
Andrey A. Aver`yanov
Research Institute of Phytopathology,
Russian Academy of Agricultural Sciences, B. Vyazemy, Moscow Region, 143050, Russia
Vera P. Lapikova
Research Institute of Phytopathology,
Russian Academy of Agricultural Sciences, B. Vyazemy, Moscow Region, 143050, Russia
Tatiana D. Pasechnik
Research Institute of Phytopathology,
Russian Academy of Agricultural Sciences, B. Vyazemy, Moscow Region, 143050, Russia
Vladimir V. Kuznetsov
Russian Peoples Friendship University, Faculty of Agriculture,
8 Miklukho-Maklaya str. Moscow, 117198, Russia
C. Jacyn Baker
Molecular Plant Pathology Lab., ARS USDA, Beltsville, Maryland 20705, USA
Plant Pathology Journal
Year: 2007 | Volume: 6 | Issue: 3 | Page No.: 242-247







ABSTRACT
Hydrogen peroxide mediates some anti-infectional responses of plants. It is toxic to pathogenous bacteria and fungi. To evaluate its minimal fungitoxic amounts, it was applied to spores of the causal agents of cucurbit scab (Cladosporium cucumerinum) and rice blast (Magnaporthe grisea, formerly Pyricularia oryzae) over the broad concentration range (10-2 to 10-14 M). Spore germination and appressorium formation were examined. The dose-response dependence was found multimodal for both fungi. The inhibition of spore germination was maximal at 10-2 M and practically disappeared at 10-5 - 10-6 M H2O2. However, weaker peroxide solutions had increased fungitoxicity. The inhibition of C. cucumerinum spore germination at 10-11- 10-12 M was almost the same extent as at 10-2 M H2O2. For M. grisea, two maxima of germination inhibition near 10-7 M and 10-10 M H2O2 were found. Here, the toxicity was higher than at its millimolar concentration. M. grisea appressoria formation repeated the aforementioned pattern generally. Therefore, hydrogen peroxide can have an antifungal action at very low concentrations. Such small amounts of H2O2 might be sufficient for plant defense responses as well.
PDF Abstract XML References
How to cite this article
Andrey A. Aver`yanov, Vera P. Lapikova, Tatiana D. Pasechnik, Vladimir V. Kuznetsov and C. Jacyn Baker, 2007. Suppression of Early Stages of Fungus Development by Hydrogen Peroxide at
Low Concentrations. Plant Pathology Journal, 6: 242-247.
DOI: 10.3923/ppj.2007.242.247
URL: https://scialert.net/abstract/?doi=ppj.2007.242.247
DOI: 10.3923/ppj.2007.242.247
URL: https://scialert.net/abstract/?doi=ppj.2007.242.247
INTRODUCTION
Hydrogen peroxide is a reactive oxygen species that presumably occurs as a metabolite in all aerobic organisms. Hydrogen peroxide is universally cytotoxic at high concentrations, mainly, due to conversion to the stronger oxidant, hydroxyl radical •OH (Apel and Hirt, 2004). Together with other reactive oxygen species, H2O2 is recognized to be involved in plant resistance mechanisms to infective diseases. These compounds suppress the development of microbes directly and participate in diverse defense responses (Baker and Orlandi, 1995; Garcia-Brugger et al., 2006).
For any biologically active compound, it is essential to know what concentrations are effective and what concentrations are actually present in vivo. To verify the role of endogenous H2O2, the latter is often applied exogenously. In many reports, concentrations of the added peroxide exceeded their natural level in plant tissues by several orders. Such high amounts are necessary because lower ones appear to be ineffective. For example, in tobacco cultured cells inoculated with incompatible bacteria Pseudomonas syringae pv syringae (Glazener et al., 1996) or zoospores of Phytophthora nicotianae (Able et al., 2000), the oxidative burst accompanies hypersensitive necrosis and yields micromolar level of H2O2 but only addition of millimolar amounts causes plant cell death. A similar discrepancy was found for tomato cells treated with the elicitor of Cladosporium fulvum (Lu and Higgins, 1999). This casts doubt on the role of the endogenous peroxide.
At first sight, tests of compounds taken in doses below their apparent activity threshold are meaningless. Meanwhile, the evergrowing body of evidence indicates that many substances are biologically active at very low levels such as 10-10-10-20 M. Importantly, the ranges of active concentrations may be divided by so called dead zones where the compound does not afford activity (Crain and Shen, 1995; Maltseva et al., 1998; Kátay and Tyihák, 2002; Ostrovskaya et al., 2003; Brailoiu1 et al., 2004; Burlakova et al., 2004). It is easy to imagine that an experimenter analyzing effects of a certain substance at more and more dilute solutions will believe the task is done when the first dead zone is achieved.
Earlier we studied the fungus Magnaporthe grisea (formerly Pyricularia oryzae), causing blast, the most harmful disease of rice. We reported that rice leaf diffusates suppressed the first two stages of the fungus development, namely, spore germination and appressorium formation. These fungitoxic effects were more pronounced in rice plants possessing inheritable or induced resistance to this disease than in susceptible plants. Hydrogen peroxide involvement in the antifungal action of diffusates was indicated by the inhibition of this action with exogenous catalase (EC 1.11.1.6) or low-molecular weight scavengers of H2O2 (Aver'yanov et al., 1993; Pasechnik et al., 1998; Aver'yanov et al., 2001). We estimated the hydrogen peroxide concentration in infection droplets on rice leaf surface as low as 5x10-7 M (Aver'yanov and Lapikova, 1988). Pure H2O2 at about 10-2 M inhibited M. grisea spore germination by 80% (Aver'yanov et al., 1987). Peng and Kuć (1992) reported that hydrogen peroxide ceased conidial germination of fungi Peronospora tabacina, Cladosporium cucumerinum and Colletotrichum lagenarium at about 2x10-5 M whereas 3x10-7 M had no effect.
This work examined effects of hydrogen peroxide on spore germination of M. grisea and Cladosporium cucumerinum over the wide range of H2O2 concentration (10-2 to 10-14 M). In addition, influence of the same amounts of peroxide on M. grisea appressorium formation was explored. The aim was to elucidate whether this substance can suppress or stimulate the fungus development at concentrations, natural in vivo but much lower than usually studied in this respect. The fungitoxicity of H2O2 was found multimodal, as the peroxide concentration decreased and actually it was significant at concentrations close to minimal.
MATERIALS AND METHODS
Fungi: The culture of the cucurbit scab fungus Cladosporium cucumerinum Ell. et Arth was obtained from the All-Russian Research Institute for Vegetable Breeding and Seed Production. The culture was initially sampled from cucumber plants. By means of colonies emerged from single spores, five fungal strains were isolated. The strain C5, aggressive against cucumber leaves of cv. Phoenix, was used. The cultures were maintained on agar-containing (20 mg mL-1) potato-glucose medium at +25°C. At the age of about 10 days, they were transferred to the refrigerator (about +6°C) for 10 days more to stimulate production of spores (conidia). The latter were harvested by washing with 50 mL distilled water per Petri dish and concentrated to 10 mL using the concentrator (Lapikova and Aver'yanov, 1992) with membrane filter (Nucleopore type PC MB, pores 8 μm, Costar). To wash spores the suspension was then diluted with fresh distilled water to 50 mL and was concentrated once more to 10 mL. Using a haemocytometer the spore count was diluted to 2x105 spore mL-1.
The natural isolate H5-3 (race 007) of the fungus Magnaporthe grisea (Hebert) Barr. was taken from the collection of the Research Institute of Phytopathology, Russia. About three months prior to the experiments, plants of the susceptible rice cv. Sha-tiao-tsao were infected with this strain. The fungus was re-isolated from leaf lesions. The culture was maintained on agarized carrot broth and spores were collected from 10 day colonies (Aver'yanov et al., 1993). They were washed and condensed to the concentration 3.5x105 spore mL-1 with the technique similar to that for C. cucumerinum.
Application of H2O2 and evaluation of its effects: Hydrogen peroxide (Suprapur, Merck, Darmstadt, Germany) was diluted to 10-2 M according to its extinction coefficient, 43.6 M-1 cm-1 at 240 nm (Claiborn, 1987). The spectrophotometer UV-260 (Shimadzu, Kyoto, Japan) was employed. From this stock, the set of standard solutions was prepared by 10-fold dilution of each previous sample. Mono-distilled water, passed through the purifying cartridge Elgacan C-114 (Elga Group, UK), was used for all procedures.
Spore suspensions of C. cucumerinum (50 μL) were mixed with 50 μL H2O2 in a well of a 96-well tissue culture plate Linbro (Flow Laboratories, UK). This gave the uniform final spore concentration 105 spore mL-1 and varied concentrations of peroxide. Each experimental treatment was represented in two equal wells. The material was incubated for 18 h in the dark at 23°C. Then it was fixed with ethanol (one drop per well) and observed under the inverted microscope (Leitz-Diavert, Wetzlar, Germany). In each treatment, the spore germination (%) was counted in 4 series of 100 spores each. The inhibition (%) of spore germination was calculated against the water control. In each treatment, the data of three independent experiments were pooled. Means and standard deviations were computed for n = 12.
Spore suspensions of M. grisea, in 10 μL drops, were placed into wells of a 96-well plate. Drops (10 μL) of water or water solution of catalase (10700 units mg-1 material, thymol-free, from bovine liver, Sigma, St. Louis, USA; EC 1.11.1.6) were placed in the same wells but separate from the suspension droplets. After 10 min, 80 μL of water or hydrogen peroxide was added to mix all components simultaneously. The final cell concentration was 3.5x104 spore mL-1 and catalase concentration was 50 μg mL-1. The final concentration of hydrogen peroxide is accounted for in the plots. The material was incubated for 5 h in the dark at 23°C and was fixed with ethanol. The inhibition of spore germination was calculated the same way as for C. cucumerinum. In two independent experiments, germination was counted in 4 and 5 series of 100 spores (n = 9) and all data were united for each treatment.
To estimate the effect of H2O2 on appressorium development, spore suspensions of M. grisea were treated as above but fixed after 9 h. Numbers of germinated spores and those carrying appressoria were counted. The inhibition (%) of appressorium formation by germinated spores against water control and the significance of this difference were evaluated for n = 9. This inhibition index shows the decrease in appressorium incidence, which was not due to decrease in spore germination.
All the experiments were carried out in the Research Institute of Phytopathology, Russia.
RESULTS
Effect of H2O2 concentrations on C. cucumerinum spore germination: In water, the germination of freshly harvested C. cucumerinum spores after 18 h varied around 53±15% in three experiments. Hydrogen peroxide suppressed germination strongly at 10-2 M. Upon dilution of peroxide, its effect decreased gradually until there was no significant difference from the water control at 10-6 M (Fig. 1). Interestingly, further dilution of H2O2 increased spore inhibition again, comparable to those of its highest concentrations. This index reached the maximum at 10-12 M H2O2 and then decreased to zero at 10-14 M.
To check whether the inhibition of fungus development was an irreversible arrest or just retardation, another experiment was carried out using longer (28 h) spore incubation in hydrogen peroxide (Table 1). In the water control, the extra time allowed more spore germination to occur. It was also observed in peroxide solutions at some but not highest concentrations.
Effect of H2O2 concentrations on M. grisea spore germination: To test whether the sensitivity of spores to low amounts of H2O2 is peculiar to species other than C. cucumerinum, we examined blast fungus M. grisea. Its spores developed faster than those of C. cucumerinum. In water, the germination of M. grisea spores was 77± 4% in 5 h. Expectedly, hydrogen peroxide was the most toxic at the maximum concentration 10-2 M (Fig. 2A). The effect decreased dramatically in weaker solutions and disappeared at 10-5 M. As with C. cucumerinum, far more diluted H2O2 also exhibited significant fungitoxicity to M. grisea, although to a lesser extent.
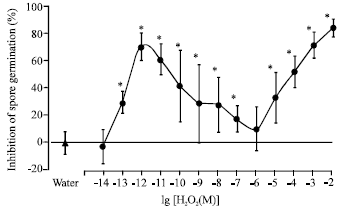 | |
| Fig. 1: | Effect of H2O2 on the germination of Cladosporium cucumerinum spores. Freshly harvested spores (2x105 spore mL-1) were incubated in peroxide for 18 h at 23°C. The plot represents the relative inhibition of spore germination in peroxide solutions against water control. Values are means±SD from three independent experiments. For every concentration, spore germination was counted in 12 series of 100 spores (n = 12). Asterisks indicate counterparts, where spore germination in H2O2 solutions differed from germination in water significantly at p<0.01 |
| Table 1: | Effect of H2O2 Concentration on Germination of Cladosporium cucumerinum spores |
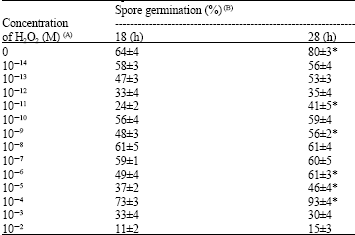 | |
| (A) Freshly harvested spores (2x105 spore mL-1) were incubated in peroxide for different times at 23°C. (B) Values are means±SD from one experiment. For every concentration, spore germination was counted in 4 series of 100 spores (n = 4). Asterisks indicate H2O2 concentrations where spore germination in 28 h differed from germination in 18 h significantly at p<0.05 | |
Two maxima were observed, at 10-7 M H2O2 and at 10-10-10-11 M H2O2. Again, there was no inhibition at 10-9 or 10-14 M H2O2. Exogenous catalase did not affect spore germination of M. grisea in water but abolished the fungitoxicity of peroxide over the entire concentration range.
To test the effect of H2O2 on appressorium formation, M. grisea spores were incubated for 9 h. The germination rate increased to 84±7% in water.
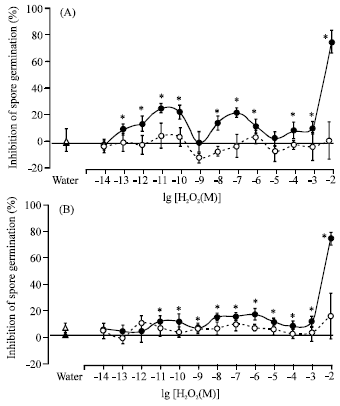 | |
| Fig. 2: | Effect of H2O2 on the germination of Magnaporthe grisea spores. Freshly harvested spores (3.5x105 spore mL-1) were incubated in peroxide at 23°C for 5 h (A) or 9 h (B). The plots represent the relative inhibition of spore germination in peroxide solutions against water control. Values are means±SD from two independent experiments. For every concentration, spore germination was counted in 9 series of 100 spores (n = 9). Asterisks indicate the same as in Fig. 1. Empty symbols represent treatments where media contained catalase (535 units mL-1) |
Hydrogen peroxide at 10-2 M reduced the germination rate in 5 and 9 h equally. However, at lower H2O2 concentrations, its inhibitory effect was reduced with the increased time of incubation. The bimodal dependency plot observed with the 5 h incubation looked less pronounced with the 9 h incubation treatments. Catalase remained protective under these conditions as well (Fig. 2B).
Effect of H2O2 concentrations on M. grisea appressorium formation: The frequency of appressoria formation was 12±4% of total spores and 13±5% of germinated spores after 9 h. The latter index represents the suppression of the fungus development in addition to that occurring with spore germination. As that index was diminished in H2O2 solutions (Fig. 3), the second phase of the development was peroxide-sensitive per se, not as a result of inhibition of the previous phase.
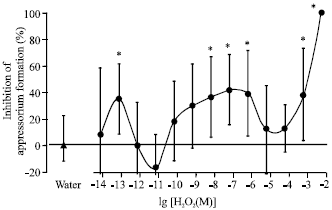 | |
| Fig. 3: | Effect of H2O2 on appressorium formation by Magnaporthe grisea spores. Freshly prepared spores whose germination is depicted on the Figure 2b were incubated in peroxide for 9 h at 23°C. The percentage of appressorium formation was calculated as a fraction of germinated spores. Asterisks indicate counterparts, where appressorium formation in peroxide solution differs from that in water significantly at p<0.05. The negative values mean stimulation towards water control but they were insignificant |
Although the percentage of appressoria varied more than that of germination it is seen that both processes were affected by peroxide concentration in a similar manner. The strongest peroxide concentration prevented the appressorium development completely and lost the effect at 10-4-10-5 M. Then the anti-appressorial activity returned at 10-6-10-8 M and 10-13 M H2O2, whereas other concentrations of the range tested were not effective.
DISCUSSION
As anticipated, hydrogen peroxide suppressed the spore germination of C. cucumerinum and M. grisea in a concentration-dependent manner. Its fungitoxicity decreased with its dilution and nearly disappeared at 10-5 -10-6 M. However, further dilutions of peroxide exhibited the toxicity again and in some cases even stronger than at millimolar levels. One mode of inhibitory activity was found for C. cucumerinum and two modes were found for M. grisea. The lowest concentration tested, 10-14 M, was ineffective at inhibiting spore germination of both fungi. Broadly speaking, it is possible that hydrogen peroxide may have some effects even under this level but we did not test this. In all treatments the added peroxide was the inhibitory factor of germination because catalase diminished or abolished the action. The second phase of the spore development for M. grisea, the appressorium formation, was also sensitive to hydrogen peroxide at concentrations as low as 10-13 M. The dose-response plot revealed dead zones similar to those found for spore germination and occupied approximately the same parts of the concentration axis. High variability of the appressorium frequency did not allow precise location of principal points of the concentration dependency.
We did not exclude possible stimulation of the fungus development by low doses of peroxide but found only inhibition. These effects may be universal among various fungi inasmuch as it was observed on two different fungal species. C. cucumerinum was somewhat more sensitive to peroxide although it is rather difficult to interpret the difference.
Therefore, the initial suggestion was supported that H2O2, at concentrations far beneath the apparent threshold of its biological activity, can affect the fungus development. The concentration/effect relations were found not simple but multimodal. Such a dependency including dead zones and peaks of activity at trace concentrations agrees with the concept of ultra-low doses (Burlakova et al., 2004). Although various substances in various biological and biochemical systems behave themselves in terms of this concept our results appear to be the first with H2O2 and fungal spores.
This work was not aimed at unraveling mechanisms of the effects reported. In general, dead zones may appear if a factor applied causes certain effect through multiple mechanisms, which are different in their sensitivity to the factor. Then some intensity of the factor (i.e., some concentration of a chemical) will be ineffective if it is too low for one and too high for other mechanism. Such relations are scarcely pertinent to gross structural damages but feasible in signaling. To our opinion, H2O2 damaged spores at maximal concentrations but was regulatory below 10-5-10-6 M.
In these experiments, high H2O2 amounts suppressed spores strongly regardless of the treatment time probably by irreversible oxidizing alterations. At some, not high peroxide concentrations, the inhibitory effect decreased with time and might be reversible to some extent. It is feasible that this slowing down of a parasite development caused by plant-originated hydrogen peroxide could contribute to the disease resistance as the host has more time to mobilize defense mechanisms.
In case of a multimodal dose-effect dependency, the term ED50 becomes inapplicable. Consequently, estimation of H2O2 concentrations underlying some biological effects by adding exogenous peroxide solutions also becomes meaningless because some one degree of the effect may be caused by several concentrations of peroxide which differ by several orders.
CONCLUSION
We think that not only direct fungitoxicity but also diverse anti-infectional plant responses, which are reversed by exogenous catalase, may be driven by low concentrations of endogenous hydrogen peroxide. In living tissues, these concentrations are more probable than, for example, millimolar ones. Our observations make it possible whereas the premise that only high amounts of H2O2 are needed to elicit defense responses suggests that endogenous amounts are inadequate for elicitation (Glazener et al., 1996; Lu and Higgins, 1999; Able et al., 2000).
It would be interesting to study effectiveness in other plant defense responses of H2O2 and other signal molecules (salicylic acid, for example) taken at low concentrations.
ACKNOWLEDGMENTS
The authors are very much grateful to Prof. E.B. Burlakova (Emanuel Institute of Biochemical Physics) for the valuable discussion of the work. We also thank Dr. A.N. Samokhvalov (All-Russian Research Institute for Vegetable Breeding and Seed Production) for the kindly provided culture of C. cucumerinum. This investigation was partially supported by the grant No 2682p sponsored by ARS USDA and mediated by the International Science and Technology Center.
REFERENCES
- Able, A.J., D.I. Guest and M.W. Sutherland, 2000. Hydrogen peroxide yields during the incompatible interaction of tobacco suspension cells inoculated with Phytophthora nicotianae. Plant Physiol., 124: 899-910.
PubMedDirect Link - Apel, K. and H. Hirt, 2004. Reactive oxygen species: Metabolism, oxidative stress and signal transduction. Annu. Rev. Plant Biol., 55: 373-399.
CrossRefPubMedDirect Link - Averyanov, A.A., T.D. Pasechnik, V.P. Lapikova and L.M. Gaivoronskaya, 2001. Fungitoxic responses of rice callus culture as an expression of inheritable resistance to blast. Implication of active oxygen. Plant Physiol. Biochem., 39: 415-424.
Direct Link - Baker, C.J. and E.W. Orlandi, 1995. Active oxygen in plant pathogenesis. Annu. Rev. Phytopathol., 33: 299-321.
CrossRefDirect Link - Brailoiu, E., J. Hoard, G., C. Brailoiu, M. Chi, R. Godbolde and N.J. Dun, 2004. Ultra low concentrations of morphine increase neurite outgrowth in cultured rat spinal cord and cerebral cortical neurons. Neurosci. Lett., 365: 10-13.
CrossRefDirect Link - Crain, S.M. and K.F. Shen, 1995. Ultra-low concentrations of naloxone selectively antagonize excitatory effects of morphine on sensory neurons, thereby increasing its antinociceptive potency and attenuating tolerance/dependance during chronic cotreatment. Proc. Natl. Acad. Sci. USA., 92: 10540-10544.
PubMedDirect Link - Garcia-Brugger, A., O. Lamotte, E. Vandelle, S. Bourque and D. Lecourieux et al., 2006. Early signaling events induced by elicitors of plant defenses. MPMI, 19: 711-724.
PubMedDirect Link - Glazener, J.A., E.W. Orlandi and C.J. Baker, 1996. The active oxygen response of cell suspensions to incompatible bacteria is not sufficient to cause hypersensitive cell death. Plant Physiol., 766: 759-763.
PubMedDirect Link - Lu, H. and V.J. Higgins, 1999. The effect of hydrogen peroxide on the viability of tomato cells and of the fungal pathogen Cladosporium fulvum. Physiol. Mol. Plant Pathol., 54: 131-143.
CrossRefDirect Link - Ostrovskaya, L.A., N.V. Blyukhterova, M.M. Fomina, V.A. Rykova, D.B. Korman and E.B. Burlakova, 2003. Inhibition of tumor growth with ultralow doses of doxorubicin under experimental conditions. Bull. Exp. Biol. Med., 135: 48-49.
CrossRefDirect Link - Pasechnik, T.D., A.A. Averyanov, V.P. Lapikova, E.D. Kovalenko and T.M. Kolomiets, 1998. The involvement of activated oxygen in the expression of the vertical and horizontal resistance of rice to blast disease. Russian J. Plant Physiol., 45: 371-378.
Direct Link