
Berna Bas
Department of Biology, Faculty of Science and Letter, University of Gaziantep,
27310 Sahinbey Gaziantep, Turkey
Namik Kemal Koc
Department of Plant Protection Agriculture, Faculty of Cukurova, University 01330-Balcall Adana, Turkey
Plant Pathology Journal
Year: 2006 | Volume: 5 | Issue: 1 | Page No.: 35-40







ABSTRACT
Embryogenic callus of Kütdiken lemon highly sensitive to Phoma tracheiphila was successfully used for selection via in vitro tolerance to diluted toxin of Phoma tracheiphila. Twenty four callus masses were obtained after six consecutive subcultures on the medium supplemented with toxin and then these callus clusters selected were dual cultured with Phoma tracheiphila, Penicillium italicum, P. digitatum, Phytophthora citrophthora, Colletotrichum gloeosporioides, Phomopsis citri, Diplodia natalensis and Geotrichum candidum. One stable resistant callus mass was selected and labeled as Kütdiken lemon 20b. The fungal growth of Phoma tracheiphila was strongly inhibited by culture liquid of Kütdiken lemon 20b cell suspension cultures in coculture experiment comparing to sensitive Kütdiken lemon and moderate resistant Zagara Bianca. In addition, the same boiled liquid has inhibited the fungal growth. By these results, rather than any PR proteins as inhibitors to fungal development, an alternative cause for defense reaction may be secondary metabolites which are toxic to the fungus.
PDF Abstract XML References
How to cite this article
Berna Bas and Namik Kemal Koc, 2006. In vitro Selection of Kütdiken Lemon 20b to Canditate for Resistance to Phoma tracheiphila. Plant Pathology Journal, 5: 35-40.
DOI: 10.3923/ppj.2006.35.40
URL: https://scialert.net/abstract/?doi=ppj.2006.35.40
DOI: 10.3923/ppj.2006.35.40
URL: https://scialert.net/abstract/?doi=ppj.2006.35.40
INTRODUCTION
One of the most serious problems encountered in lemon is mal secco disease caused by Phoma tracheiphila ((Petri) Kanc. Et Ghick.). This pathogen of lemon exists in all the Mediterranean countries except Spain. Citron (Citrus medica L.), bergamot (C. bergamiarisso), sour orange (C. aurantium L.) and lemon (C. lemon Burm) are highly sensitive to mal secco disease[1].
Applications of fungicides and cultural management to control the fungus have caused problems on environmental pollution and resistant fungal strains[2,3]. Therefore, breeding of varieties resistance to mal secco disease is of major importance. Although in the countries where the fungus has been widespread conventional breeding has been worked in order to obtain resistant varieties, no lemon cultivars improved for the disease resistance have been developed yet and high quality with horticulture value for lemon is demanded by market[4]. Because lemon is apomictic and vegetatively propagated even if exposed to mutagenic agents, possibilities to obtain resistant cultivars are very low.
Recently developments of in vitro selection techniques have provided novel facility for reliable selection of mutant variants in laboratories through exposure of plant cell and tissue to stress factors in vitro[5-7]. Cell culture is a valuable tool for screening the induced variants for direct or indirect utilization in breeding programme for crop improvement[8]. Recovery of disease-resistant plants by selecting cell cultures resistant to a toxin produced by a pathogen represents a direct application of cell culture approaches to crop improvement[9]. Induction of mutations for resistance to various pathogens at the cellular level and regeneration of plants may facilitate the selection of disease resistant plants[10]. Successfully selections have been reported in various plants[11-17].
The use of the tissue culture methods for citrus crop species have practical benefits[18]. Friable embryogenic calli of citrus is an important tool for breeding objective improvements. Classical breeding of citrus, which is perennial and has long juvenility, requires several years. Selection of plant cell cultures resistant to toxin of P. tracheiphila was studied by Nadel and Spiegel-Roy[10] and Gentile et al.,[19]. The present research describes the selection procedure for inducing and selecting resistant callus masses to toxins of P. tracheiphila from embryogenic Kütdiken lemon calli which is local variety in Turkey.
MATERIALS AND METHODS
Plant and fungal materials: Previously obtained embryogenic calli of Kütdiken (KT) lemon susceptible to mal secco were used in this work P. tracheiphila isolate was obtained from Alata Horticulture Research Institute (Içel-Turkey) by Dr. Ismail Çimen. Penicillium italicum, P. digitatum, Phytophthora citrophthora (Ph-30), Colletotrichum gloeosporioides (C-40), Phomopsis citri (P107), Diplodia natalensis and Geotrichum candidum (G-11) were kindly provided by Dr. Eldon Brown (Citrus Research and Education Center, Lake Alfred, Florida, USA).
Toxin production and calibration: Culture filtrate production was carried out as described previously[20]. Purification procedure was according to Nadel and Spiegel-Roy[10]. Filtrated fungal culture liquid was concentrated to 5 mL (1/80 v/v) at 45-50°C by rotary evaporation, precipitated with 3 vol. acetone at -18°C for overnight, lyophilized and then dissolved in the buffer Na-Phosphate pH 5.5. After centrifugation, supernatant including partially purified toxin was filter-sterilized through 0.22 μm membrane filter and stored at -30°C in deep freeze until use.
The effective dosage (ED50), which was expressed by the concentration of the toxin causing 50% disease of the population, was determined using multiple regression analyses[21].
Callus maintenance and establishment of cell suspension cultures: The calli previously established were subcultured every 5-6 weeks on MT[22] medium containing 5% sucrose, solidified with 1% agar (Difco) and maintained in culture room at 251°C under 16 h light (1000 lux) condition.
Cell suspension cultures were obtained by placing 100 mg of calli in a 100 mL flask containing 25 mL liquid basal MT medium on shaker at 125 rpm and subcultured at 2 week intervals.
Selection of callus: Partially purified toxin was incorporated to the autocloved basal MT medium at 1/40, 1/50 and 1/60 dilutions which are below and above ratios of effective dose (ED50 = 1/50) including ED. Twenty ml each of these conditioned media were dispensed to sterile Petri dishes (9 cm diameter). Fifty callus masses of KT 2.5 mg each were placed on the media containing diluted toxin. Each of toxin treatment contained 5 replicates. The control was basal MT medium. The calli were incubated in a culture room under the same conditions as described above by subculturing at 5-6 week intervals. After each subculture, growth index of calli was calculated by measuring fresh weights of the callus masses (first weight subtraction from last weight). Calli were subcultured 6 times on the media containing same concentrations of toxin for 9 months then transferred to without toxin media for 6 consequent subculturing. Preliminary evaluation of all callus lines was allowed to eliminate transient escaping callus which exhibited excess growth in the presence of mal secco toxin or to adopt retarded growth of callus during 9-month subculture.
Cell suspension cultures for selection of toxin-resistant cells: The cell suspension cultures were filtered through nylon mesh (500 μm pore size) and then plated on petri dishes (5 cm diameter) at density of 103 cells mL–1 in solid basal MT media supplemented with the toxin at 1/40, 1/50 and 1/60 dilution ratios and 0.4% final agar. Grown calli were subcultured 3 times on the media with same concentrations of toxin under the same conditions as described above.
Dual culture of fungi and calli and/or total protein extract: Experiment was done in two different manner:
| • | Dual cultures were carried out according the method of Storti et al.[23]. Two same callus pieces (500 mg each) originated from first selected ones were oppositely placed at the same distance from the center of 5 cm petri dishes containing solid basal MT medium. After incubation for 30 days, culture disks in 4 mm diameter of P. tracheiphila, Penicillium italicum, P. digitatum, Phytophthora citrophthora (Ph-30), Colletotrichum gloeosporiodes (G-40), Phomopsis citri (P107), Diplodia natalensis and Geotrichum candidum (G-11) were put on the center of each Petri dish. Five replicates were prepared for respective pathogens. They were incubated for additional 6-10 days in the dark under the same conditions as described above. Fungal growth was visually estimated. |
| • | After sterile PDA media (20 mL) at 45-50°C was incorporated with 5 μL of Penicillium italicum spore suspension (9x106 density at 700 nm and 0.75 absorbance) sterile discs of Watmann No:1 paper were placed at same distance in triangular position on 9 cm petri dishes containing same media. Each disc was dropped with 100 μL of total protein. Negative controls were consisted of sterile ddH2O and extraction buffer of protein. Results was evaluated depending on development of a fungal colony in density around paper discs in 3-5 days after incubation. |
Total protein was obtained from liquid media according to de Vries et al.[24] then one part of the extract was hold in boiling water for 100 sec and applied on various fungus.
Conditioned medium with cell suspension liquid: Each culture liquid of from first selected callus cell suspension cultures containing 3 different levels of toxin was aseptically obtained by centrifugation and then mixed with basal MT medium containing 1.8% agar in the ratio of 1:1 (v/v) in 9 cm petri dishes with 3 replicates for each culture liquid. Culture disks in 4 mm diameter of P. tracheiphila were placed on the center of the solidified media. Fungal growth was monitored after incubation for 10 days in the dark under the same conditions as described above. The control was solid basal MT medium without culture liquid. Only P. tracheiphila was used in this experiment.
Statistical analysis: Duncan’s Multiple Range Test was used to analyse the differences between means of the three toxin treatments and controls for callus and fungal growth of P. tracheiphila on the respective conditioned media.
RESULTS AND DISCUSSION
Inhibition of cell growth on the media containing three different concentrations of toxin was significant up to 5th subculture. Especially, the effect was greatest at 5th subculture, callus development being only 6-10% of that for the sensitive calli KT (Table 1). The cells from suspension cultures transferred to solid MT media exhibited greater sensitivity to the toxin than callus grown on solid MT media. Although two cell masses developed in 2 mm diameter were subcultured to solid MT media containing the same concentrations of toxin, cell growth was inhibited at the second subculture and no growth was recorded for 6 months. This could be due to inhibition of cell division by the toxin in the culture medium.
| Table 1: | Growth index of the callus on the medium modified with toxin and without toxin. Mean separation was performed with General Linear Models (GLM) procedure of SAS 6.12 |
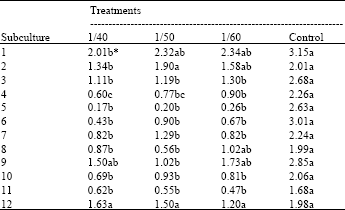 | |
| *Means followed by the same letter within lines are not significantly different at p = 0.05 | |
First obvious visual evidence of toxin resistance at callus level was obtained on the media containing 1/40 and 1/50 dilutions of toxin at the 5th subculture (Fig. 1). In spite of the toxin, a few callus colonies have grown continuously in white color like control colonies although growth of a greater number of colonies ceased and colored in brownish. Totally 50 survived callus masses were subjected to the last 6th subculture by transferring to the MT media containing the same diluted toxin (1/40 and 1/50). Twenty six callus masses out of 50 were eliminated, thus totally 24 calli successfully survived (3 colonies from 1/50, 21 colonies from 1/40). The survived callus masses were transferred to the basal MT medium without the toxin and further subcultured 6 times on the same medium. After 12th subculture, selected callus masses indicated almost normal growth as control (Table 1). These selected calli were cocultured with P. tracheiphila in order to test the continuous tolerance. One line of the calli derived from 1/40 dilution was labeled as Kütdiken 20b which strongly inhibited mycelial growth of P. tracheiphila (Fig. 1). While fungal invasion by Phytophthora citrophthora, Geotrichum candidum and Diplodia natalensis isolates in dual culture were strongly inhibited by selected callus Kütdiken 20b, inhibition was relatively weak against Colletotrichum gloeosporioides (C-40) and Phomopsis citri isolates by visual estimation. The inhibitory effect of selected Kütdiken 20b callus on Penicillium italicum was same as that on P. tracheiphila. These results indicate that the tolerance of Kütdiken 20b callus acquired by exposure to P. tracheiphila toxin is non specific to the pathogens tested here and this tolerance remained even after the subculture without exposure to the toxin.
Culture liquid from cell suspension cultures of Kütdiken 20b callus inhibited fungal growth of P. tracheiphila as shown in Fig. 2. Diameter of fungal colony of P. tracheiphila grown on the conditioned medium with culture liquid of Kütdiken 20b was the smallest in comparison to the media of sensitive Kütdiken and moderately resistant Zagara Bianca (Table 2).
| Table 2: | Different fungal growth of P. tracheiphila on the solid MT media containing culture liquid from 30 day old cell suspension cultures of sensitive Kütdiken lemon, tolerant Kütdiken lemon 20b and moderately tolerant Zagara Bianca |
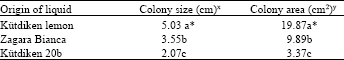 | |
| xValues are means of three replicates and obtained by measurement of colony size at three different positions in petri dishes. yValues are means of three replicates and obtained by calculation of covered area *Means within a column followed by the same letters are not significantly different at p = 0.05 | |
 | |
| Fig. 1: | Growth of callus under different conditions. a) Development of callus lines on the selection medium containing 1/50 diluted toxin in 20 days(top at left) and 40 days(top at right), respectively, after 6th subculture. b) Inhibition zones obtained by dual culture of selected lemon calli and P. tracheiphila. Selected "Kutdiken 20b" callus on 1/40 diluted toxin medium, selected calli on 1/50 diluted toxin medium, and control Kütdiken on basal MT medium were cultured for 30 days, culture disk of P. tracheiphila in 4 mm diameter was inoculated, and then incubated for additional 1 week |
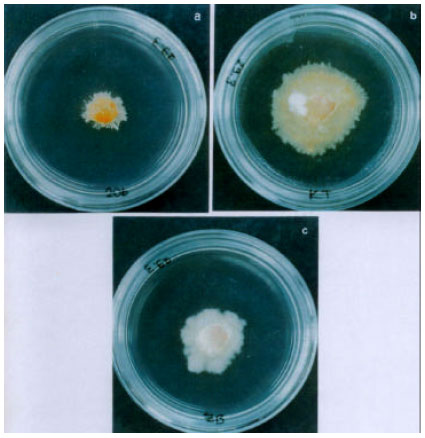 | |
| Fig. 2: | Inhibition of fungal growth of P. tracheiphila by culture liquid from cell suspension cultures exposed to diluted toxin of P. tracheiphila . Culture liquid from "Kutdiken 20b" (a), Zagara Bianca (b), and Kutdiken cell suspension cultures (c) was mixed with basal MT medium before solidifying with agar. The media were inoculated with culture disks of P. tracheiphila in 4 mm diameter, and then incubated for 1 week |
When the boiled culture liquid of Kütdiken 20b was mixed with MT medium and used for inhibition test against P. tracheiphila and P. italicum, the inhibitory effect still remained. Gentile et al.[25] have suggested that there is a positive correlation between secretion of Pathogenesis Related (PR) proteins such as chitinase and β-1,3-glucanase by tolerant cells and resistance to P. tracheiphila by these enzymes. In the case of our study, however, we concluded that if any PR proteins as secreted substances in the conditioned liquid media of Kütdiken 20b were contained, fungal growth would not be prevented because these proteins were inactivated by boiling. The results obtained from boiled and non-boiled conditioned liquid medium of Kütdiken 20b confirm the assumption that PR proteins do not play role in tolerance to P. tracheiphila for Kütdiken 20b. We consider that not PR proteins but other organic substances may be secreted into liquid culture media, leading to fungal inhibition. Egorava [26] has reported that resistant trees to mal secco contain high levels of alkaloids. Plant cell cultures produce a variety of secondary metabolites such as terpenoids, alkaloids and phenols. Some of these compounds occur constitutively and are thought to function as preformed inhibitors associated with non-host resistance[27-30]. In this preliminary experiment, high performance liquid chromatography analysis of organic compounds from cultured cells and conditioned media of Kütdiken 20b and sensitive Kütdiken brought an idea in which may be possible for Kütdiken 20b to secrete some secondary metabolites to protect against the toxin by these metabolites. However, it is needed to obtain more data on biochemical and physiological studies of the toxin-defense compounds.
ACKNOWLEDGMENTS
This project was supported by Research Foundation of Çukurova University (Project No: FBE 95-26). We are grateful to Scientific and Technical Research Council of Turkey (TÜBITAK-ANKARA) that supported with scholarship (NATO-A2) for Berna Bas and Citrus Research and Education Center (CREC Lake Alfred-Florida) that supported partly the project as a part of doctorate dissertation.
REFERENCES
- Rines, H. and H.H. Luke, 1985. Selection and regeneration of toxin-insensitive plants from tissue cultures of oats (Avena sativa) susceptible to Helminthosporium victoriae. Theor. Applied Genet., 71: 16-21.
CrossRefDirect Link - Behnke, M., 1979. Selection of potato callus for resistance to culture filtrates of Phytophthora infestans and regeneration of resistant plants. Theor. Applied Genet., 55: 69-71.
CrossRef - Gentile, A., E. Tribulato, Z.N. Deng, E. Galun, R. Fluhr and A. Vardi, 1993. Nucellar callus of 'Femminello' lemon, selected for tolerance to Phoma tracheiphila toxin, shows enhanced release of chitinase and glucanase into the culture medium. Theor. Applied Genet., 86: 527-532.
CrossRefDirect Link