
Kan-Fa Chang
Alberta Agriculture, Food and Rural Development, Field Crop Development Centre, Lacombe, AB T4L 1Wl,
Sheau-Fang Hwang
Alberta Research Council, Vegreville, AB T9C 1T4,
Heping Wang
Alberta Research Council, Vegreville, AB T9C 1T4,
George Turnbull
Alberta Research Council, Vegreville, AB T9C 1T4,
Ronald Howard
Alberta Agriculture, Food and Rural Development, Crop Diversification
Centre South, Brooks, AB Tl R 1E6, Canada
Plant Pathology Journal
Year: 2006 | Volume: 5 | Issue: 1 | Page No.: 15-19







ABSTRACT
The present research was conducted to characterize possible biocontrol agents and to determine specific etiological factors affecting severity of infection by S. sclerotiorum. Greenhouse studies showed that incubation period, inoculation technique and temperature had significant effects on sclerotinia blight of coneflower. Longer periods of incubation after inoculation usually induced more infection in both Echinacea pallida and E. purpurea, although the former was more susceptible. For E. pallida, E. purpurea and E. angustifolia, seedling wilt and root rot symptoms caused by S. sclerotiorum were most severe when inoculum was placed 1 cm from seedling roots and diminished as the distance increased. Seedling wilt and root rot were more severe in plants grown at 5-15°C than in those grown at 10-25°C. A total of 74 strains of Trichoderma sp. were evaluated for antagonism against S. sclerotiorum in vitro. Strong or very strong antagonism was identified in 36 strains. This antagonism was confirmed for selected strains in greenhouse evaluations. Under field conditions, the efficacy of the Trichoderma treatment surpassed that of the fungicide fludioxonil and a Bacillus sp. biocontrol agent. These results suggest that certain Trichoderma strains have an excellent potential for use in managing sclerotinia blight of coneflower.
PDF Abstract XML References
How to cite this article
Kan-Fa Chang, Sheau-Fang Hwang, Heping Wang, George Turnbull and Ronald Howard, 2006. Etiology and Biological Control of Sclerotinia Blight of Coneflower Using Trichoderma Species. Plant Pathology Journal, 5: 15-19.
DOI: 10.3923/ppj.2006.15.19
URL: https://scialert.net/abstract/?doi=ppj.2006.15.19
DOI: 10.3923/ppj.2006.15.19
URL: https://scialert.net/abstract/?doi=ppj.2006.15.19
INTRODUCTION
The coneflower species Echinacea pallida Nutt. (pale-purple coneflower), E. angustifolia DC. (narrow-leaved coneflower) and E. purpurea (L.) Moench. (purple coneflower) are widely used for medicinal purposes and have the potential to become viable commercial crops in Alberta, Canada. Sclerotinia blight (Sclerotinia sclerotiorum (Lib.) de Bary) was first reported on coneflower in Alberta by Chang et al.[1] and is one of the most widespread and destructive diseases occurring on this crop. Sclerotinia usually produces dark brown to black stem lesions near the soil level and causes leaves to wilt, then die and roots to become rotted. Severely diseased plants show a droop symptom in which all leaves wilt completely and characteristic brown lesions and white mycelia appear at the base of the stems. Over 40% of coneflower fields surveyed in Alberta in 1997 showed infection by this omnivorous, soilborne plant pathogenic fungus[2].
Virulence of S. sclerotiorum is affected by environmental conditions. Air temperature, relative humidity and water potential have significant effects on the survival and growth of S. sclerotiorum[3,5]. Huang and Hoes[6] found that disease incidence increased when the distance between seed and sclerotia was reduced. Wang et al.[7] have reported successful fungicidal control of sclerotinia blight on coneflower.
The present investigation was concerned with the effect of type and position of S. sclerotiorum inoculum on disease initiation, as well as the effect of incubation period and air temperature on disease development on three species of coneflower. Currently, there are no fungicides registered for controlling sclerotinia blight of coneflower in Canada; therefore, biological control has the potential to become a vital component of disease management practices. In this study, Trichoderma isolates were screened for antagonism to S. sclerotiorum and the efficacy of antagonistic Trichoderma sp. were evaluated under both greenhouse and field conditions.
MATERIALS AND METHODS
Culture and inoculum: An isolate of S. sclerotiorum, originally obtained from E. angustifolia in Brooks, Alberta[1], was used in this study. Two types of inocula were prepared: (1) five-mm mycelial disks of S. sclerotiorum were removed from 2-3 day-old cultures, which were actively growing on Potato-Dextrose Agar (PDA), using cork borers and (2) plastic bags filled with sterile oat grains were inoculated with three to four mycelial disks. Inoculated bags were incubated at room temperature (20-22°C) for three weeks and shaken periodically to ensure complete colonization of the grain. The inoculum was not dried before application.
Effect of incubation period on S. sclerotiorum infection: E. pallida and E. purpurea were grown in fibre pots (12 cm diameter) under greenhouse conditions. Two mycelial disks (5 mm diameter) were buried 0.5 cm below the soil surface next to the roots of the seedlings (2-3 leaf stages) in each pot. Inoculated pots were sealed in black plastic bags to maintain constant high temperature (15-28°C) and relative humidity (32-98%) conditions and were incubated for 24, 48 and 72 h. The maximum/minimum temperature and relative humidity were recorded by a digital thermo-hygrometer (Friendswood, TX, USA). Ten pots were used for each of three replications for each incubation period. Seedling wilt (percent) was recorded at 1, 5 and 10 days after inoculation. Root rot severity was measured two weeks after inoculation according to the following scale: 0 = healthy roots; 1 = lesions on lower stems or roots; 2 = partially dead seedling still attached to the roots; 3 = dead seedlings completely separated from the roots and 4 = severely rotted roots. The experiment was repeated once.
Effect of temperature and inoculation distance on S. sclerotiorum infection: Effect of temperature on host infection by S. sclerotiorum was assessed on E. angustifolia using two controlled environmental chambers set on a 12/12 h day/night cycle, one at 5/15°C and the other at 10/25°C. A 5-mm disk of mycelial inoculum was placed 1 and 2 cm away from the seedling roots for each temperature setting. Each treatment consisted of 10 pots, 10 seedlings in each, replicated three times in a two-factor, completely randomized design. Percent seedling wilt and root rot severity, using the same root rot severity scales as described above, were recorded two weeks after inoculation. The experiment was repeated once.
Concentration effect of S. sclerotiorum on disease development: S. sclerotiorum was propagated in sterilized oat grains, then air-dry and ground into powder. The resulting inoculum was incorporated into soilless mix at ratios of 1:5,000, 1:1,000, 1:500, 1:100, 1:50 and 1:25 (w:w). Four-week-old seedlings of E. angustifolia were transplanted into these inoculated substrates (ten pots for each concentration) and incubated in a growth chamber at 22°C. Plant mortality was recorded for each concentration after 6, 12, 18 and 24 days of incubation.
In vitro assessment of Trichoderma sp. for antagonism to S. sclerotiorum: Mycelial disks were cut from the margins of 74 growing colonies of Trichoderma sp. isolates collected from fields at Arrowwood, Bow Island and Carmangay in Alberta, Canada and in Tianjin, China. These disks were transferred to the outer edge of PDA plates and mycelial discs of S. sclerotiorum were transferred to the opposite side of each plate. The plates were incubated at 20°C. Growth of Trichoderma sp. colonies over the S. sclerotiorum colonies was measured after eight days of growth in paired culture. The trial was repeated using six isolates showing strong antagonism to the pathogen.
Suppression of S. sclerotiorum using Trichoderma strains
Growth chamber study: Ten isolates of Trichoderma sp. and one isolate of S. sclerotiorum were propagated on sterilized oat grains. The isolates were air-dried separately and ground into powder. Each powdered isolate of Trichoderma sp. was mixed with an equal amount of S. sclerotiorum powder, then incorporated into soilless mix at the six rates described above. Noninoculated soilless mix and soilless mix inoculated with S. sclerotiorum served as controls. E. angustifolia seedlings were transplanted into these substrates at the two-leaf stage and incubated in a growth chamber at 22°C. Plant survival was recorded for each treatment after two and four weeks of incubation.
Field study: Trials comparing the inhibitory effect of two biocontrol agents and a commercial fungicide against S. sclerotiorum were initiated at two sites, the McLeod and Lendrum Farms, near Brooks, Alberta. Treatments included the fungicide fludioxonil (Maxim XL), an isolate of Trichoderma sp., an isolate of Bacillus sp. and an inoculated control. The pathogen, the fungicide and the biocontrol agents (Trichoderma sp. and Bacillus sp.) were incorporated into the soil while the seedlings were being transplanted.
Data analysis: Seedling wilt and root rot severity from all experiments were analyzed by analysis of variance, Least Significant Difference (LSD) mean separation and Bartlett’s test for homogeneity of variance with the Statistical Analysis System 8.1 (SAS Institute, Cary, NC).
RESULTS AND DISCUSSION
Incubation periods: The results of two experiments for both E. pallida and E. purpurea were pooled because heterogeneity of variance was not validated by Bartlett’s test (Table 1). Incubation periods affected S. sclerotiorum infection in both coneflower species.
| Table 1: | Effect of length of incubation period after inoculation with Sclerotinia sclerotiorum on seedling wilt and root rot severity of Echinacea pallida and E. purpurea in greenhouse experiments |
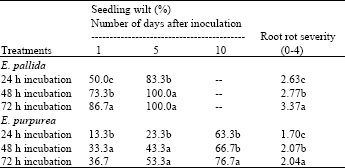 | |
| Values are the means of three replications in each of ten pots inoculated with two 5-mm diameter mycelial disks buried next to seedling roots. Means in a column followed by a common letter(s) are not significantly different (P≤0.05) according to least significant difference test | |
| Table 2: | Effect of temperature and inoculum distances from seedling roots on seedling wilt and root rot severity of Echinacea angustifolia seedlings inoculated with either infested oat grains or mycelial disks of Sclerotinia sclerotiorum |
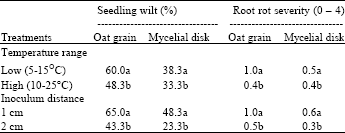 | |
| Values are the means of three replications in each of ten pots inoculated with one 5 mm diameter mycelial disk or 3-5 infested oat grains buried at designated distances from the seedling roots. The experiment was conducted in controlled environmental chambers. Means in a column followed by a common letter(s) are not significantly different (P≤0.05) according to least significant difference test | |
| Table 3: | Effect of concentration of Sclerotinia sclerotiorum inoculum placed in the growth substrate on survival of E. angustifolia seedlings in greenhouse evaluations |
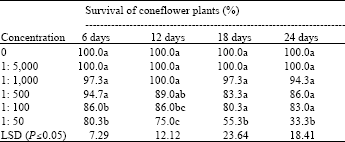 | |
| Values are the means of ten replicates. Means in a column followed by the same letter(s) are not significantly different according to least significant difference test at P≤0.05 | |
Longer periods of incubation usually induced more disease. For E. pallida, seedling wilt caused by Sclerotinia infection was 50% after 24 h incubation and reached 86.7% after 72 h. Echinacea purpurea was considerably less susceptible to Sclerotinia infection; seedling wilt was 37-50% lower after 24 h incubation. All E. pallida seedlings were severely infected or dead 10 days after inoculation, while 23-37% of E. purpurea seedlings remained healthy.
| Table 4: | Evaluation of 74 Trichoderma sp. isolates for aggressiveness to Sclerotinia sclerotiorum on potato dextrose agar plates |
 | |
| Weak: Slight overgrowth of Trichoderma onto S. sclerotiorum mycelia. Moderate (A): Overgrowth of ≤ 25% of the area covered by S. sclerotiorum mycelia. Strong (A+): Overgrowth of 25-50% of the area covered by S. sclerotiorum mycelia. Very strong (A++): Overgrowth of ≥50% of the area covered by S. sclerotiorum mycelia | |
| Table 5: | Effect of ten isolates of Trichoderma spp. on survival of coneflower seedlings inoculated with Sclerotinia sclerotiorum in greenhouse evaluations |
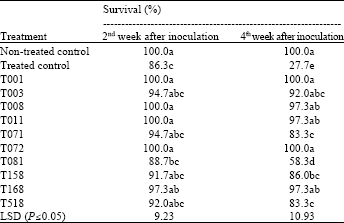 | |
| Values are the means of ten replicates. Means in a column followed by the same letter(s) are not significantly different according to least significant difference test at P≤0.05 | |
| Table 6: | Field assessment of biological control of Sclerotinia sclerotiorum at the McLeod and Lendrum Farms in Brooks, AB in 2001 |
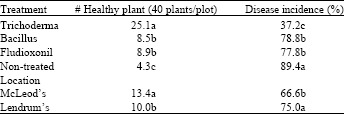 | |
| Data were combined across treatments or locations to show the effect of treatment or location on number of healthy plant and disease incidence since there was no statistical interaction (P = 0.05) between location and treatment. Means in a column followed by a common letter(s) are not significantly different (P≤0.05) according to least significant difference test. | |
Results from the incubation period trials indicated that high relative humidity favored rapid disease initiation and development. Abawi and Grogan[3] reported similar observations in naturally infested soils and stated that moisture was a limiting factor in the development of white mold on beans.
Temperature and inoculation distance: There were no significant interactions between temperature and inoculation distance. The results of two identical trials were combined according to Bartlett’s homogeneity test. Lower temperatures (5-15°C) resulted in more Sclerotinia infection than did higher temperatures (10-25°C) with both oat grain and mycelial disk inocula (Table 2). This suggests that the lower temperatures are more optimal for infection by S. sclerotiorum. This is consistent with results from other researchers, since carpogenic germination of S. sclerotiorum is favored by lower temperatures (10-15°C). Imolehin et al.[8] also observed that maximal radial growth and sclerotial germination for S. minor occurred at 18°C and that higher temperatures were less favorable for both radical growth and sclerotial germination. Seedling wilt was 12% higher at 5-15°C than at 10-25°C and disease severity was 0.6 points higher where oat grain inoculum was applied. Seedling wilt and root rot were 5% and 0.1 severity units higher, respectively, at 5-15°C than at 10-25°C where the mycelial disk inoculum was used. The effect of inoculation distance under greenhouse conditions was the same as that observed in controlled environmental chambers. The closer the inoculum was placed to seedlings, the higher the wilt incidence and root rot severity.
Colonized oat grain was more effective than mycelial disks for artificial inoculation, although mycelial disks could also cause substantial infection. Huang and Kozub[9] confirmed that mycelia could survive the severe winter conditions in Alberta and so may be an important source of field infection.
Inoculum concentration: Plant mortality increased with the concentration of S. sclerotiorum and with incubation time (Table 3). The percent survival of E. angustifolia plants was significantly (P≤0.05) lower when they were infected with S. sclerotiorum at the concentration of 1:50 than at almost all other concentrations for each incubation period. Longer incubation periods raised the infection level, especially under higher inoculum concentrations. For instance, plant survival declined by 47% from day 6 to day 18 at the concentration of 1:50, but declined by only 6-11 % at the inoculum concentration range of 1:100-500 and remained constant at concentrations of 1:1000 and 1:5000.
Isolation of Trichoderma sp. antagonistic to S. sclerotiorum: Thirty six of the 74 isolates showed strong to very strong antagonism to the pathogen in vitro, while 26 isolates showed no antagonism (Table 4). On the PDA plates, very strongly aggressive isolates overgrew more than 50% the area covered by S. sclerotiorum. Among the six isolates of Trichoderma sp. in the second trial (data not presented), isolate TJ-1 showed the most aggressiveness and completely overgrew the S. sclerotiorum colonies. At a later stage, Trichoderma sp. mycelia colonized the sclerotia and produced abundant conidia on them.
Biological control of S. sclerotiorum on E. angustifolia: In the growth chamber study, most of the E. angustifolia plants survived the first two-week incubation period, but the treatment effects only became apparent two weeks later (Table 5). Trichoderma strains T008, T011, T072 and T168 significantly increased the plant survival two weeks after inoculation, while all tested Trichoderma isolates had a significant effect on plant survival after four weeks of incubation. Plant mortality was significantly higher in the inoculated control treatment than in all substrates amended with Trichoderma sp. Isolates T001 and T072 completely controlled the disease and isolates T008, T011 and T168 showed significantly (P≤0.05) better control of the disease than the remainder of the isolates.
The field study produced comparable results to those of the greenhouse assessment. The results from two field locations (two farms) were combined because no statistical interactions were found between treatment and location (Table 6). All three treatments effectively increased the number of healthy plants and reduced the disease incidence. However, the Trichoderma treatment outperformed the Bacillus and fludioxonil fungicide treatments, with 25 healthy plants compared to 9 plants and 37% disease incidence compared to 78-79% in the plots treated with Bacillus and fludioxonil, respectively. Disease occurrence was less widespread at the McLeod farm than at the Lendrum farm.
This study addressed a need for information on Sclerotinia disease management using non-chemical means. Most growers and marketers require that echinacea products be organically grown and a biological approach to disease management satisfies these requirements. As our data suggest, many Trichoderma isolates showed strong antagonism to S. sclerotiorum in vitro and significantly reduced mortality of coneflower under both controlled environment and field conditions. Since coneflower is a valued specialty crop in Canada, biological control of sclerotinia blight may be economically feasible under conditions of moderate to high disease pressure.
REFERENCES
- Abawi, G.S. and R.G. Grogan, 1975. Source of primary inoculum and effects of temperature and moisture on infection of beans by Whetzelinia sclerotiorum. Phytopatholoy, 65: 300-309.
Direct Link - Huang, H.C. and J.A. Hoes, 1980. Importance of plant spacing and sclerotial position to development of sclerotinia wilt of sunflower. Plant Dis., 64: 81-84.
Direct Link