
M. Abrinbana
Department of Plant Protection, Faculty of Agriculture, University of Urmia, Urmia, Iran
A. Babai-Ahary
Department of Plant Protection, Faculty of Agriculture, University of Tabriz, Tabriz, Iran
I. Madjidi Heravan
Agricultural Biotechnology Research Institute, Karaj, Iran
Plant Pathology Journal
Year: 2007 | Volume: 6 | Issue: 3 | Page No.: 266-270







ABSTRACT
To evaluate the resistance in seedlings of 20 sugarbeet lines against two isolates of P. ultimum var. ultimum under greenhouse conditions, seeds were sown in sterilized soil treated with certain amounts of P. ultimum inoculum. Seeds of the line Rhizofort had already been treated with a fungicidal substance containing iprodion and hymexazol. The resulting seedlings were evaluated 3 week after sowing date. A zero to five rating system was applied to assess different levels of resistance. As a result of this evaluation, 16 lines were grouped as very susceptible and susceptible. The remaining 4 lines including 9597-P.58, 8150-Bulk, Mst231 and MstC2 appeared to be resistant to the infection. Also the line Rhizofort was grouped as the resistant because of its healthy appearance which might be due to the fungicidal treatment of the seeds. Results from this experiment revealed that the efficacy of resistance in the lines against damping-off, was more than the fungicidal treatment of the seeds.
PDF Abstract XML References
How to cite this article
M. Abrinbana, A. Babai-Ahary and I. Madjidi Heravan, 2007. Assessment of Resistance in Sugarbeet Lines to Damping-Off Caused by Pythium ultimum Trow var. ultimum Under Greenhouse Conditions. Plant Pathology Journal, 6: 266-270.
DOI: 10.3923/ppj.2007.266.270
URL: https://scialert.net/abstract/?doi=ppj.2007.266.270
DOI: 10.3923/ppj.2007.266.270
URL: https://scialert.net/abstract/?doi=ppj.2007.266.270
INTRODUCTION
Germinating seeds of sugarbeet (Beta vulgaris L.) and other crops are susceptible to seed rot and damping-off caused by Pythium ultimum Trow. The pathogen is ubiquitous and can reduce stand establishment (Babai-Ahary et al., 2004). Present survey in the northwest Iran indicated that P. ultimum var. ultimum is the dominant species among Pythium sp. in sugarbeet fields that are capable of causing damping-off (Babai-Ahary et al., 2004).
Where seedling damping-off diseases are economically important, control is achieved almost solely by fungicides applied to the surface of seeds. But, the most effective fungicides (e.g., hymexazol) should provide useful protection from pathogens under moderate disease pressure (Payne and Williams, 1990).
With the greater awareness of the environmental impact and cost of pesticides, more attention is being paid to improving genetic resistance to the damping-off pathogens. Differences in resistance to damping-off caused by P. ultimum in Beta species reported, but the number of resistant sugarbeet cultivars available to growers is limited at present (Luterbacher et al., 2000, 2005).
Currently, there is no information available on the resistance in existing sugarbeet cultivars and lines in Iran. Therefore, in this study, we evaluated 20 sugarbeet lines for resistance to the damping-off in soils on the basis of ED50 values of two P. ultimum var. ultimum isolates under greenhouse conditions.
MATERIALS AND METHODS
Fungal isolates: Two isolates of P. ulimum var. ultimum including Pu8 and Pu2, were used for assessing the resistance. These isolates which were already isolated from the sugarbeet fields of the northwest region of Iran possessed the highest pathogenicity among the isolates (Babai-Ahary et al., 2004).
Sugarbeet lines: Twenty lines of sugarbeet (Beta vulgaris L. subsp. vulgaris) were used in this evaluation along with the line Elit 7233 as the control (Table 4). Seeds of the line Rhizofort were already treated with a fungicidal substance containing hymexazol and iprodion. For each line, 10 seeds were sown in two infested and non-infested (control) pots.
Preparation of inoculum: Inoculum was prepared according to Martin (1992). Briefly, the isolates were grown in 9 cm petri dishes containing PDB (potato dextrose broth). A PDA (potato dextrose agar) disk, 1 cm in diameter, obtained from the edge of 2-day-old colonies of Pu8 and Pu2 were placed in each Petri dish. After 48 h of inoculation at 22±1°C in darkness, the medium was decanted from each Petri dish and the remaining mycelia were induced according to Nelson and Craft (1989) to produce hyphal swellings. The mycelia were then added into the plastic pots containing sterile soil using mycelia of 100 Petri-dishes per kg of the soil. To prevent soil contamination, all plastic pots were capped and placed at 25-30°C for 7-10 days to dry the contents of the pots. The contents of plastic pots were used as inoculum following drying and grinding.
Cfu determination of inoculum: Soil drop technique was used to determine colony forming units (cfu) as described by Stanghellini and Hancock (1970). To do so, the dilution series of 1:10, 1:50, 1:100, 1:200, 1:500, 1:1000 and 1:10000 soil suspensions were prepared in sterilized distilled water and each dilution was shaken for 2 min on a test tube shaker. Then 1 mL of each suspension was removed immediately and pipetted dropwise on the surface of 3-day-old 2% water agar. Each milliliter was dispensed in 40 drops, 10 drops/plate. One drop was placed in the center and nine drops in the periphery of the agar plate. The dishes were then incubated at 24±1°C for 24 h in darkness. The drops on water agar were examined with a microscope (40x magnification) and the numbers of hyphae emerging from any of the drops were counted. This experiment was repeated three times.
ED50 determination of the isolates: ED50 (50% effective dosage) determination of the isolates was done on Elit7233 which has been shown to BE susceptible to seedling damping-off (Babai-Ahary et al., 2004). To do so, steril soil was infested with several concentrations of hyphal swellings of P. ultimum var. ultimum according to logarithmic intervals (20, 35, 65, 120, 220 and 440 CFU). Inocula were mixed with steril soil and each concentration was then distributed into four pots (12 cm deep, 10 cm top diameter) and non-infested (steril) soil was used as the control. Sugarbeet seeds were surface-disinfested using 0.5% NaOCl solution for 1 min and ten seeds were then sown in 2 cm depth in each pot. The pots were kept in the greenhouse at 22±2°C, 70±5 RH and 12 h light/dark for 3 week. To provide the required moisture, all the pots were irrigated three times a week.
A disease severity rating was done 3 week after planting according to Chen et al. (1987) as described in Table 1. Mean value of disease severity of ten seedlings in a pot was computed to represent one replication. The diseased seedlings and non-germinated seeds were washed in running tap water for 20 min. These seedlings and seeds were then treated with 0.5% NaOCl solution for 30 sec, rinsed with steril distilled water and plated on selective medium (Babai-Ahary et al., 2004) to reisolate the pathogen.
| Table 1: | Rating scale of disease severity |
 | |
The selective medium containing Cornmeal Agar (CMA) was amended with pimaricin (20 μg meq-1), penicillin (20 units mL-1), streptomycin sulfate (50 μg mL-1) and benomyl (50 μg mL-1). Then, the disease index was calculated for each treatment using the disease severity,
Y = (X-1)/3, |
| Where, | ||
| Y | = | Disease index, |
| X | = | Disease severity, |
| 3 | = | Total number of increments between one and four in the disease severity rating scale. At the end, ED50 for the isolates Pu8 and Pu2 was determined using SPSS software. |
Assessment of resistance in seedlings of sugarbeet lines: Seedlings of sugarbeet lines were evaluated for resistance to two P. ultimum isolates in a greenhouse of the Department of Plant Protection, Faculty of Agriculture, University of Tabriz, Tabriz. According to the quantities of ED50 obtained in the preliminary experiments the soil used for this experiment was mixed equally with inoculum. To be certain about a suitable concentration of inoculum in the soil, the mixtures were determined by soil drop technique. One-half of the plastic pots (10 cm in diameter and 12 cm in height) were filled with sterile soil mixture and the other half (upper half) with the soil containing inoculum. Seeds of sugarbeet lines were surface-disinfected using 0.5% sodium hypocholorid solution for 1 min. Ten seeds were then sown separately in 2 cm depth and the pots were maintained in the greenhouse at 22±2°C, 70±5 RH and 12 h light/dark for three weeks. The pots were watered and evaluated three times a week. Elit7233 was used as the susceptible control. Seeds of Elit7233 were also sown in steril soil and percentages of their germination were noted. The experiment was conducted using a factorial randomized block design with four replicates.
An evaluation approach to assess the resistance in sugarbeet lines was achieved using the modified 0-5 rating scale as described by Luterbacher et al. (2000) and shown in Table 2.
| Table 2: | Zero to five rating scale as modified from that of Luterbacher et al. (2000) |
 | |
An inoculated pot and a non-inoculated one, as the control, each containing 10 seeds were examined for each line. To eliminate the effect of growth failure, due to non-germination, in the control pots with that of treated pots, the 0-5 rating scale was applied to all the seedlings and the average rate obtained from control pots were deducted from the average rate of the treated pots to assign a disease rating for each line. Likewise, in polygerm lines, an average rate was calculated for all the seedlings emerged from a single seed.
RESULTS
Cfu determination of inoculum: Cfu developed in 1:10, 1:50, 1:100 dilutions series were too numerous to count. According to Stanghellini and Hancock (1970). The number of the emerged hyphae from the soil suspension drops should not be more than 4. The number of hyphae were observed in 1:1000 dilution was accordingly and it was set as the basis for cfu determination. After repeating the experiment for three times, cfu was shown to be 2.3x105 for Pu2 and 1.4x105 for Pu8.
ED50 determination of isolates: The mean value of disease severity and disease index were calculated for the treatments and ED50 value of either isolates was determined on the basis of the disease index. Results from this experiment for either of Pu2 and Pu8 isolates were noted briefly in Table 3 and the relations of the disease index (probit) to inoculum density (logarithm) was shown in Fig. 1 and 2. As shown in Table 3, ED50 value of the Pu8 was lower than that of Pu2 and Pu8 was more aggressive than the Pu2.
Assessment of resistance in sugarbeet lines: The sugarbeet lines reacted similarly to both isolates of P. ultimum var. ultimum (Pu8 and Pu2). The results from the resistance evaluation in the sugarbeet lines against the isolates Pu8 and Pu2 are shown in Table 4. The lines 8150-Bulk, Rhizofort, MstC2, Mst231 and 9597-P.58 were shown to be more resistant that the other lines. It should be noted that the treatment of Rhizofort with hymexazol and iprodion and apparent resistance in this line might have been due to the influence of these fungicides especially hymexazole.
| Table 3: | ED50 value and other parameters as calculated for the isolates of Pythium ultimum |
 | |
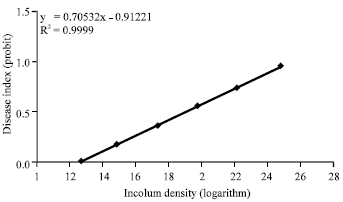 | |
| Fig. 1: | Relationship between inoculum density (logarithm) and disease severity (probit) in isolate Pu2 |
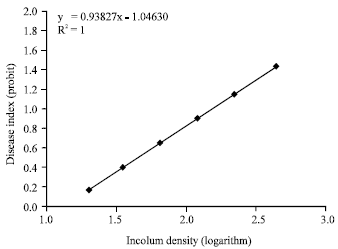 | |
| Fig. 2: | Relationship between inoculum density (logarithm) and disease severity (probit) in isolate Pu8 |
Results from assessing different levels of resistance revealed that resistance in the four resistant lines was as effective as the fungicide treatment in Rhizofort. Among the other lines, 7233-P.29, 19669T, C3.3 and Lit13 were more susceptible than the control (Table 4).
Grouping of the 20 sugarbeet lines and Elit7233 (control) was performed using ward’s cluster algorithm, based on mean scores of seedlings planted in the soils mixed with inoculum of the two isolates Pu2 and Pu8 (Fig. 3).
Three clusters obtained at cutting point with distance of 5. Results indicated that grouping pattern was in
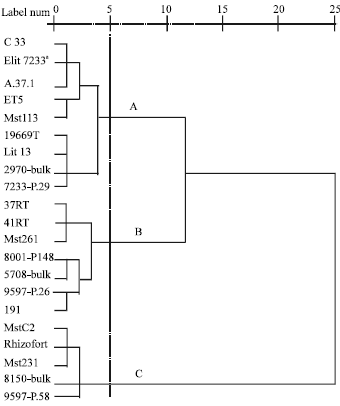 | |
| Fig. 3: | The obtained dendrogram from the clustering of 20 sugarbeet lines based on susceptibility; a: Control; A: Very susceptible; B: Susceptible; C: Resistant |
| Table 4: | Mean values of damping-off observed in seedlings of different sugarbeet lines grown in soil amended with inoculum of Pu2 and Pu8 isolates of P. ultimum |
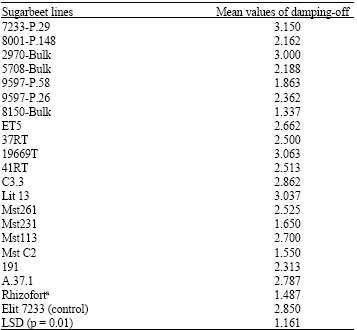 | |
| a: Pellets of seeds of this line contained hymexazol and iprodion | |
accordance with line susceptibility. Lines 8150-Bulk, Rhizofort, MstC2, Mst231 and 9597-P.58 were resistant and grouped at cluster C, whereas cluster A consisted of control (Elit7233) and the other 8 lines, which were very susceptible. The remaining lines were grouped as cluster B, which were susceptible. As noted earlier, apparent resistance in Rhizofort and its placement at cluster C might have been due to the influence of fungicides used in the seed treatment.
DISCUSSION
In the preliminary experiments, in order to produce enough hyphal swellings, isolates were grown in PDB for 2 days and mycelial mats were isolated. These mycelial mats were induced to produce hyphal swellings by three methods. In the two first methods, mycelial mats were washed with Sterilized Distilled Water (SDW). Then mycelial mats were incubated in petri plates containing SDW and/or 0.001M CaCl2 (Martin, 1992). In the third method, mycelial mats were rinsed in leaching buffer and were incubated in SDW at 20±1°C and darkness (Nelson and Craft, 1989). The third method gave the best results and we used this method in present study (Data not shown).
Luterbacher et al. (2000, 2005) have inoculated sterilized soil with P. ultimum (0.75% W/W), grown on cornmeal/sand medium, in order to screen Beta germplasm for resistance to damping-off. Instead, in this study, we have calculated and used ED50 values of either P. ultimum var. ultimum isolate. The ED50 values and virulence of the two selected isolates were different. This is in agreement with our previous studies in which Pu2 and Pu8 have been grouped as severe and very severe pathogens, respectively (Babai-Ahary et al., 2004). Present results emphasis on the necessity in using a quantified inoculum level in assessing resistance in plants against soilborne plant pathogens.
The evaluated sugarbeet lines had the same reactions to the both isolates of P. ultimum var. ultimum, but they had different levels of susceptibility and/or resistance. Comparison of mean values of damping-off (p = 0.01) of 20 sugarbeet lines and Elit7233 showed that the lines 8150-Bulk, Rhizofort, MstC2, Mst231 and 9597-P.58 were more resistant than other lines. The remaining lines were susceptible and the lines 7233-P.29, 19669T, C3.3 and Lit13 with higher mean values were more susceptible than Elit7233 (Table 4).
Cluster analysis of mean scores of 20 sugarbeet lines and Elit7233 (control) seedlings planted in the soils mixed with the inoculum of two P. ultimum var. ultimum isolates showed that the lines 8150-Bulk, Rhizofort, MstC2, Mst231 and 9597-P.58 were resistant, whereas the other lines were susceptible and very susceptible (Fig. 3).
The results from cluster analysis of mean scores of the sugarbeet lines (Fig. 3) were in agreement with the results from the comparison of mean values of damping-off (Table 4). The lines recognized as the susceptible hosts by comparison of mean values of damping-off were not significantly differed from the control (Elit7233) and as a result of cluster analysis they were precisely distinguishable from each other, appeared as susceptible and very susceptible lines and each formed a distinct cluster. Also, the lines 8150-Bulk, Rhizofort, MstC2, Mst231 and 9597-P.58 which were significantly differed from the control by the comparison of mean values of damping-off and that recognized as resistant lines, all formed one cluster. Apparent resistance of Rhizofort might have been due to the influence of Iprodion and especially hymexazole used in the pellets of the seeds. Hymexazol is one of the most effective fungicides that control seedling diseases caused by Aphanomyces cochlioides and Pythium sp. (Martin, 1992). These fungicides were effective in controlling damping-off, because the percent of emergence in Rhizofort did not differ significantly in infested and non-infested (steril) soils, also post-emergence damping-off was not observed in Rhizofort (data not shown). But, root and hypocotyl rot of the seedlings of this line were observed at the later stages so that even in some cases the rotting was so severe that some seedlings, in terms of the severity, were rated as number 3. Therefore, one may consider Rhizofort as a susceptible line to P. ultimum and ascribe a lower disease score for this line because of lack of incidence of pre and postemergence damping-off which, in turn, may be resulting from the fungicidal seed treatment. Root and hypocotyl rot at later growth stages were not detected in the resistant lines, therefore, the efficacy of resistance in these lines in controlling damping-off was more effective than the treatment by the fungicides.
Luterbacher et al. (2000, 2005) have similarly screened Beta germplasms for resistance to several important sugarbeet diseases, including beet mild yellowing virus (BMYV), Beet Yellows Virus (BYV), powdery mildew caused by Erysiphe betae and damping-off caused by P. ultimum and A. cochlioides. Their results show that, some of the germplasms tested for P. ultimum appear resistant with minimal symptom expression. In present study, we also observed resistance as the low severity of the disease.
ACKNOWLEDGMENTS
The authors are indebted to Dr. M.C. Luterbacher for kindly sending more information about rating scale. We also thank Dr. M. Moghaddam for his thoughtful comments on data analysis and the manuscript.
REFERENCES
- Babai-Ahary, A., M. Abrinnia and I. Majidi, 2004. Identification and pathogenicity of Pythium species causing damping-off in sugarbeet in Northwest of Iran. Aust. Plant Pathol., 33: 343-347.
CrossRefDirect Link - Chen, W., H.A.J. Hoitink and A.F. Schmitthenner, 1987. Factors affecting suppression of Pythium damping-off in container media amended with composts. Phytopathology, 77: 755-760.
Direct Link - Luterbacher, M.C., J.M. Smith, M.J.C. Ahser and L. Frese, 2000. Disease resistance in collections of Beta species. J. Sugarbeet Res., 37: 39-47.
Direct Link - Luterbacher, M.C., M.J.C. Asher, W. Beyer, G. Mandolino and O.E. Scholten et al., 2005. Sources of resistance to diseases of sugar beet in related Beta germplasm: 2. Soil-borne diseases. Euphytica, 141: 49-63.
Direct Link