-arbuscular mycorrhizal fungi on growth of rhodiola under greenhouse conditions |
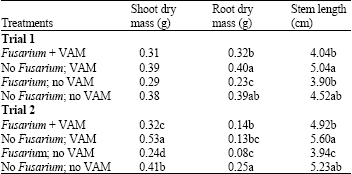 | |
| Data are the means of 10 replications. Means followed by the same letter(s) are not significantly different according to Fisher’s protected least significant difference test (p≤0.05) | |
The lowest shoot dry mass was recorded when the pathogen was inoculated alone in trial 2. The trend was similar in trial 1, although the treatment effects were not statistically significant. In both trials, the root dry mass was highest when VAM was added alone followed by treatments with no VAM or pathogen. This was followed by the treatments with both pathogen and VAM and was the lowest when the pathogen was added alone. A similar trend was observed for stem length.
| Table 5: | Effects of the interaction of Pythium sp. (isolate RD50) and vesicular-arbuscular mycorrhizal fungi on the growth of rhodiola under greenhouse conditions |
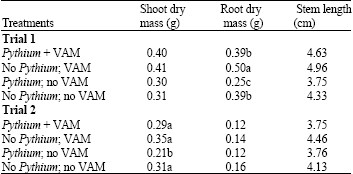 | |
| Data are the means of 10 replications. Means followed by the same letter(s) are not significantly different according to Fisher’s protected least significant difference test (p≤0.05) | |
| Table 6: | Effects of the interaction of Rhizoctonia solani (isolate RD12) and vesicular-arbuscular mycorrhizal fungi on the growth of Rhodiola under greenhouse conditions. |
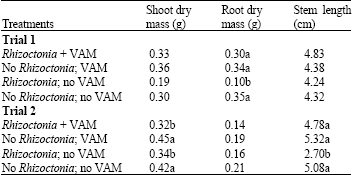 | |
| Data are the means of 10 replications. Means followed by the same letter(s) are not significantly different according to Fisher’s protected least significant difference test (p≤0.05) | |
In the case of seedlings inoculated with R. solani in trial 1, the shoot dry mass, the root dry mass and stem length were the lowest when VAM was not added with the pathogen, although shoot dry mass and stem length were not significantly different (Table 5). In trial 2, the shoot dry mass was the same in inoculated treatments whether VAM was applied or not. Similarly, for non-inoculated treatments, the shoot dry mass was similar in both VAM and non-VAM treatments. Similar trend was evident for root dry mass. However, stem length was significantly shorter when VAM was not added to Rhizoctonia-inoculated soil.
Shoot dry mass was significantly higher when VAM was added to Pythium-inoculated soil compared to pathogen inoculation alone in trial 2 (Table 6). However, the difference in shoot dry mass in trial 1 was not statistically significant. In trial 1, the root dry mass was highest in the treatment with VAM alone. Soil inoculated with Pythium sp. alone yielded the lowest root dry mass. In trial 2 the plant survival was reduced by Pythium sp. when VAM was not added. No significant differences in stem length were observed.
DISCUSSION
Because of the many beneficial pharmacological effects of rhodiola the commercial cultivation of this crop is increasing in Alberta. In this study, we identified Fusarium and Pythium sp. and R. solani as the principal root rot causing pathogens. These soil-borne pathogens could be of particular significance, since it takes 4-5 years after seeding to produce a rhodiola crop and the roots are the economically important organ. All three are common soil-borne pathogens and have wide host ranges (Adam, 1988; Chang et al., 2006; Leslie and Summerell, 2006). They may also remain viable in the soil for extended periods of time and can reduce seedling establishment in other crops (Hwang, 1992; Hwang et al., 2001).
In this study, we identified three Fusarium species, F. avenaceum, F. culmorum and F. equiseti in diseased rhodiola roots. These pathogens may cross-infect other crops like canola (Brassica napus L.), pea (Pisum sativum L.) and wheat (Triticum aestivum L.) and may cause severe stand and yield loss. A study by Zhou et al. (2009) indicated that the predominant anastomosis group of Rhizoctonia solani affecting lupin (Lupinus angustifolius L.) crops was AG-4, followed by AG-2-1 and AG-2-2. In this study, we showed that all of the Rhizoctonia isolates affecting rhodiola belonged to AG-4. Pythium sp. becomes most active in cool, moist soils. They are important early-season pathogens of canola, pulse crops and forage legumes.
Fusarium and Pythium sp. and R. solani all significantly reduced plant height, shoot vigour and shoot dry mass in rhodiola. Therefore, it is essential to develop a suitable disease management strategy to minimize the impact of these pathogens on rhodiola productivity. Among disease management strategies, seed treatments with fungicides may be one option to control damping off or seedling blight, thereby allowing for better stand establishment (Hwang et al., 2001). Considering the long cultivation period for rhodiola and the need to transplant the seedlings from greenhouse to field, it may be difficult to apply any chemical control measures other than chemical drenching, which has a negative impact on the environment and on the organic nature of the end product.
Vesicular-Arbuscular Mycorrhizal (VAM) fungi, used as biological control agents, have proven to be effective in controlling fungal pathogens (Pal and Gardener, 2006). In this study, while we demonstrated that Fusarium and Pythium sp. and R. solani all are capable of reducing rhodiola biomass, application of VAM not only ameliorated the effects of these pathogens, but improved biomass production in non-infested soils. Shoot dry mass was significantly lower in our study in Pythium-inoculated soil compared with non-inoculated soil. VAM fungi increase efficiency of phosphorus uptake (Chaudhary et al., 2008) which may contribute to the increase in shoot biomass found in this study. Several mechanisms are described how VAM interacts with the root systems to prevent pathogen infections. Pal and Gardener (2006) mentioned that during colonization VAM reduce the infection sites, stimulate host defenses and form an intricate network of fungal hyphae around the roots. This may block pathogen access to infection sites and may have resulted in the increase in biomass found in this study. However, this study suggests that VAM could be useful and appropriate as an organic disease management tool in the cultivation of rhodiola in Alberta. In addition, the VAM fungi are critically important for all crop plants, except for species of the Brassica family. A number of researchers have shown that an absence of VAM inoculum, or the insufficient concentrations of inoculum, can result in poor plant growth, in weak competition with other plants and/or in the inability to reproduce or survive under extreme conditions (Abbott and Robson, 1982; Haas and Krikun, 1985; Fernando and Liderman, 1997). Future studies should be directed towards optimizing the application of VAM fungi, in conjunction with the manipulation of environmental factors such as soil type, soil moisture, soil chemistry and temperature to improve the productivity of rhodiola cultivation in Alberta.
A total of 74 isolates were found to be associated with the root rot symptoms of rhodiola and in this study we included only the Fusarium, Pythium and Rhizoctonia isolates since these pathogens in most cases are the causal organisms of damping off, seedling blight and root rot in Alberta. However, the role of rest of the root rot associated mycoflora warrants investigation to explore their presence with root rot as they may be either beneficial as biocontrol agents or pathogenic to rhodiola. Furthermore, this study was conducted under controlled environmental conditions; field experiments should be conducted to validate the benefit of the use of VAM in the rhodiola cultivation system. We concluded that Fusarium, Pythium and Rhizoctonia are associated with root rot and reduced growth parameters of rhodiola and VAM was found to be effective to combat the negative impact of these pathogens on rhodiola.
ACKNOWLEDGEMENT
This study was supported in part by AVAC Ltd., Alberta and Alberta Natural Health Agriculture Network (ANHAN), Canada. The authors are thankful to Jackie Tieulie, Dereck Rennie and Michael Holtz for their help in conducting this experiment.
REFERENCES
- Abbott, L.K. and A.D. Robson, 1982. The role of vesicular arbuscular mycorrizal fungi in agriculture and the selection of fungi for inoculum. Aust. J. Agric. Res., 33: 389-408.
CrossRef - Brown, R.P., P.L. Gerbarg and Z. Ramazanov, 2002. Rhodiola rosea: A phytomedicinal overview. HerbalGram, 56: 40-52.
Direct Link - Fernando, W.G.D. and R.G. Linderman, 1997. The effect of mycorrhizal (Glomus intraradices) colonization on the development of root and stem rot (Phytophthora vignae) of cowpea. J. Natn. Sci. Coun. Sri Lanka, 25: 39-47.
Direct Link - Galambosi, B., 2005. Demand and availability of Rhodiola rosea L. raw material. Proceedings of the Frontis Workshop on Medicinal and Aromatic Plants, April 17-20, 2005, Waginengen, The Netherlands, pp: 223-236.
Direct Link - Haas, J.H. and J. Krikun, 1985. Efficacy of endomycorrizal-fungus isolates and inoculum quantities required for growth response. New Phytol., 100: 613-621.
CrossRef - Hwang, S.F., 1988. Effect of VA mycorhizae and metalaxyl on growth of alfalfa seedlings in soil from fields with alfalfa sickness in Alberta. Plant Dis., 72: 448-452.
CrossRef - Hwang, S.F., K.F. Chang and P. Chakravarty, 1992. Effects of vesicular-arbuscular mycorrhizal fungi on the development of Verticillium and Fusarium wilts of alfalfa. Plant Dis., 76: 239-243.
CrossRef - Yao, M., R. Tweddell and H. Desilets, 2002. Effect of two vesicular-arbuscular mycorrhizal fungi on the growth of micropropagated potato plantlets and on the extent of disease caused by Rhizoctonia solani. Mycorriza, 12: 235-242.
CrossRefDirect Link - Chaudhary, V., R. Kapoorand and A.K. Bhatnagar, 2008. Mycorrhizal fungi on concentrations of essential oil and artemisinin in three accessions of Artemisiinin annua L. Applied Soil Ecol., 40: 174-181.
CrossRef - Chang, K.F., S.F. Hwang, B.D. Gossen, R.L. Conner and D.L. McLaren et al., 2006. Host range and cultivar response to Fusarium root rot of field pea. Proceedings of the 6th Canadian Pulse Research Workshop, Nov. 1-3, Saskatoon, SK., pp: 1-2.
Direct Link