
G.M. Riungu
Department of Plant Science and Crop Protection, University of Nairobi, P.O. Box 30197, 00100 GPO, Nairobi, Kenya
J.W. Muthomi
Department of Plant Science and Crop Protection, University of Nairobi, P.O. Box 30197, 00100 GPO, Nairobi, Kenya
R.D. Narla
Department of Plant Science and Crop Protection, University of Nairobi, P.O. Box 30197, 00100 GPO, Nairobi, Kenya
J.M. Wagacha
Department of Plant Science and Crop Protection, University of Nairobi, P.O. Box 30197, 00100 GPO, Nairobi, Kenya
J.K. Gathumbi
Department of Veterinary Pathology, Microbiology and Parasitology, University of Nairobi, P.O. Box 30197, 00100 GPO, Nairobi, Kenya
Plant Pathology Journal
Year: 2008 | Volume: 7 | Issue: 1 | Page No.: 13-19







ABSTRACT
Laboratory and green house studies were conducted at the Faculty of Agriculture, University of Nairobi, to evaluate the efficacy of Epicoccum sp., Alternaria sp., Trichoderma sp. and Bacillus sp. in control of Fusarium head blight of wheat caused by F. graminearum. Fungicides folicur® and copper oxychloride were used as standard checks. Laboratory assay was carried out by paired cultures and antagonism was measured as reduction in pathogen colony diameter. Green house experiments involved dual inoculation of pathogen and antagonist onto wheat ears and head blight severity and grain yield determined. Doxynivalenol content in the resulting grain was determined by competitive direct ELISA. The antagonists and fungicides significantly reduced the growth of Fusarium graminearum colonies in culture. Folicur® and copper oxychloride completely inhibited the growth of the pathogen while Trichoderma sp. showed 64% colony growth reduction. However, the antagonists showed limited reduction in head blight severity in green house trials. Trichoderma sp. reduced head blight severity by 18% while folicur® reduced the disease by 28%. All the antagonists had little or no significant effect on grain yield. Only folicur®, copper oxychloride and Alternaria sp. reduced DON in grain by 76 to 93%. Obtained results indicate that microbial antagonists may offer potential benefit in FHB management and screening of more antagonists both under controlled and field conditions is necessary.
PDF Abstract XML References Citation
How to cite this article
G.M. Riungu, J.W. Muthomi, R.D. Narla, J.M. Wagacha and J.K. Gathumbi, 2008. Management of Fusarium Head Blight of Wheat and Deoxynivalenol Accumulation Using Antagonistic Microorganisms. Plant Pathology Journal, 7: 13-19.
DOI: 10.3923/ppj.2008.13.19
URL: https://scialert.net/abstract/?doi=ppj.2008.13.19
DOI: 10.3923/ppj.2008.13.19
URL: https://scialert.net/abstract/?doi=ppj.2008.13.19
INTRODUCTION
Fusarium Head Blight (FHB) is a serious disease of small grain cereals and has caused severe and repeated epidemics resulting in enormous losses (Bateman, 2005; Kolombet et al., 2005; Brennan et al., 2007). In addition to grain yield reduction, FHB can result in the reduction of grain quality, either by affecting grain processing qualities or by producing a range of toxic metabolites that have adverse effects on humans and livestock (Dohlman, 2004; Grosjean and Barrier-Guillot, 2004; Goyarts et al., 2007). Fusarium graminearum, one of the major causal organisms of FHB, produces mycotoxin Deoxynivalenol (DON), which may accumulate to unacceptable levels in harvested grain (Demeke et al., 2005; Paul et al., 2005; Browne, 2007). Levels of DON above 2 ppm may render grain and their by-products unfit for commercialization and consumption.
Efforts to minimize the impact of FHB and DON have been centred on the use of management strategies such as crop rotation, host plant resistance, tillage and fungicides application (Heier et al., 2005; Wisniewska and Kowalczyk, 2005; Kriel, 2006; Browne, 2007). An integrated approach to management of FHB that includes chemical, cultural and host plant resistance seems the most logical way to reduce losses (Pirgozliev et al., 2003; Pereyra and Dill-Macky, 2004). Several studies on chemical control of FHB have been reported but conflicting evidence exists regarding the effect of fungicides on the development of FHB and accumulation of trichothecene mycotoxins in grain (Halley et al., 2005; Ramirez et al., 2004). Fungicides have been found to affect DON concentrations indirectly by influencing the inoculum of Fusarium species in the grain (Suty et al., 1996; Pirgozliev et al., 2003). Fungicide application has been reported to reduce FHB severity without significant reduction of DON (Edwards et al., 2001). However, Draper et al. (2005) found no effect on both DON and the disease severity after using folicur® and metconazole. Ramirez et al. (2004) tested four azoles and one strobulurin and found that while all of them reduced F. graminearum growth in vitro, there was reduced efficacy on the same pathogen when applied in the field. The fungicides had a varied effect on production and accumulation of DON with some resulting in stimulation of DON production.
All labelled systemic fungicides appear to increase yield, but those that contain a triazole, instead of a strobilurin active ingredient are more effective in reducing mycotoxin (DON) levels in infected grain (Hollingsworth, 2004; Ramirez et al., 2004; Müllenborn et al., 2007). However, use of fungicides on wheat ears has the disadvantage of accumulation of residues in the resulting grain. The use of biological control would lead to reduction, if not elimination of the possible chemical residues in grain, environmental pollution and potential hazards to humans. Therefore, this study was carried out with the objective of evaluating the efficacy of fungal and bacterial antagonists in management of FHB caused by F. graminearum.
MATERIALS AND METHODS
Laboratory and green house studies were conducted in 2006 at the Faculty of Agriculture, University of Nairobi, to evaluate the efficacy of Epicoccum sp., Alternaria sp., Trichoderma sp. and Bacillus sp. to control Fusarium head blight of wheat caused by F. graminearum.
Isolation and multiplication of pathogen and antagonists: Isolates of Fusarium graminearum, Epicoccum, Alternaria and Trichoderma sp. were isolated from wheat kernels by plating on low strength PDA amended with mineral salts and antibiotics (Muthomi, 2001; PDA 17 g, KH2PO4 1.0 g, KNO3 1.0 g, MgSO4 0.5 g and Agar 10 g). The fungi were identified based on cultural and morphological characteristics like colony colour, pigment production, presence of aerial mycelium in addition to morphological characteristics like conidia shape, septation and conidiophores. Bacillus sp. was also isolated from wheat seeds plated on Nutrient Agar (NA) and identified based on cultural characteristics.
Inoculum of three highly pathogenic isolates of F. graminearum was produced in mung bean broth (Bai and Shaner, 1994). Mung bean (40 g) was cooked in 1000 mL of water for 10 min and the extract was filtered through double layer cheesecloth, the volume completed and the extract sterilized. The sterilized mung bean extract was inoculated with mycelial agar discs cut from 14 day old pathogen cultures and placed on a mechanical shaker (50-70 cycles min-1) for 4 days followed by 10 day incubation under stationary conditions. Epicoccum, Alternaria and Trichoderma sp. were raised on PDA for 14 days at 25°C in cycles of 12 h daylight and 12 h darkness while Bacillus sp. was grown on nutrient agar for 2 days. Pathogen inoculum was harvested by passing the liquid culture through double layer cheesecloth while that of the antagonists was by flooding the cultures with distilled water and passing the solution through a double layer of cheese cloth. The fungal inoculum was adjusted to 1x105 spores mL-1 using a haemocytometer while Bacillus sp. inoculum was adjusted to about 1 x104 cfu mL-1.
Determination of efficacy of antagonists to suppress growth of F. graminearum in culture: Antagonism was determined by paired cultures method, where the pathogen agar disc was inoculated at the middle of plate and the antagonist at 4 equidistant points located 2 cm from plate edge. Each of the antagonists Epicoccum, Alternaria, Trichoderma and Bacillus sp. was tested separately. Fungicides folicur® (1000 ppm) and copper oxychloride (1000 ppm) were used as standard checks while negative control consisted of F. graminearum cultured alone. Each treatment was replicated four times and the plates arranged in a completely randomized design on laboratory benches and incubated at 25°C for 7 days in cycles of 12 h daylight and 12 h darkness. Degree of antagonism was determined by measuring the pathogen colony diameters and percentage inhibition calculated:
Efficacy of antagonists to reduce FHB under greenhouse conditions: Highly susceptible wheat variety Mbuni, was planted in 22 cm diameter pots in greenhouse following recommended fertilization regimes. Inoculation was done at 50% flowering (GS 65; Zadoks et al., 1974) and treatments consisted of inoculation of ears with antagonist together with F. graminearum, F. graminearum together with fungicide, antagonist alone and F. graminearum alone. The antagonists tested were Epicoccum, Alternaria, Trichoderma and Bacillus sp. Folicur®, a systemic fungicide and copper oxychloride, a contact fungicide, were used as standard checks at the rate of 1.5 and 6 g L-1 of water, respectively. Control plants were sprayed with sterile distilled water. The antagonists and the fungicides were sprayed two days before and after inoculation with F. graminearum. Inoculation with F. graminearum was repeated 5 days after the first inoculation. Each treatment was replicated four times and arranged in a randomised complete block design. The treated heads were covered with polythene bags for 48 h to maintain humidity conducive for infection.
Head blight severity was assessed five days after the last inoculation and after every 5 days thereafter until ripening (GS 87). Proportion of ear bleached was determined based on a 1-9 scale, where 1% = no symptoms, 2 = <5%, 3 = 5-15%, 4 = 16-25%, 5 = 26-45%, 6 = 46-65%, 7 = 66-85%, 8 = 86-95%, 9 = 96-100% of spikelet bleached. Assessment was done on ten average-sized ears per pot. Mean disease severity and the area under disease progress curve (AUDPC; Shaner and Finney, 1977) were calculated from single ratings recorded over the assessment period. At maturity (GS 95) the ten heads assessed for disease were harvested and threshed separately for weight determination. The kernels harvested (100 kernels per treatment) were plated on culture media for re- isolation of the causal pathogen. The experiment was conducted over two greenhouse cycles.
Analysis of Deoxynivalenol (DON) content: Deoxynivalenol in the grain from greenhouse experiments was analyzed by direct competitive Enzyme-Linked Immunosorbent Assay (ELISA; Gathumbi et al., 2001). Each sample was homogenized and 100 g ground to fine powder. Five grams of the ground sample was extracted with 25 mL of methanol:water (50:50 v/v). The extract was de-fatted with 10 mL hexane and 4 mL of the methanolic layer taken and diluted to 10% using Phosphate Buffer Solution (PBS). Microtitre polystyrene plates were coated with 100 μL of anti-deoxynivalenol antiserum DON143/16 (Usleber et al., 1992) in bicarbonate buffer (pH 9.6) per well. Absorbance was determined using spectrophotometer Elisa reader at 450 nm wavelength. A calibration curve for the standard dilutions was plotted using log10 of standard concentration against the percentage inhibition of the standard.
Data analysis: All data was subjected to Analysis of Variance (ANOVA) using the PROC ANOVA procedure of Genstat (Lawes Agricultural Trust Rothamsted Experimental station 2006, Version 9) and differences among the treatment means were compared using the Fisher’s protected LSD test at 5% probability level.
RESULTS
All the antagonists and fungicides tested significantly (p≤0.05) reduced colony diameters of F. graminearum (Table 1). Folicur® and copper oxychloride completely inhibited the growth of F. graminearum in vitro. The highest colony diameter reduction (64%) was observed in the treatment with Trichoderma sp. while the least reduction (45%). was observed in paired cultures with Epicoccum sp.
| Table 1: | Colony diameter (cm) and percentage reduction in colony diameter of F. graminearum in paired cultures |
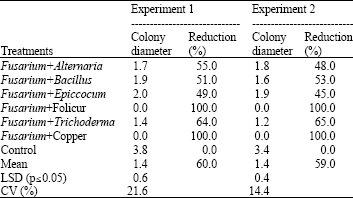 | |
| LSD: Least Significant Difference; CV: Coefficient of Variation | |
| Table 2: | Severity percentage of FHB over time, mean severity, AUDPC and re-isolation rate for the plants inoculated with F. graminearum and the respective antagonists |
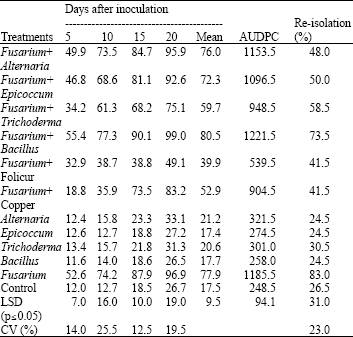 | |
| AUDPC: Area Under Disease Progress Curve; LSD: Least Significant Difference; CV: Coefficient of Variation | |
Head blight severity and the Area Under Disease Progress Curve (AUDPC) were significantly different (p≤0.05) among the antagonists (Table 2). Folicur® reduced the disease severity by up to 47% while copper oxychloride reduced disease severity by up to 36%. Among the antagonists, Trichoderma sp. was the most effective with a significant reduction of FHB of up to 25%. Epicoccum and Alternaria sp. had minimal effect on FHB and Bacillus sp. had no significant effect on head blight severity. Minimal amounts of disease were observed on plants inoculated with antagonist alone and the control. Fusarium graminearum was re-isolated at very high levels from kernels harvested from ears inoculated with the pathogen alone but the re-isolation rate differed for the kernels from ears inoculated with different antagonists.
| Table 3: | Ten-ear weight (g) and weight per pot (g) for plants treated with F. graminearum and respective antagonists |
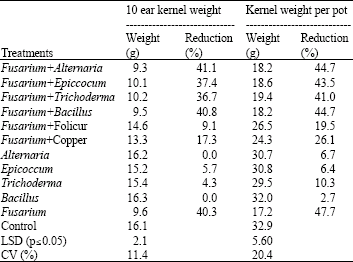 | |
| LSD: Least Significant Difference; CV: Coefficient of Variation | |
| Table 4: | Deoxynivalenol (DON) content (μg kg-1) of wheat grain harvested from ears inoculated with different antagonistic microorganisms |
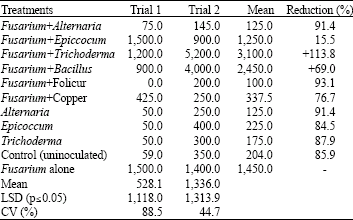 | |
| LSD: Least Significant Difference; CV: Coefficient of Variation | |
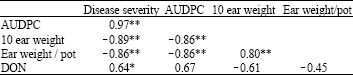
| Table 5: | Correlation coefficients (p<0.05) among disease severity, Area Under Disease Progress Curve (AUDPC), grain weight and Deoxynivalenol (DON) content |
 | |
| *: Indicates significant correlation; **: Indicates highly significant correlation at p≤0.05 | |
The antagonists had little or no significant effect on grain weight (Table 3). However, folicur® and copper oxychloride significantly (p≤0.05) increased grain weight by between 47 and 94%, respectively, compared to ears inoculated with F. graminearum alone. Folicur®, copper oxychloride and Alternaria sp. reduced DON content in the grain by between 76 and 93% but Trichoderma and Bacillus sp. increased DON compared to treatments with F. graminearum alone (Table 4). Head blight severity, kernel weight and DON content in grain were significantly (p≤0.05) correlated (Table 5).
DISCUSSION
All the antagonists inhibited the growth of F. graminearum in culture, indicating a possible release of extracellular volatile metabolites that diffused through the media. However, folicur® and copper oxychloride were the most effective, inhibiting the growth of the pathogen completely. In the two greenhouse trials, the fungicides reduced the disease severity by between 28-58%, although no complete control was found, therefore confirming earlier findings (Chala et al., 2003; Müllenborn et al., 2007). The low disease levels observed on plants inoculated with antagonist alone and the control could have been caused by pathogen conidia spread by wind from the inoculated ears. Among the antagonists, Trichoderma sp. reduced disease severity and this is in agreement with the finding by Perello et al. (2002), Lutz et al. (2003) and Müllenborn et al. (2007). Perello et al. (2002) reported that among the antagonists tested, Bacillus sp. was the one with the highest inhibitory effect to pathogens in culture. Epicoccum purpurascens (E. nigrum) produces antifungal compounds, which may increase its effectiveness (Brown et al., 1987). This has also been reported to be true for Trichoderma and Bacillus sp. (Schunmacher et al., 2007; Seddon, 2007). Production of antifungal secondary metabolites by Trichiderma atroviride can induce resistance of plants against infection by pathogenic microorganisms.
Application of folicur® and copper oxychloride led to an increase in yield thus confirming earlier findings by Pirgozliev et al. (2003) and Mesterhazy et al. (2003). The effect of fungicides on Fusarium sp. is dependent on timing and frequency of applications (Parry et al., 1995) and treatments at anthesis seems to be the best time for reduction of Fusarium infection (Mesterhazy et al., 2003; Hollingsworth, 2004). In the current study, Trichoderma sp. was found to reduce both disease severity and slightly increase the grain yield. This shows that biological control has considerable promise for reducing FHB as has been reported by Kolombet et al. (2005) and Seddon (2007). Bateman (1979) demonstrated biological control of Microdochium nivale with Sporobolomyces sp. which significantly reduced grain contamination. Rodemann (2007) reported a 40 and 30% reduction of FHB severity and DON, respectively on wheat treated with the antagonist Plectosporium tabacinum at anthesis under greenhouse conditions. Inoculations before anthesis with Cladosporium sp. were effective when applied before M. nivale, while Alternaria sp. were effective whether applied before or after. Pseudomonas fluorescens biov1, B. subtilis and Streptomyces sp. were found to be antagonistic to F. graminearum. Fusarium graminearum mycelial growth was reduced by cell free and volatile metabolites of the bacterial antagonists by 37-97%. However, Draper et al. (2005) and Jochum et al. (2004) found no yield benefits when using Lysobacter enzymogenes, Bacillus sp. and Pseudomonas fluorescens alone but dual application with folicur® led to an increase in test kernel weight. A recent trial by Bacon and Hinton (2007) using Bacillus mojavensis showed that several strains of the bacterium were antagonistic to F. graminearum and F. verticillioides in vitro and in vivo. Fusarium is capable of suppressing the expression of the genes that control the antagonistic activity of Trichoderma harzianum but not that of Pseudomonas fluorescens (Brion, 2001; Lutz et al., 2003). A recent study by Seddon (2007) reported susceptibility of Fusarium sp. to Bacillus brevis.
The antagonist effect could be explained by two mechanisms. Firstly, through production of antifungal metabolites which inhibits conidial germination and subsequent growth of the pathogen (Brunner et al., 2003; Seddon, 2007), Secondly, by indirect action through production of a biosurfactant, which modifies the plant surface reducing surface wetness. From the results obtained in this study, it is clear that all the antagonists reduced the growth of F. graminearum in culture indicating that they produced antifungal compounds. The results also indicate that three antagonists reduced FHB severity although not very pronounced. Testing of a broader spectrum of other possible antagonists under field conditions and determination of their mechanisms of action would be necessary.
ACKNOWLEDGMENT
This research was funded by the International Foundation for Science (IFS) grant No. E/3654-1.
REFERENCES
- Bacon, C.W. and D.M. Hinton, 2007. Potential for control of seedling blight of wheat caused by Fusarium graminearum and related species using bacterial endophyte Bacillus mojavensis. Biocontrol Sci. Technol., 17: 81-94.
Direct Link - Bateman, G.L., 2005. The contribution of ground-level inoculum of Fusarium culmorum to ear blight of winter wheat. Plant Pathol., 54: 299-307.
Direct Link - Brennan, J.M., G. Leonard, B. Fagan, B.M. Cooke and A. Ritieni et al., 2007. Comparison of commercial European wheat cultivars to Fusarium infection of head and seedling tissue. Plant Pathol., 56: 55-64.
Direct Link - Browne, R.A., 2007. Components of resistance to Fusarium head blight (FHB) in wheat detected in a seed-germination assay with Microdochium majus and the relationship to FHB disease development and mycotoxin accumulation from Fusarium graminearum infection. Plant Pathol., 56: 65-72.
Direct Link - Brunner, K., C.K. Peterbauer, R.L. Mach, M. Lorito, S. Zeilinger and C.P. Kubicek, 2003. The Nag1 N-acetylglucosaminidase of Trichoderma atroviride is essential for chitinase induction by chitin and of major relevance to biocontrol. Curr. Genet., 43: 289-295.
CrossRefDirect Link - Chala, A., J. Weinert and G.A. Wolf, 2003. An integrated approach to the evaluation of the efficacy of fungicides against Fusarium culmorum, the cause of head blight of wheat. J. Phytopathol., 151: 673-678.
Direct Link - Demeke, T., R.M. Clear, S.K. Patrick and D. Gaba, 2005. Species-specific PCR based assays for the detection of Fusarium species and a comparison with the whole seed agar plate method and trichothecene analysis. Int. J. Food Microbiol., 103: 271-284.
Direct Link - Draper, M.A., B. Bleakley, K.R. Ruden, S.M. Thompson and D.S. Wittmeier, 2005. Uniform trials for the performance of biological control agents in the suppression of Fusarium head blight in South Dakota. Proceedings of the National Fusarium Head Blight Forum, Milwaukee, WI, December 11-13, 2005, US Wheat and Barley Scab Initiative, Michigan State University, East Lansing, MI., pp: 189-189.
- Edwards, S.G., S.R. Pirgozliev, M.C. Hare and P. Jenkinson, 2001. Quantification of trichothecene-producing Fusarium species in harvested grain by competitive PCR to determine efficacies of fungicides against Fusarium head blight of winter wheat. Applied Environ. Microbiol., 67: 1575-1580.
CrossRefDirect Link - Gathumbi, J.K., E. Usleber and E. Mrtlbauer, 2001. Production of ultrasensitive antibodies against aflatoxin B1. Lett. Applied Microbiol., 32: 349-351.
CrossRef - Goyarts, T., S. Dänicke, H. Valenta and K.H. Ueberschär, 2007. Carry-over of Fusarium toxins (deoxynivalenol and zearalenone) from naturally contaminated wheat to pigs. Food Additives and Contaminants, 24: 369-380.
Direct Link - Halley, S., E.G. Van and V. Hofman, 2005. Effect of nozzles on fungicide efficacy for control of Fusarium Head blight on barley. Proceedings of the National Fusarium Head Blight Forum, Milwaukee, December 11-13, 2005, US. Wheat and Barley Scab Initiative, Michigan State University, East Lansing, pp: 1-1.
- Heier, T., S.K. Jain, K.H. Kogel and J. Pons-Kühnemann, 2005. Influence of N-fertilization and fungicide strategies on Fusarium head blight severity and mycotoxins content in winter wheat. J. Phytopathol., 153: 551-557.
Direct Link - Jochum, C.C., G.Y. Yuen and B. Tisserat, 2004. Effects of induced systemic resistance-activating agents on Fusarium head blight. Proceedings of the National Fusarium Head Blight Forum, Milwaukee, December 11-13, 2004, US. Wheat and Barley Scab Initiative, Michigan State University, East Lansing, MI, pp: 211-214.
- Kolombet, L.V., A.A. Starshov and D.A. Schisler, 2005. Greenhouse testing Trichoderma asperellum GJS 03-35 and yeast Cryptococcus nodaensis OH 182.9 as biocontrol agents against Fusarium head blight of wheat. Mycol. Phytopathol., 5: 80-88.
Direct Link - Lutz, M.P., G. Feichtinger, G. D'efago and B. Duffy, 2003. Mycotoxigenic Fusarium and deoxynivalenol production repress chitinase gene expression in the biocontrol agent Trichoderma atroviride P1. Applied Environ. Microbiol., 69: 3077-3084.
Direct Link - Mesterhazy, A., J. Bartok and C. Lamper, 2003. Influence of wheat cultivar, species of Fusarium and isolate aggressiveness on the efficacy of fungicides for control of fusarium head blight. Plant Dis., 87: 1107-1115.
CrossRefDirect Link - Parry, D.W., P. Jenkinson and L. McLeod, 1995. Fusarium ear blight (scab) in small grain cereals. Plant Pathol., 44: 207-238.
Direct Link - Paul, P.A., P.E. Lipps and L.V. Madden, 2005. Relationship between visual estimates of Fusarium head blight intensity and deoxynivalenol accumulation in harvested wheat grain: A meta-analysis. Phytopathology, 95: 1225-1236.
Direct Link - Perello, A., M.R. Simon and A.M. Arambarri, 2002. Interactions between foliar pathogens and the saprophytic microflora of the wheat (Triticum aestivum L.) phylloplane. J. Phytopathol., 150: 232-243.
Direct Link - Pereyra, S.A., R. Dill-Macky and A.L. Sims, 2004. Survival and inoculum production of Gibberella zeae in wheat residue. Plant Dis., 88: 724-730.
CrossRefDirect Link - Pirgozliev, S.R., S.G. Edwards, M.C. Hare and P. Jenkinson, 2003. Strategies for the control of Fusarium head blight in cereals. Eur. J. Plant Pathol., 109: 731-742.
CrossRefDirect Link - Ramirez, L., S. Chulze and N. Magan, 2004. Impact of environmental factors and fungicides on growth and deoxinivalenol production by Fusarium graminearum isolates from Argentinian wheat. Crop Protect., 23: 117-125.
CrossRefDirect Link - Schunmacher, R., N. Stopacher, B. Reithner, M. Omann, S. Zeillinger and R. Krska, 2007. Peptaibol profiles in cultures of Trichoderma atroviride: Detection and characterization by LC-MS/MS. Poceedings of the 15th International Symposium on Modern Fungicides and Antifungal Compounds, May 6-10, Ramada Treff Hotel, Friendrichroda, Germany.
- Shaner, G. and R.E. Finney, 1977. The effect of nitrogen fertilization on the expression of slow-mildewing resistance in knox wheat. Phytopathology, 67: 1051-1056.
CrossRefDirect Link - Usleber, E., V. Renz, E. Martlbauer and G. Terplan, 1992. Studies on the application of enzyme immunoassays for the Fusarium mycotoxins deoxynivalenol, 3-acetyldeoxynivalenol and zearalenone. Zoonoses Public Health, 39: 617-627.
CrossRefDirect Link - Wisniewska, H. and K. Kowalczyk, 2005. Resistance of cultivars and breeding lines of spring wheat to Fusarium culmorum and powdery mildew. J. Applied Genet., 46: 35-40.
Direct Link - Zadoks, J.C., T.T. Chang and C.F. Konzak, 1974. A decimal code for the growth stages of cereals. Weed Res., 14: 415-421.
CrossRefDirect Link