
Heidi I.G. Abo-Elnaga
Department of Plant Pathology, Faculty of Agriculture, Assiut University, Assiut, Egypt
Naglaa G. Ahmed
Institute of Plant Pathology, Agriculture Research Center, Giza, Egypt
Plant Pathology Journal
Year: 2007 | Volume: 6 | Issue: 4 | Page No.: 318-323







ABSTRACT
The objective of present research is to study the pathogenic capabilities of two isolate Fusarium moniliforme and the production of Fumonisin, Zearalenon and gibberellic acid in relation to the isolate virulence. Fusarium moniliforme (Sheldom) was isolated from naturally infected roots of different pepper plants. Pathogenicity tests indicated that Fusarium moniliforme was more pathogenic to lang red and cagenne (hot pepper) than to the local pepper (sweet pepper), California Wonder (sweet pepper) and California (sweet pepper) under greenhouse conditions. The two isolates differed in their capability of inducing the infection. Different quantities of each of fumonisin, zearelonen and gibberellic acid were produced by the tested isolates of Fusarium moniliforme in vitro. The infected pepper plants were evaluated for resistance to Fusarium moniliforme and Fumonisin Zearelonen, concentration.
PDF Abstract XML References
How to cite this article
Heidi I.G. Abo-Elnaga and Naglaa G. Ahmed, 2007. Pathogenicity, Toxicity and Gibberellic Acid Content of Fusarium moniliforme Causing Root Rot and Damping Off of Pepper. Plant Pathology Journal, 6: 318-323.
DOI: 10.3923/ppj.2007.318.323
URL: https://scialert.net/abstract/?doi=ppj.2007.318.323
DOI: 10.3923/ppj.2007.318.323
URL: https://scialert.net/abstract/?doi=ppj.2007.318.323
INTRODUCTION
Pepper (Capsicum annum) is one of the vegetables that spreads and is grown world wide as a spice and a medical crop. It is eaten raw and cooked immature and mature.
Pepper is considered to be an excellent source of vitamin A, B1, B2, B3, C and calcium. Pepper is subject to be attacked by many diseases in Egypt as well as in many different parts of the world (Lee et al., 1990; Kim et al., 1990; Saimmaime et al., 1991; Gonzalez et al., 1998; Harris, 1999; Goicoechea et al., 2000; Ivey and Miller, 2000; Kobori et al., 2000). Damping off disease caused by some Fusarium species is considered to be one of the most destructive diseases and serious diseases in many parts of the world as well as in Egypt (Uma, 1981; Sarhan and Hegazi, 1988; Freire et al., 2000; Utkhede and Mathur, 2003; Nutsugah et al., 2004). Infection of pepper plants with Fusarium species can cause contamination of pepper fruit with mycotoxins that effect human health (Desjardins et al., 2000; Leslie et al., 1992; DeNijs and Notermans, 1996; Desjardins et al., 1997; Abbas et al., 1998; Giridhar and Reddy, 1999; Douglas et al., 1999; Bagi et al., 2000; Desjardins et al., 2000; Freire et al., 2000; Machado and Kemmel-Meier, 2001; Rodriguez-Amaya and Sabino, 2002; Antonio et al., 2003; Clements and Kleinschmpdt, 2003; Bush et al., 2004). The objective of this research is to study the pathogenic capabilities of two isolates Fusarium moniliforme and the production of fumonisin, zearalenone and gibberellic acid in relation to the isolate virulence.
MATERIALS AND METHODS
Naturally infected seedlings of local pepper varieties showing damping off were obtained from several fields from different localities of Assiut Governorate Egypt (Assiut, Badary). Isolation procedures of the caused pathogen were carried out using small pieces of infected roots which were washed in running tap water, surface sterilized with 1% sodium hypochlorite solution for two minutes washed in three changes of sterilized water and dried between sterilized filter papers. The surface sterilized root pieces were then placed on PDA medium in petri dishes and incubated at 25°C for seven days. Preliminary identification of the isolated fungi was carried out according to Booth (1997) and Moubasher (1993) and confirmed by Assiut University Mycological Center (AUMC).
The isolated fungi were tested for their pathogenic capabilities on California wonder (Sweet pepper) California Lang Red, Cagenne (Hot pepper) and Local (Sweet pepper) during 2005 season under greenhouse conditions. Earthen pots (23 cm diameter) filled with loamy and soil (84% sand, 14.5% clay and 1.5% silt) were used. Each pot was filled with 3 kg sterilized (autoclaved) soil. Four pots were used for each tested variety. Four replicates were used for each isolate. The pots were inoculated with the fungal inoculum, grown on barley grains medium, 7 days before sowing at the rate of 5 g pot-1 and mixed thoroughly with the filling soil. Five surface sterilized pepper seeds were sown in each pot for each variety after disinfection with 2% sodium hypochlorite solution for two minutes.
Pots containing non-infested soil mixed with 5 g of sterilized barely medium were used as a control. The pots were irrigated when necessary and examined daily. The percentage of pre-and post emergence damping off was recorded after 21 days and 45 days, respectively. Disease index was recorded after the end of the experiment by using the scale of 0 to 4 used by AICRIP (1968). Re-isolation of the pathogenic fungal isolates from the infected plants was carried out to meet Koch postulates.
Determination of Fusarium mycotoxins
Chemical analysis of fungal culture: The production of fumonisin and zearalenone was assayed in a culture grown on a yeast medium for seven days at temperature 25°C±2.
Fumonisin analysis: Fumonisin toxins produced was determined as follows: 50 g of culture material (biomass) with 5 g salt and 100 mL methanol water (80:20) was blended at high speed for 1 min, then filtered through fluted filter paper. Ten milliliter of the filtrate was diluted with 40 mL of wash buffer and filtered again through 1.0 Um micro-fiber filter. Ten milliliter of the diluted extract was passed through fumontest column (Vicam Company) and then column was washed by 10 mL of the same dilute solution. The fumonisin determined by High Performance Liquid Chromatography (HPLC) was eluted by passing 1 mL of HPLC of orthophthaladehyde derivatives as reported by Nelson et al. (1993) and grade methanol through the column and elutes were re-collected again. One milliliter of developer A (Vicam product No. G 5005) and developer B (Vicam product No. G5004) was added to the elute and placed in calibrated fluorometer (Series-4/Vicam). The fumonisin concentration was measured after 4 min according to the method described by Nazzani et al. (2001).
Zearalenone analysis: Zearalenone concentration was determined as mentioned in the fumonisin toxins determination method but the dilution was made with 49 mL distilled water which was passed through zearalest column (Vican Company) and then measured in a calibrated fluorometer model Series-4/Vicam according to Martins and Bernardo (2003).
Chemical analysis of fungal culture: The production of Gibberellic Acid (GA) was assayed in a culture grown on Elliott’s solution medium for chemical analyses. The entire contents of each of culture material 10 g was extracted with 70% methanol and stirred overnight at 4°C. The extract was filtered through a whattman filter paper and the methanol was evaporated in vacuo. The aqueous phase was adjusted to pH 8.5 with a 0.1 M phosphate buffer and then partitioned three times with ethyl acetate. After removal of the ethyl acetate phase the aqueous phase was adjusted to pH 2.5 with 1 N HCl. The solution was partitioned three times with ethyl acetate. After removal of ethyl acetate phase, the aqueous phase was adjusted to pH 2.5 with 1 N HCl. The solution was partitioned three times with diethyl ether and then passed through water less sodium sulphate. After the diethyl ether was evaporated in vacuo, the dry residua containing hormones was dissolved in methanol and stored in vials at 4°C (Chen, 1990).
HPLC analysis was used to study the growth hormones in extracts dissolved in 1 mL grade methanol. Analysis of GA was performed on a model hp 1050 HPLC equipped with UV detector. Separations and determinations were performed on a Cl8 column (4.6x150 mm). The mobile phase yielded results of 30% methanol (adjusted to pH 3.0 with 0.1 M H3P04) for GA, Wavelengths in the UV detector were 208 nm. Total run time for separations was approximately 5 min at a flow rate of 1 mL min-1 according to (Baydar and Ulger, 1998).
Mycotoxin analysis of pepper root samples
Fumonisin and zearalenone: Selected pepper root samples 50 g of ground sample (biomass), with 5 g slat and 100 mL methanol water (80: 20), was blended at high speed for 1 min. The following determination steps were carried out mentioned before in the chemical analysis of the fungal cultures.
Statistical analysis: The collected data were statistically analyzed using two factor Completely Randomized Block Design. Treatments were compared at 0.05 and 0.01 level of probability LSD (Fisher, 1984).
RESULTS AND DISCUSSION
Isolation and identification of the causal pathogen: Identification procedures of the isolated fungi proved the presence of two Fusarium monilifrome isolates (Pepper varieties Califorina (sweet pepper), Califorina wonder (sweet pepper), local pepper (sweet pepper) and lang red cagenne (hot pepper) were tested in this study).
 | |
| Fig. 1: | A, B, C and D Symptoms on different varieties of pepper inoculated with Fusarium moniliforme are similar in reducing the growth of roots while each variety showed different symptoms as mentioned |
The data shown in Table 1 and Fig. 1 show that all tested pepper varieties were susceptible to infection by the two Fusarium isolates. The fungal isolates varied in their virulence on the tested pepper varieties. The obtained data revealed that there is a significant difference between the testes isolated either in the percentage of pre, post emergence, survival and the disease index. It also show that isolate 2 was more virulent in the case of the percentage of pre and post emergence, survival and the disease index. The variability in the isolates, severity could be attributed to differences gibberellic acid and toxins producing capability. These results agree with the observation of Leslie et al. (1992), De Nijs and Notermans (1996), Desjardins et al. (1997), Abbas et al. (1998), Giridhar and Reddy (1999), Desjardins et al. (2000), Rodriguenz-Amaya and Sabno (2002) and Bush et al. (2004).
On the other hand the tested pepper varieties varied markedly in their resistance and suscepability to infection. California (sweet pepper) and California wonder (Sweet pepper) were resistant to infection compared to local (sweet pepper) and lang Red Cagenne (hot pepper) varieties as reported by Rivelli (1989) and Sanogo (2003).
The resistance and susceptibility to infection with Fusarium isolates and have also been previously mentioned by Uma (1981) Sarhan and Hegazi (1988), Utkhede and Mathur (2003) and Nutsugah et al. (2004) observed have among the tested varieties may be due to the various defense mechanisms available to these varieties.
| Table 1: | Pathogenicity test of Fusarium moniliforme isolates causal pathogen of damping off and root rot in four varieties of pepper plants |
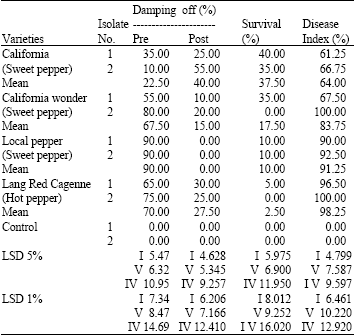 | |
| I = Isolates, V = Variety, IV = Interaction isolates, x variety | |
Determination of fumonisin and zearalenone mycotoxin in Fusarium moniliforme isolates and in diseased pepper tissues with Fusarium moniliforme: In order to study role of the mycotoxin on the pathogenic process determination procedure was carried out using the two tested isolates of Fusarium moniliforme. The presented data in Table 2 indicates that isolate II, inserted more fumonisin when compared with that isolate 1. This may indicate the difference in their pathogenic capabilities which may due to the mycotoxin fumonisin. These results agree with the findings of Jimenez et al. (2000) who reported that there are differences among Fusarium isolates in the producing of fumonisin. Other workers mentioned that there is a correlation between the ability of the causal pathogen to insert fumonisin in vivo and the severity of maize seedling blight disease in vivo (Desjardins et al., 1995).
Data presented in Table 2 also show that the two tested Fusarium isolates produced zearalenone under laboratory conditions. The production of zearalenone in Fusarium culture was confirmed by Machado and Kemmel-Meier (2001).
Data in Table 2 also showed that the infected pepper tissues contained more Zearalenone when infected with isolate II, compared to isolate I. Which it was free from fumonisin in tissues infected with both tested isolates. These results indicate that there is a correlation between the severity of infection with Fusarium isolate and their production of Zearalenone in plant tissues. This may be due to that Zearalenone is generally considered to be inhibitory of root and shoot elongation and fresh mass accumulation (Mclean, 1995).
| Table 2: | Determination of mycotoxin in two isolates of Fusarium moniliforme and in diseased pepper tissues with Fusarium moniliforme |
 | |
| Table 3: | Determination of gibberellic acid in two isolates of Fusarium moniliforme |
This findings agree with Bagi et al. (2000), Bata et al. (2001), Antonio et al. (2003) and Clements et al. (2003). There are no references concerning pepper plant at hand but there are many workers who worked on other different crops, wheat, maize and grain.
Determination of Gibberllic acid in two isolates of Fusarium moniliforme. Results presented in Table 3 show that the tested isolates varied in their production of gibberellic acid. Isolate I which proved to be less pathogenic than isolate II inserted more gibberellic acid than isolate II. The variability in pathogenic capability of the two isolates may be due to the difference in their ability to insert gibberellic acid which is well know to be an antifungal substance (Gowily-Ahlam, 1987; El-Ganaiemy et al., 2002).
REFERENCES
- Bata, A., P. Rafai and G. Kovacs, 2001. Investigation and a new evaluation method of resistance of maize hybrids grown in Hungary to Fusarium moulds. J. Phytopathol. Phtopathologische Zeitschrifit, 149: 107-111.
Direct Link - Bush, B.J., M.L. Carson, M.A. Cubeta, W.M. Hagler and G.A. Payne, 2004. Infection and Fumonisin production by Fusarium verticillioides in developing maize kernels. Phytopathology, 94: 88-93.
Direct Link - Chen, W.S., 1990. Endogenous growth substances in xylem and shoot tip diffusate of lychee in relation to flowering. Hortic. Sci., 25: 314-315.
Direct Link - Clements, M.J. and C.E. Kleinschmidt, 2003. Evaluation of inoculation, techniques for Fusarium ear rot and Fumonisin contamination of corn. Plant Dis., 87: 147-153.
Direct Link - Delia, B., Rodriguez-Amaya and M. Sabino, 2002. Mycotoxin research in Brazil: The last decade in review. Brazilian J. Microbiol., 33: 1517-1529.
CrossRefDirect Link - Desjardins, A.E., R.D. Manandhar, G.G. Manandhar, S.M. Poling and C.M. Maragos, 2000. Fusarium species from Nepalese rice and production of mycotoxins and gibberellic acid by selected species. Applied Environ. Microbiol., 66: 1020-1025.
Direct Link - El-Ganaieny, R.M.A., H.Y. Mohamed, A.A. Omran and N.M. Abdel-Gawad, 2002. Methods to reduce the peanut damping off, root and pod rot diseases and their damage in newly reclaimed lands of Beni-Suef governorate. Proceedings of the 3rd Scientific Conference Agricultural Science, Assiut., October 15-17, 2002, Egypt, pp: 267-284.
- Freire, F.C.O., Z. Kozakiewicz and R.R.M. Paterson, 2000. Mycoflora and mycotoxins in Brazilian black pepper, white pepper and Brazil nuts. Mycopathology, 149: 13-19.
Direct Link - Ivey, M.L.L. and S.A. Miller, 2000. First report of bacterial cankers of pepper in Ohio. Plant Dis., 84: 810-810.
CrossRefDirect Link - Jimenez, M., S. Rodriguez, J.J. Mateo, J.V. Gil and R. Mateo, 2000. Characterization of Gibberella fujikuroi complex isolates by fumonisin fujikuroi complex isolates by fumonisin B1 and B2 analysis and by RAPD and restriction analysis of PCR-amplified internal transcribed spacers of ribosomal DNA. Systemic Applied Microbiol., 23: 45-555.
Direct Link - Kobori, R.F., P.T.D. Vecchia, C. Kurozawa and J.M.P. Soler, 2000. Evaluation of the resistance Capsicum annuum lines when inoculated with three inoculum concentrations of Phytophthora capsici Summa. Summa Phytopathol. Brazil, 26: 77-81.
Direct Link - Leslie, J.F., R.D. Plattner, A.E. Desjardins and C.J.R. Klittich, 1992. Fumonisin B1 Production by strains from different mating populations of Gibberella fujikuroi (Fusarium moniliforme). Phytopathology, 82: 341-345.
Direct Link - Martins, M.L., H.M. Martins and F. Bernardo, 2003. Fungal flora and mycotoxin detection in commercial pet food. Revista Portugusesa de Ciencias Veterinarias, 48: 179-183.
Direct Link - Mazzani, C., O. Borges, O. Luzon, V. Barrientos and P. Quijada, 2001. Occurrence of Fusarium moniliforme and fumonisins in kernels of maize hybrids in venezuela. Brazilian J. Microbiol., 32: 345-349.
CrossRefDirect Link - Nutsugah, S.K., L. Vibeke, I.D.K. Atokple and F.K. Ayensu, 2004. Seed-borne Mycoflora of major food crops in Ghana. J. Sci. Technol. (Ghana), 24: 22-31.
Direct Link - Utkhede, R. and S. Mathur, 2003. Fusarium fruit rot of greenhouse sweet peppers in Canada. Plant Dis., 87: 100-100.
CrossRefDirect Link - Logrieco, A., A. Bottalico, G. Mule, A. Moretti and G. Perrone, 2003. Epidemiology of toxigenic fungi and their associated mycotoxins for some Mediterranean crops. Eur. J. Plant Pathol., 109: 645-667.
CrossRef