
E. I. Eziashi
Nigerian Institute for Oil Palm Research (NIFOR), Plant Pathology Division, P.MB. 1030, Benin City, Edo State, Nigeria
I. B. Omamor
Nigerian Institute for Oil Palm Research (NIFOR), Plant Pathology Division, P.MB. 1030, Benin City, Edo State, Nigeria
E. A. Dimaro-Oruade
Nigerian Institute for Oil Palm Research (NIFOR), Plant Pathology Division, P.MB. 1030, Benin City, Edo State, Nigeria
L. A. Ogunkanmi
Department of Cell Biology and Genetics, University of Lagos, Akoka, Yaba, Lagos State, Nigeria
Plant Pathology Journal
Year: 2007 | Volume: 6 | Issue: 4 | Page No.: 324-329







ABSTRACT
Ceratocystis paradoxa, causal agent of oil palm sprouted seeds rot produced phytotoxin, which induced brown discoloration on healthy oil palm sprouted seeds. This was observed 48 h after immersion in 70 and 100% solutions harvested at 21, 28 and 35 days. Brown discoloration observed on the apex of the radicle extended down to the base. Dryness of the tissue appeared grey-black, six days post inoculation. Symptom of brown discoloration was first observed on sprouted seeds treated with lyophilized phytotoxin from C. paradoxa, 22 h post inoculation. This was at 50, 70 and 100% concentrations, followed by non-sterilized and sterilized phytotoxins at 70 and 100% concentration each, 48 h post inoculation. One Hundred percent infection was reached ten days post inoculation with lyophilized phytotoxin at 100% concentration. Brown discoloration on radicle was observed on the 3rd day with phytotoxins from T. species treatment against C. paradoxa phytotoxin. T. viride, T. polysporum, T. hamatum and T. aureoviride produced phytotoxins, reduced infection rates on sprouted seeds immersed in phytotoxin from C. paradoxa. Concentration of Phytotoxins from T. species treatments without C. paradoxa phytotoxin, did not record symptoms at all levels of concentrations.
PDF Abstract XML References
How to cite this article
E. I. Eziashi, I. B. Omamor, E. A. Dimaro-Oruade and L. A. Ogunkanmi, 2007. Control of Phytotoxin from Ceratocystis paradoxa Using Trichoderma Species Phytotoxins on Oil Palm (Elaeis quineensis Jacq.) Sprouted Seeds. Plant Pathology Journal, 6: 324-329.
DOI: 10.3923/ppj.2007.324.329
URL: https://scialert.net/abstract/?doi=ppj.2007.324.329
DOI: 10.3923/ppj.2007.324.329
URL: https://scialert.net/abstract/?doi=ppj.2007.324.329
INTRODUCTION
A soil borne fungus Ceratocystis paradoxa is causing black seed rot disease in oil palm (Elaeis quineensis Jacq.) sprouted seeds (Omamor, 1985). An outbreak of the disease caused a black rot of plumule and radicle of sprouted seeds in 1982 in a seed store at Nigerian Institute for Oil Palm Research (NIFOR), for sale to farmers. About 109,000 sprouted seeds were affected, resulting in a huge financial loss to the Institute (Omamor, 1985).
Typical symptoms of most plants diseases revealed the involvement of phytotoxic metabolites, which therefore suggest a role for toxic metabolites secreted by the pathogen in the disease development. Metabolites of many fungi may have adverse or stimulatory effects on plants (Heisey et al., 1985; Rice, 1995).
Pathogenic fungi and bacteria often damage their host plants by producing toxins, which cause various symptoms including necrosis, chlorosis, wilting, water soaking and eventually the death of plants (Sceffer, 1983). One criterion of the importance of a toxin in a disease syndrome caused by a pathogen is that toxigenicity is often related to pathogenicity or virulence (Yoder, 1980). Fungi can produce a wide range of toxic compounds with varied biochemical structures and modes of action. These include polypeptides, glycoproteins, aminoacid derivatives, polypeptides, terpenoids, sterols and quinines (Kono et al., 1981; Stoessl, 1981). Toxin production has been reported for several Trichoderma. The production of antibiotics: Ergokonin (Kumeda et al., 1994) viridin (Grove et al., 1995, 1996) and viridin fungin A, B and C (Harris et al., 1985) by Trichoderma viride have been involved in biological control.
The first objective of this study was to determine whether Ceratocystis paradoxa produced phytotoxin, could cause rot disease on oil palm sprouted seeds. The second objective was to determine the effects of phytotoxins from Trichoderma species for the control of C. paradoxa produced phytotoxin on oil palm sprouted seeds.
MATERIALS AND METHODS
Material and fungal isolates: Sprouted seeds of the oil palm used in these investigations were raised from germinated improved seed (Tenera hybrid) produced at the Nigerian Institute for Oil Palm Research (NIFOR). The seeds were sprouted by the heat treatment method (Anonymous, 1965).
The studies were conducted in 2006 at botany and microbiology laboratories, University of Lagos, Nigeria. The fungal cultures used in this investigation were previously isolated by the authors from diseased sprouted seeds of the oil palm collected from the above-mentioned location (NIFOR) in 2004. They were Trichoderma viride, T. hamatum, T. polysporum, T. aureoviride and the pathogenic fungus Ceratocystis paradoxa, the causal agent of black seed rot of sprouted oil palm seeds.
Phytotoxin extraction from culture medium: A small mycelial block (4 mm) was removed from each of the 7 day-old pure cultures of Trichoderma species and Ceratocystis paradoxa. Each was transferred to each of the 1 L Erlenmeyer cornical flask containing 500 mL, which contained dextrose 25.00 g, Ammonium tartrate 2.00 g, KH2PO4 2.00 g, MgSO4 1.00 g, Ferrous sulphate 0.01 g and water 1000.00 mL (Brian and Hemming, 1945). Inoculated media were incubated statically 28°C±2 for 35 days under light and dark. Filtrates were harvested by filtering first through a muslin cloth twice and through Whatman no 1 filter paper twice. The culture filtrate was again obtained by centrifugation at 5000 x g for 15 min. The filtrates (500 mL) was concentrated to 10-fold by lyophilization.
Experiments: Phytotoxin produced by C. paradoxa was used for testing the response of incubation and concentration on oil palm sprouted seeds for induction of symptoms. Cornical flasks (500 mL each) were harvested at 7, 14, 21, 28 and 35 days. Phytotoxin extracts were diluted with sterile-distilled water to give concentrations of 20, 50, 70 and 100% solution, respectively. Before the seeds were sprouted, they were soaked in 90% (vol./vol.) ethanol for 2 min and washed five times with sterile distilled water. Treated seeds were transferred to sterile filter paper to absorb excess water and then disinfected in 2% (wt./vol.) sodium hypochlorite (Sigma-Aldrich Chenire) for 2 min (Yedidia et al., 1999). Sprouted seeds selected with the same uniform lengths (newly developed radicles) were immersed in different solutions for 1 h to test the biological activity. The control-sprouted seeds were immersed in non-inoculated medium. They were incubated for six days.
In order to determine the response of autoclaving (121°C for 15 min) and lyophilization on symptom development on oil palm sprouted seeds. The medium was harvested after 28 days. Each flask (500 mL) of the control (non-inoculated medium), sterilized, non-sterilized and lyophilized phytotoxin (500 mL to 10-fold mL) was each diluted to give concentrations of 20, 50, 70 and 100%, respectively. The sprouted seeds were immersed in these solutions. They were incubated for 14 days.
Sterilized phytotoxins produced by Trichoderma species and C. paradoxa (500 mL each) were investigated, first by immersion of sprouted seeds in each of T. species phytotoxins at different concentrations (20, 50, 70 and 100%) for 1 h and later immersed in C. paradoxa phytotoxin for 1 h. Control, had phytotoxin from C. paradoxa without phytotoxins from T species. They were incubated for 14 days. Ten sprouted seeds replicated 4 times for each treatment. % Infection = Number of infected/Total Number of sprouted seedsx% (Omamor et al., 2000). All the treatments were incubated at 28°C±2 and repeated twice.
RESULTS
Toxicity of different ages and concentration of C. paradoxa induced light brown coloration on the radicle apex of the oil palm sprouted seeds (Fig. 1A). The brown spots were observed 48 h after immersion in 70% and 100% solutions harvested at 21, 28 and 35 days. The brown discoloration that was first observed on the apex of radicle extended down to the radicle base. This resulted in complete dryness of the tissue, which appeared dark brown six days post inoculation (Fig. 1B). No brown spots were observed from the control treatment (Fig. 1C). Phytotoxins harvested at different ages showed some differences. Significant differences (p<0.5) were observed among phytotoxins harvested at 21, 28 and 35 days when compared with 7 and 14 days (Table 1). Phytotoxins harvested on the 7 and 14 days did not record symptoms at 20, 50, 70 and 100% concentrations 48 h post inoculation. The control treatment did not record symptoms at all level of concentrations.
There were no significant differences between sterilized and non-sterilized phytotoxin. Sterilized phytotoxin was weakened slightly by heat (Table 2), but significant difference was obvious on lyophilized phytotoxin when compared. Symptoms of infections were first observed on sprouted seeds treated with lyophilized, followed by non-sterilized and sterilized phytotoxins two days post inoculation at 50, 70 and 100% concentrations, respectively. One hundred percent infections on sprouted seeds was reached 10 days post inoculation with Fig. 1 (A-C):Oil palm sprouted seeds. The arrow heads are showing infections from the apex and the gradual invasion of the tissue 48 h after immersion (1A). Diseased oil palm sprouted seeds, six days after immersion in phytotoxin from Ceratocystis paradoxa, showing complete invasion from the apex to the base with brown shrinked tissues (1B). Healthy oil palm sprouted seeds, the radicle and plumule remained uninfected six days post inoculation with sterile water (1C) lyophilized phytotoxin at 100% concentration.
 | |
| Fig. 1: | oil Palm sprouted seeds. |
| Table 1: | Response of phytotoxin produced from C. paradoxa at different ages and concentrations on oil palm sprouted seeds |
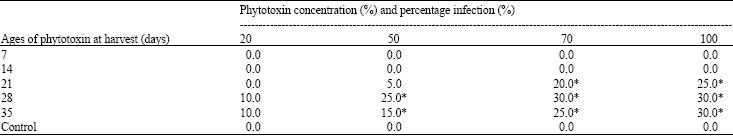 | |
| Mean values followed by * in the same column are significant (p<0.05) according to one-way analysis of variance as compared to the control. Values were averages of 4 replicate | |
| Table 2: | Response of heat and lyophilization on phytotoxin produced by C. paradoxa on oil palm sprouted seeds |
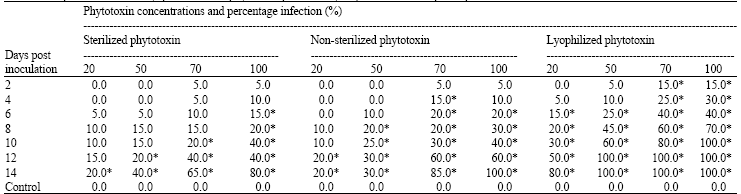 | |
| Mean values followed by * in the same column are significant (p<0.05) according to one-way, analysis of variance as compared to the control. Values were averages of 4 replicates | |
Only 80 and 60 infections were reached by non-sterilized and sterilized phytotoxins 14 days post inoculation at 100% concentration. In the experiment, there were slight significant differences between concentrations of 70% and 100%, but there were significant differences (p<0.5) when compared with concentrations at 20 and 50% respectively (Table 2).
Symptoms on infested sprouted seeds appeared on the third day at all the concentrations of Trichoderma species treated (with phytotoxin from C. paradoxa) sprouted seeds, except T. viride (Table 3). Although, there were no significant differences. Phytotoxin concentrations (20, 50, 70 and 100%) from Trichoderma species (without phytotoxin from C. paradoxa) treated sprouted seeds, did not record any symptom of brown discoloration 14 days post inoculation. However, on the 14 day post inoculation, there were slight significant differences (p<0.5) at all the concentrations (20, 50, 70 and 100%) of Trichoderma species treated (with phytotoxin from C. paradoxa) sprouted seeds (Table 3).
| Table 3: | Response of phytotoxins produced by Trichoderma species against phytotoxin produced by C. paradoxa on oil palm sprouted seeds |
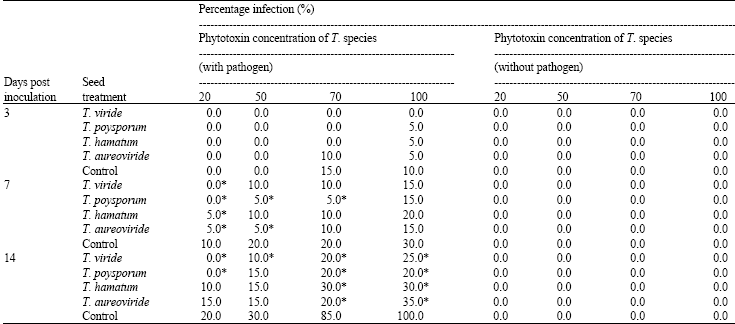 | |
| Mean values followed by * in the same column are significant (p<0.05) according to one-way analysis of variance as compared to the control. Values were averages of 4 replicates | |
DISCUSSION
In this study, C. paradoxa produced phytotoxin in antibiotic fermentation medium. Production of phytotoxin from C. paradoxa reached a peak after 28 days of static incubation at 28°C±2. The effect of inoculum concentration on the time needed for maximum toxin production has previously been described for the enniantins (Rudolph, 1976). Symptoms on oil palm sprouted seeds caused by phytotoxin produced by C. paradoxa were similar to symptoms produced by C. paradoxa (spores) infested on oil palm sprouted seeds. (Omamor, 1985) reported that sprouted seeds inoculated with C. paradoxa (spores) changed from light brown to brown and black. In this study, sprouted seeds inoculated with phytotoxin from C. paradoxa changed from brown coloration to dark brown. This is likely to be as a result of the red pigment produced by C. paradoxa during the period of static incubation. Red pigment produced by C. paradoxa in antibiotic fermentation medium, suggested that the pigment might have toxigenic activity. This coincided with the hypothesis that, isolates, which produced red pigments in liquid culture, were more efficient in producing biological active culture filtrate (Duarte and Archer, 2003). Scheffer (1983), reported that pathogenic fungi and bacteria often damage their host plants by producing toxins, which cause various symptoms including necrosis, chlorosis, wilting, water soaking and eventually the death of plants.
Infested oil palm sprouted seeds showed symptoms of brown coloration after immersion in sterilized, non-sterilized and lyophilized phytotoxins. Slight reduction of biological activity was observed in sterilized phytotoxin, this suggested that some substances might be weakened by sterilization but the toxic components remained active. This agreed with the hypothesis of Metha and Brogin (2000) which observed that culture filtrate produced by Stemphylium solani Weber was shown to be stable during autoclaving, however, the 10-fold concentration of phytotoxin using lyophilizer did not ten-fold increase in activity, compare with the original phytotoxin that was not lyophilized. This suggested that the concentration process under lyophilization might have inactivated some of the phytotoxins. Thermostability has been observed for many host-specific and non-host specific toxins (Hartman et al., 1984; Patel et al., 1987).
This study showed that infested oil palm sprouted seeds with phytotoxins from T. species reduced infection of phytotoxin from C. paradoxa on oil palm sprouted seeds. Both Trichoderma species and C. paradoxa produced phytotoxins, responded to different concentration treatments on oil palm sprouted seeds. Each of the T. species produced phytotoxin, treated against phytotoxin from C. paradoxa reduced infections on oil palm sprouted seeds when compared with the control treatment. On the 14 day post inoculation, T. viride and T. polysporum produced phytotoxins, reduced percentage infections (phytotoxin concentration of T. species with pathogen) when compared with T. hamatum, T. aureoviride and control. Eziashi et al. (2006a, b) reported that, T. viride and T. polysporum (spores) reduced germination failure on oil palm sprouted seeds treated with C. paradoxa (spores). However, there was no infection 14 day post inoculation (phytotoxin concentration of T. species without pathogen) with sprouted seeds treated with T. species alone. This suggested that T. species did not cause infection.
The mechanisms of actions by Trichoderma species must have been associated with the production of metabolite compounds. Eziashi et al. (2006b) reported that the growth of C. paradoxa was inhibited in vitro by Trichoderma species during volatile, non-volatile and direct-diffusible metabolite tests. Similarly, fungi and bacteria synthesize a number of proteins and peptides that are antifungal (Chae et al., 1999; De Lucca and Waish 1999; Iijima et al., 1994; Kimura et al., 1999; Levitz, 1992). The metabolite compounds and structures involved in this study are yet to be determined. Phytotoxin from C. paradoxa caused brown rot infection on oil palm sprouted seeds while phytotoxins from T. viride and T. polysporum exhibited better control of phytotoxin from C. paradoxa.
REFERENCES
- De Lucca, A.J. and T.J. Waish, 1999. Antifungal peptides: Novel therapeutic compounds against emerging pathogens. Antimicrob. Agents Chemomother., 43: 1-11.
Direct Link - Eziashi, E.L., N.U. Uma, A.A. Adekunle and C.E. Airede, 2006. Effect of metabolites produced by Trichoderma species against Ceratocystis paradoxa in culture medium. Afr. J. Biotechnol., 5: 703-706.
Direct Link - Eziashi, E.I., N.U. Uma, A.A. Adekunle and I.B. Omamor, 2006. Biological control of Ceratocystis paradoxa causing black seed rot in oil palm sprouted seeds by Trichoderma species. Pak. J. Biol. Sci., 9: 1987-1990.
CrossRefDirect Link - Metha, Y.R. and R.I. Brogin, 2000. Phytotoxity of a culture filtrate produced by Stemphylium solani of cotton. Plant Dise., 84: 838-842.
Direct Link - Yedidia, I., N. Benhamou and I. Chet, 1999. Induction of defense responses in cucumber plants (Cucumis sativus L.) By the biocontrol agent Trichoderma harzianum. Applied Environ. Microbiol., 65: 1061-1070.
PubMedDirect Link - Grove, J.F., J.S. Moffatt and E.B. Vischer, 1965. Viridin. Part I. Isolation and characterization. J. Chem. Soc., 1965: 3803-3811.
CrossRef