
Idorenyin A. Udo
Department of Crop Science, University of Calabar, Calabar, Nigeria
Michael I. Uguru
Department of Crop Science, University of Nigeria, Nsukka, Nigeria
Rufus O. Ogbuji
Department of Crop Science, University of Nigeria, Nsukka, Nigeria
Donald A. Ukeh
School of Biological Sciences, University of Aberdeen, Tillydrone Avenue, AB24 2TZ, United Kingdom
Plant Pathology Journal
Year: 2008 | Volume: 7 | Issue: 1 | Page No.: 40-44







ABSTRACT
Five elite tomato cultivars (HT-5044, HTA-18, HTA 19, HTA-31 and HT-Ravid 244), two locally gown Nigerian cultivars (Roma VF and Nsukka local) and a wild tomato related species (Lycopersicon pimpinellifolium) were assayed for their responses to an indigenous population of M. javanica in the greenhouse. Infected plants were scored for galling on a 0-5 rating scale. The cultivars HTA-18 and HTA-31 had significantly (p< 0.05) fewer gall counts per root system and per gram fresh weight of root than the other cultivars. Single degree of freedom analysis showed that HTA-18, HTA-19 and HTA-31 had significantly (p< 0.05) fewer galls than the locally grown cultivars. HTA-18 and HTA-31 were moderately susceptible with Gall Index (GI) of 3, while the rest of the cultivars were highly susceptible (GI>4). The nematode caused a significant delay in flowering and a decrease in fresh and dry weights of roots, stems and leaves in the cultivars, HT-5044, Roma VF and Nsukka local. HTA-18, HT-Ravid 244 and the wild species were considered tolerant.
PDF Abstract XML References Citation
How to cite this article
Idorenyin A. Udo, Michael I. Uguru, Rufus O. Ogbuji and Donald A. Ukeh, 2008. Sources of Tolerance to Root-Knot Nematode, Meloidogyne javanica, in Cultivated and Wild Tomato Species. Plant Pathology Journal, 7: 40-44.
DOI: 10.3923/ppj.2008.40.44
URL: https://scialert.net/abstract/?doi=ppj.2008.40.44
DOI: 10.3923/ppj.2008.40.44
URL: https://scialert.net/abstract/?doi=ppj.2008.40.44
INTRODUCTION
The production of tomato under field conditions is highly limited by pests and diseases in the tropics (Uguru, 1996; Uguru and Igili, 2002). The crop is highly susceptible to the root-knot disease caused by Meloidogyne spp. (Montasser et al., 1986; Sorribas et al., 2005). The symptoms of attack include root gallings and some forms of root distortions that impair nutrient and water uptake with severe implications in growth and development (Melakeberhan et al., 1990; Williamson and Gleason, 2003). Although accurate information on yield losses attributable to root-knot nematode in Nigeria is unavailable, conservative reports indicated more than 50% losses depending on the cultivars, population density, cultural practices and environmental conditions (Olowe, 2004, 2005). Three Meloidogyne species namely; M. incognita, M. javanica and M. arenaria are found in Nigerian soils (IITA, 1992) and are associated with severe attacks on vegetable crops (Jonathan and Hedwig, 1991). Chemical control method of nematode species in Nigeria has been discouraged due to the prohibitive cost as well as the hazards associated with the use of chemicals. Generally, wild relatives of the cultivated plant species have contributed useful genes for resistance to pests and diseases (Stalker, 1980). Nematode-resistant accessions of wild relatives of potato, tobacco and tomato have been successfully used as sources of genes for resistance to cyst and root-knot nematodes (Cook and Evans, 1987). The wild tomato species, Lycopersicon pimpinellifolium has been an important reservoir of useful genes for resistance against many diseases of tomato (Willamson, 1998; Uguru and Igili, 2002). This study is aimed at identifying resistance to an indigenous population of M. javanica in cultivated tomatoes and a wild accession.
MATERIALS AND METHODS
Seven cultivated tomato and a closely related wild species, L. pimpinellifolium, were used in the study. Five of the commercial varieties: HT-5044, HTA-18, HTA-19, HTA-31 and HT Ravid 244, were procured from Israel, while Roma VF, Nsukka Local and the wild species were obtained from the Department of Crop Science, University of Nigeria, Nsukka tomato germplasm. Seeds of each cultivar were surface sterilized separately in 0.5% chlorox for 5 min and washed three times in tap water. Seedlings were raised in steam-sterilized soil mixture of sandy loam, compost soil and river sand mixed at a ratio 3:2:1 by volume. Nigerian population of M. javanica maintained on begonia plants (Begonia rex-cultorum) served as inoculum source. This population was multiplied on Indian Spinach (Bassela rubra) in the greenhouse. Root-knot nematode eggs were extracted from the heavily galled roots of the Indian spinach using sodium hypochlorite (NaOCl) technique (Hussey and Barker, 1973). Thirty millilitre of the inoculum suspension contained approximately 5,000 eggs. Four-week-old tomato seedlings were transplanted on 1st November, 2002, into 15 cm diameter clay pots containing 1 kg steam-sterilized soil mixture. Seedlings were inoculated with 5,000 eggs per pot by adding the inoculum in depression made in the soil round the young seedlings. Uninoculated seedlings of each cultivar served as the control. The experiment was a Completely Randomized Design (CRD) with ten replicates. The pots were placed on the greenhouse benches. The plants were watered as and when necessary. Twenty grams of a compound fertilizer N-P-K (15-15-15) was dissolved in 30 L of tap water and was applied every other day throughout the duration of the experiment (Oscar and Donald, 1988). Plants were grown at mean day temperature of 30°C and mean night temperature of 20°C for 79 days. Data were collected on number of galls per root system, number of galls per gram fresh weight of root as determined by (Khan and Khan, 1995), fresh weights of root, stem and leaves, number of fruits per plant, days to flowering and total weight of fruits per plant. Roots, stems and leaves were dried in hot air oven at 60°C for 48 h to determine dry weights. Root-gall index was determined on a 0-5 scale rating used in the International Meloidogyne Project (IMP, 1978): 0 = 0, 1 = 1-2, 2 = 3-10, 3 = 11-30, 4 = 31-100 and 5 = >100 galls per root system.
Statistical analysis: The statistical software package MINITAB 15 was utilized for data analysis using the analysis of variance (ANOVA). Means were tested using the F-LSD at 5% level of probability. The uninoculated (control) were compared with the inoculated treatments using the t-test. The elite cultivars were compared with the locally grown Nigerian cultivars by single degree of freedom analysis thus:
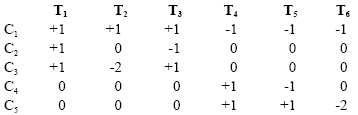 |
Where, T1, T2 and T3 represent treatment totals for the elite cultivars; T4, T5 represent treatment totals for the locally grown cultivars and T6 represents the wild species. C1 to C5 are orthogonal set of comparisons. The positive and negative integers are contrast coefficients.
RESULTS
The tomato cultivars differed significantly (p<0.05) in their galling responses (Table 1). The cultivars, HTA-18 and HTA-31 had significantly fewer gall counts per root system and per gram fresh weight of root than the others. HTA-18 and HTA-31 were moderately susceptible with Gall Index (GI) of 3.0, while the rest of the cultivars had gall indices greater than 4.0. Comparing HTA-18, HTA-19 and HTA-31 on one hand with the three locally grown lines on the other by single degree of freedom analysis showed that the former had significantly (p<0.01) fewer gall counts per root system and per gram fresh weight of root than the latter (Table 1). The fresh and dry weights of root, stem and leaves of the uninoculated controls were greater than those of the inoculated plants in all the test cultivars (Table 2). However, the decrease in dry matter production by M. javanica was statistically significant only in cultivars HT-5044, Roma VF and Nsukka Local. There was no significant decrease in fresh and dry weights of root, stem and leaves in the wild species (Table 2). The cultivars differed significantly (p<0.05) in fresh and dry weights of root, stem and leaves (Table 2). HTA-18, HTA-31, HT-Ravid 244 and the wild species had relatively higher fresh and dry root weights than the other cultivars. Similarly, HTA-18 and HTA-19 had relatively higher fresh and dry leaf weights than the other cultivars. The tomato cultivars differed significantly (p<0.05) in days to flowering. Inoculation with nematodes tended to prolong the flowering periods in each cultivar (Table 3). M. javanica significantly delayed flowering in the cultivars HT- 5044, HTA-19 and Roma VF when compared with the uninoculated controls. Except for HT-Ravid 244, the nematode inhibited fruit-set in all the test cultivars when compared with their respective controls. However, inhibition of fruit-set was not significant in HT-5044 and the wild species (Table 3). The wild species produced more fruits than the other cultivars. The nematode also caused a decline in total weight of fruits per plant in all the test cultivars when compared with their respective controls. Yields were suppressed by 35, 32, 47, 38, 21, 63, 65 and 22% in the cultivars, HT-5044, HTA-18, HTA-19, HTA-31, HT-Ravid 244, Roma VF, Nsukka Local and the wild species, respectively (Table 3).
| Table 1: | Susceptibility of cultivated and wild species of tomatoes to Meloidogyne javanica and single degree of freedom analysis showing mean squares for number of galls |
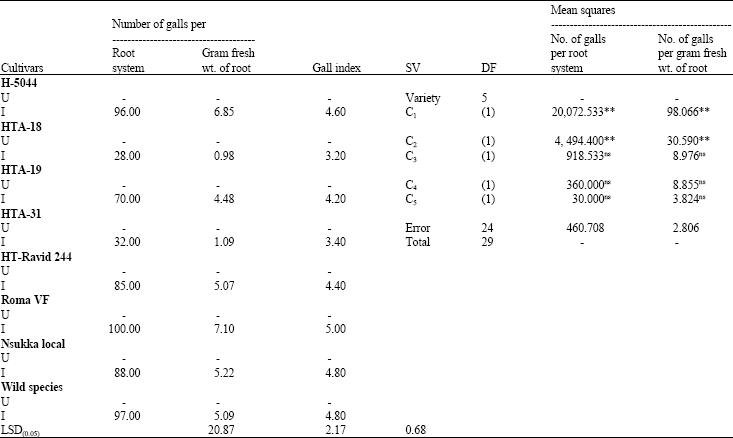 | |
| U = Uninoculated control, I = Inoculated, ** = Significant at 1% level of probability, ns = Not significant, C1 = Compares the elite with the locally grown cultivars, C2 = Compares HTA-18 and HTA-19, C3 = Compares (HTA-18, HTA-19) and (HTA-31), C4 = Compares Roma VF and Nsukka local, C5 = Compares (Roma VF, Nsukka Local) and the wild species | |
| Table 2: | Effect of Meloidogyne javanica on dry matter production of cultivated and wild species of tomato plants |
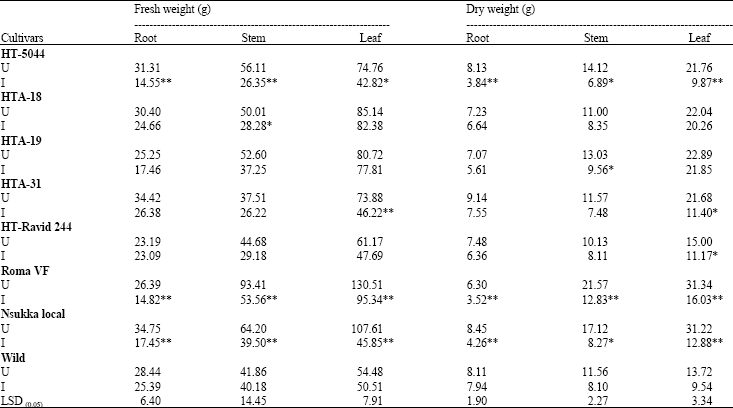 | |
| U = Uninoculated control, I = Inoculated, LSD compares the means of cultivars inoculated with the nematode species, *, **: Significantly different from their respective controls at 5 and 1%, respectively, using the t-statistic | |
| Table 3: | Effect of Meloidogyne javanica on days to first flowering and yield of cultivated and wild species of tomato plants |
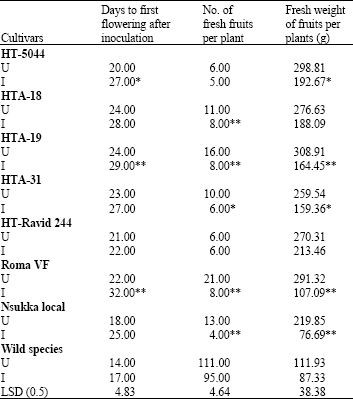 | |
| U = Uninoculated control, I = Inoculated, LSD Compares the means of cultivars inoculated with the nematode species, *, **: Significantly different from their respective controls at 5 and 1%, respectively, using the t-statistic | |
Statistically, yields were not significantly suppressed in HTA-18, HT-Ravid 244 and the wild species. There were significant yield differences among the cultivars. The yields of the elite cultivars were higher than their counterparts.
DISCUSSION
Evidence from this study showed that all the test cultivars were susceptible to M. javanica. However, HTA-18 and HTA-31 were moderately susceptible. This apparent susceptibility of wild species may be attributed to the different races of the nematode species used or to environmental effects on the plant cultivars and nematode species (Tzortzakakis and Gowen, 1996; Castagnone-Sereno, 2002). Although, the wild species was susceptible to the nematode, it appeared to posses some innate attributes for tolerance as indicated by the high dry matter production and fresh fruit yield. Dry matter production in the roots, stems and leaves were not significantly reduced in the wild species. A plant that survives and gives satisfactory yield at a level of infection that causes economic loss on other varieties of the same species is tolerant. Canto-Saenz (1985) defines a tolerant plant as an efficient host that suffers no statistically significant growth and yield reduction. Host efficiency is measured by gall index or index of nematode reproduction. The significant decrease in dry matter contents of the other susceptible cultivars and the discernable decline in fresh fruit yield are indices of intolerance to the nematode species. The metabolic activities of an economic plant are reflected in its total dry matter content and yield. The delay in flowering and reduction in plant biomass of the susceptible and non-tolerant cultivars appear to suggest a decrease in metabolic activities. Fox and Spasoff (1976) had shown that tolerance to attack by Globodera solanacearum in tobacco was genetically independent of resistance and could be identified and selected. It follows then that, if tomato lines tolerant of root-knot nematode attack were identifiable and the trait inherited, tolerance could be incorporated in cultivars for commercial use. Ogbuji (1981) and Evans and Franco (1979) reported tolerance in Lima bean cv. 6405 and potato cv. Pentland crown to infection by M. incognita race I. and G. rostochinensis RoI, respectively. Recently, the wild genotype of the species L. pimpinellifolium INRA-Hirsute has been reported to confer tolerance and resistance to the vector transmission of tomato yellow leaf curl virus (Delatte et al., 2006). Tolerance is sometimes achieved through the compensating growth of new roots when existing ones had been damaged by the plant parasitic nematodes. This would make up for the impaired nutritional intake which the plant would have suffered as result of infection. Vidavsky et al. (1998) reported that genes involved in tolerance originating from L. pimpinellifolium provides different levels of protection in tomatoes and combinations of various tolerant sources and levels in a single genotype could give a higher level of tolerance.
Therefore, L. pimpinellifolium could be a potential gene source for tolerance to M. javanica. Similarly, the non-significant reduction in dry matter contents and yield in the cultivars, HTA-18 and HT-Ravid 244-would appear to suggest some degree of tolerance of the two varieties to M. javanica. However, planting materials developed from these cultivars should be judiciously used to curb the menace of M. javanica in heavily infested soil as tolerant plants may increase pre-plant nematode density tremendously. In such cases, other cultural practices, such as soil desiccation may be adopted. Further screening of other accessions of the wild species may offer solutions to the endemic problem of root-knot nematodes of tomatoes in the humid tropics.
REFERENCES
- Castagnone-Sereno, P., 2002. Genetic variability in pathenogenic root-knot nematodes, Meloidogyne spp. and their ability to overcome plant resistance genes. Nematology, 4: 605-608.
CrossRefDirect Link - Delatte, H., H. Holota, B. Reynaud and J. Dintinger, 2006. Characterisation of a quantitative resistance to vector transmission of Tomato yellow leaf curl virus in Lycopersicon pimpinellifolium. Eur. J. Plant Pathol., 114: 245-253.
Direct Link - Evans, K. and J. Franco, 1979. Tolerance to cyst-nematode attack in commercial potato cultivars and some possible mechanisms for its operation. Nematology, 25: 153-162.
CrossRefDirect Link - Hussey, R.S. and K.R. Barker, 1973. Comparison of methods of collecting inocula of Meloidogyne spp., including a new technique. Plant Dis. Rep., 57: 1025-1028.
Direct Link - IMP (International Meloidogyne Project), 1978. Section V. Guidelines for conducting differential host test for Meloidogyne species. Proceedings of the Research Planning Conference on Root-Knot Nematode Meloidogyne sp., June 7-11, 1976, Nematology Research Centre Cairo University, Giza Egypt, pp: 85-85.
- Khan, M.R. and M.W. Khan, 1995. Effects of ammonia and root-knot nematode on tomato. Agric. Ecosyst. Environ., 53: 71-81.
CrossRefDirect Link - Melakeberhan, H., H. Ferris and J. Dias, 1990. Physiological response of resistant and susceptible Vitis vinifiera cultivars to Meloidogyne incognita. J. Nematol., 22: 224-230.
Direct Link - Olowe, T., 2004. Occurrence and distribution of root-knot nematodes, Meloidogyne spp., in cowpea growing areas of Nigeria. Nematology, 6: 811-817.
CrossRefDirect Link - Sorribas, F.J., C. Ornat, S. Verdejo-lucas, M. Galeano and J. Valero, 2005. Effectiveness and profitability of the Mi-resistance tomatoes to control root-knot nematodes. Eur. J. Plant Pathol., 111: 29-38.
Direct Link - Stalker, H.T., 1980. Utilization of wild species for crop improvement. Adv. Agron., 33: 111-147.
CrossRefDirect Link - Tzortzakakis, E.A. and S.R. Gowen, 1996. Occurrence of a resistance breaking pathotype of Meloidogyne javanica on tomatoes in Crete, Greece. Applied Nematol., 19: 283-288.
Direct Link - Uguru, M.I. and D.N. Igili, 2002. Field reactions of segregating populations of interspecific hybrids of Lycopersicon species to natural infection by Xanthomonas campestris P.V vesicatoria (Diodge) dye. Nig. J. Hortic. Sci., 6: 5-11.
Direct Link - Vidavsky, F., S. Leviatov, J. Milo, H.D. Rabinowitch, N. Kedar and H. Czosnek, 1998. Response of tolerant breeding lines of tomato, Lycopersicon esculentum, originating from three different sources (L. peruvianum, L. pimpinellifolium and L. chilense) to early controlled inoculation by Tomato Yellow Leaf Curl Virus (TYLCV). Plant Breed., 117: 165-169.
CrossRefDirect Link - Williamson, V.M., 1998. Root-Knot nematode resistance genes in tomato and their potential for future use. Annu. Rev. Phytopathol., 36: 277-293.
CrossRefPubMedDirect Link - Williamson, V.M. and C. Gleason, 2003. Plant-nematode interactions. Curr. Opin. Plant Biol., 6: 327-333.
CrossRefDirect Link