
M. Salehpour
Department of Plant Protection, Abourayhan Campus, University of Tehran, P.O. Box 11365/4117, Tehran, Pakdasht, Iran
H.R. Etebarian
Department of Plant Protection, Abourayhan Campus, University of Tehran, P.O. Box 11365/4117, Tehran, Pakdasht, Iran
A. Roustaei
Department of Plant Protection, Abourayhan Campus, University of Tehran, P.O. Box 11365/4117, Tehran, Pakdasht, Iran
G. Khodakaramian
Department of Plant Protection, Abourayhan Campus, University of Tehran, P.O. Box 11365/4117, Tehran, Pakdasht, Iran
H. Aminian
Department of Plant Protection, Abourayhan Campus, University of Tehran, P.O. Box 11365/4117, Tehran, Pakdasht, Iran
Plant Pathology Journal
Year: 2005 | Volume: 4 | Issue: 1 | Page No.: 85-90







ABSTRACT
Trichoderma viride T112 and T. viride (MO), T. harzianum (M) and T. harzianum T194 were used as potential biological agent for control of common root rot caused by Bipolaris sorokiniana. Cell free and antifungal metabolites produced by all Trichoderma isolates inhibited growth of Bipolaris sorokiniana. The inhibition varied among isolates of Trichoderma and ranged from 58.20 to 93.93% using the cellophane overlay method and from 66.66 to 98.25% in volatile test. Mycelial growth of B. sorokiniana was numerically reduced more by T. viride T112 than the other isolate tested (p<0.01). The seed soaking treatment and also soil treatment with T. vride T112 and T. viride (MO) were the most effective in reducing infection by the pathogen compared with the corresponding control (p<0.01) All isolates of Trichoderma increased plant height, fresh and dry weight of roots and shoots of wheat seedling compared with the uninoculated control. Among Trichoderma isolates, T. viride T112 and T. viride (MO) proved to be the best in supporting the growth of wheat (p<0.01).
PDF Abstract XML References
How to cite this article
M. Salehpour, H.R. Etebarian, A. Roustaei, G. Khodakaramian and H. Aminian, 2005. Biological Control of Common Root Rot of Wheat (Bipolaris sorokiniana)
by Trichoderma Isolates. Plant Pathology Journal, 4: 85-90.
DOI: 10.3923/ppj.2005.85.90
URL: https://scialert.net/abstract/?doi=ppj.2005.85.90
DOI: 10.3923/ppj.2005.85.90
URL: https://scialert.net/abstract/?doi=ppj.2005.85.90
INTRODUCTION
Root and foot rots are major diseases of wheat when moisture is deficient. The causal fungi are widely distributed as unspecialized pathogen on most small-grain cereal and numerous grasses. One of the important soilborne disease of wheat is common root rot caused by Bipolaris sorokiniana (Sacc. in Sorok.) Shoem. The most diagnostic symptoms caused by this pathogen is a dark brown or blackened subcrown internode. The infection and diagnostic discoloration may extend into the crown and a short distance up the culms. Diseased plants occur randomly or in irregular patches and appear stunted and chlorotic. Browning or blackening of the primary or secondary root system may be observed on close examination of washed roots. Seedling infection is reduced by shallow seedling and using clean or chemically disinfected seed. Also late autumn seedling of winter wheat is recommended to decrease seedling exposure to warm soil temperature. Reduced seedling infection, however is no guarantee against infection at the later growth stages Soil fertility must be adequate and balanced to support vigorous root and shoot growth. However, excessive fertilization, especially with nitrogen favors the disease by promoting vegetative growth, which in turn increases transportation and accelerates plant water stress. A few cultivars in Canada are resistant to B. sorokiniana[1], also B. sorokiniana on leaves can be controlled with conventional fungicides[2]. Control is difficult and expensive because the pathogen infects leaves, crown, rhizomes and roots of susceptible species and may be active on one or more plants of host plants throughout the growing season[3].
Biological agents could be an important components in control of B. sorokiniana if effective and reliable formulation were readily available and could be integrated with chemical fungicides. The antagonistic activities of Trichoderma and Gliocladium species against plant pathogens have been studied, extensively[4-10]. Some bacteria and fungi have been evaluated for antagonistic activity on Bipolaris and Drechslera, for example, effect of Trichoderma harzianum on sporulation of Cochliobolus sativus on excised wheat seedling leaves[11], potential biocontrol of Bipolaris sorokiniana on phyloplane of Poa pratensis with strains of Pseudomonas sp.[3], antagonistic effect of yeast against Cochliobolus sativus on wheat leaves[12]. There is no or little information on the efficacy of Trichoderma species against common root rot of wheat caused by Bipolaris sorokiniana.
The objective of this investigation was to evaluate the potential of Trichoderma viride (MO), T. viride T112, T. harzianum (M) and T. harzianum T191 for biological control of common root rot of wheat.
MATERIALS AND METHODS
Pathogen and antagonist isolates: Trichoderma viride isolate T112 and T. harzianum T114 isolated from soil surrounding of wheat of Shahzand Arak in Markazi province of Iran and T. viride (MO) and T. harzianum (M) obtained from Dr. Rouhani Bo-Alisina University, Hamedan. Isolates SH and KH of Bipolaris sorokiniana obtained by Eng. Ghalandar, Agricultural Research Center of Markazi Province were used in this study. All isolates were maintained on potato dextrose agar at 5°C.
Effect of Trichoderma isolates on mycelial growth of B. sorokiniana in vitro: Dual culture[13] and cellophane overlays[14] were used to observe the effect of Trichoderma isolates on B. sorokiniana. All antagonist-pathogen combinations were examined on 10-15 mL of potato dextrose agar in 9 cm petri plates with 4 replicate plates per treatment. For dual culture mycelial plug (5 mm in diameter) taken from actively growing, 3 days old colonies of B. sorokiniana isolate or Trichoderma isolate were placed 5 cm apart on the agar, controls consisted of pure cultures.
For cellophane overlays, cellophane membranes (Australia Cellophanes, Victoria), 9 cm in diameter were boiled in distilled water, then interleaved with filter paper and autoclaved before being placed on the medium. One 5 mm diameter plug of Trichoderma species growing on PDA was placed on the center of each cellophane membrane. For controls, a plug of sterile PDA medium was used instead of the antagonist. The cellophane membrane and adhering fungus, or agar plug were removed after 2 days[9]. A plug of B. sorokiniana was placed on the agar in the center of plate and incubated for 6 days.
For antifungal activity of volatile, a petri plate containing PDA medium was inoculated with 5 mm diameter plug of Trichoderma isolates growing on PDA. A second petri plate containing PDA was inoculated with a 5 mm plug of the B. sorokiniana in the center of the plate and inverted over the Trichoderma culture. The two plates were sealed together with nescofilm and incubated at 25°C for 6 days. This ensured that both organisms were growing on the same atmosphere. For control instead of Trichoderma one plug of PDA was placed on agar surface[15].
The surface area of the colonies B. sorokiniana was recorded compared with controls and the percentage of growth inhibition was calculated. All experiments in vitro were arranged as Randomized Complete Design with 4 replications.
Biological control of B. sorokiniana on wheat in glasshouse condition
Seed treatment: The ability of Trichoderma isolates to reduce incidence of common root rot of wheat in glasshouse was investigated. Inoculum of the pathogen was prepared as follows. Wheat seeds were soaked for 16 h and then transferred to 125 erlenmyer flask and autoclaved for 1 h at 121°C on two successive days. B. sorokiniana isolates KH and SH were grown separately on Potato Dextrose Agar (PDA) and when they were grown 5 pieces of culture about 5x2 cm in size were added to each erlenmyer flask containing autoclaved wheat, mixed with wheat and incubated at 25°C for 30 days. The infested seeds were air-dried, powdered by grinding, then were added to autoclaved soil as a rate of 5%(w/w) and placed in 14 cm pots.
To obtain conidial suspension of Trichoderma, the fungus was grown on 9 cm petri plate for 7 days at 25°C. Conidia were harvested from surface of plates and washed several times in Sterile Distilled Water (SDW) and suspended in 0.01% tween 20. Seeds of cv. Alvand were surface disinfected by soaking in 0.5% sodium hypochlorite for 3 min then rinsed three times in SDW. The seed were soaked for 1 h in Trichoderma spore suspension and air dried in a laminar flow hood. For adhering of conidia to seed surface methyl cellulose was used[16]. The number of Trichoderma colonies was determined after 2 days of incubation at 25°C. Five Trichoderma inoculated seeds were sown in each pot. Plants were maintained in the glasshouse at 22-25°C without supplementary lighting from April to May (spring). Pots were watered at 3 days intervals until emergence and daily thereafter. Forty five days after planting, plant height, fresh and dry weight of shoots and roots and percentage of infected roots were determined. Sample of tissue were taken from the root had discoloured. Segments of tissue 5 mm long, were surface-disinfected in 50% domestic bleach (0.5% available chorine) for 3 min, rinsed three times in SDW and plated on the PDA rose bengal peptone agar.
Soil treatments: The methods and materials for soil treatments were the same as described for seed treatment. In this experiment, density of conidia of Trichoderma was adjusted to 107 mL-1 with a hemacytometer. Seventy milliliter of spore suspension was added in each pot after planting of seeds. All experiments in glasshouse were arranged as randomized complete design with 4 replicate pots.
Statistical analysis: Data on percentage inhibition of growth and percentage of infected roots were subjected to arcsin square root transformation before analysis. Data for plant height, fresh and dry weight of shoot and root were analysed directly. Analysis of variance was performed and means were separated using Duncan’s Multiple Range Test at p<0.05[17].
RESULTS AND DISCUSSION
Effect of Trichoderma isolates on mycelial growth of B. sorokiniana: Dual culture, cellophane overly technique and volatile metabolite test showed that all isolates of Trichoderma tested inhibited the growth of 2 isolates of Bipolaris sorokiniana (Table 1). The inhibition varied from 29.56 to 69.82% in dual culture, from 66.66 to 98.25% in volatile metabolite test and from 58.20 to 93.93% using the cellophane overlay method. Results of dual culture, cellophane overlay indicated that mycelial growth of B. sorokiniana was numerically reduced more by T. viride T112 than the others isolate tested and also the percentage of growth inhibition of the pathogen by T. viride T112 was not less than 85% by means of volatile metabolite test.
Effect of Trichoderma isolates on control of common root rot of wheat in glasshouse: The results of seed-soaking with Trichoderma experiments indicated that common root rot was significantly reduced by Trichoderma viride T112 and T. viride (MO). For example, calculated mean in disease severity caused by isolate SH of the pathogen with these two antagonists were 31.50 and 27.75%, respectively compared to pathogen control (75.75%). All Trichoderma isolates used in this test reduced disease severity compared to control (Table 2). Plant height, dry weight and fresh weight of shoots and roots in treatment which the seed inoculated with Trichoderma was greater than pathogen control and also four Trichoderma isolates promoted plant growth and suppressed the B. sorokiniana (Table 2). B. sorokiniana was isolated from all inoculated plants showing symptoms.
All the Trichoderma isolates suppressed (p<0.05) common root rot severity when added to soil infested with B. sorokiniana compared to disease caused by B. sorokiniana alone. Isolate T. viride T112 and T. viride (MO) were more effective in reducing common root rot than the other isolates tested. Soil treatment with Trichoderma also promoted plant growth (Table 3).
The isolates of T. harzianum T114 and T. viride T112 which were isolated from soil surrounding of wheat in Arak of Iran and two isolates T. harzianum (M) and T. viride (MO) obtained from Dr. Rouhani, were tested for their antagonistic activity in vitro and in vivo. These isolate have not previously been evaluated for potential antagonists to B. sorokiniana.
| Table 1: | Effect of Trichoderma isolates on mycelial growth of B. sorokiniana as % of colony area compared to control |
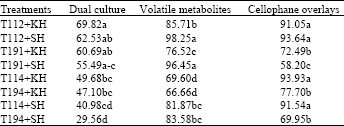 | |
| Significant differences are denoted by different letters within each column according to Multiple Range Test at p<0.01. Data are expressed as % of control colonies without antagonist and values are average of 4 replicates. Data were subjected to arcsin square root transformation prior to analysis of variance, T112= T. viride, T194= T. viride (MO), T114=T. harzianum, T191= T. harzianum (M), KH= Bipolaris sorokiniana isolated from wheat field of Khomain, Iran, SH= B. sorokiniana isolated from wheat field of Shahzand, Iran | |
| Table 2: | Effect of Trichoderma isolates on control of Bipolaris sorokiniana at glasshouse condition (seed treatment test) |
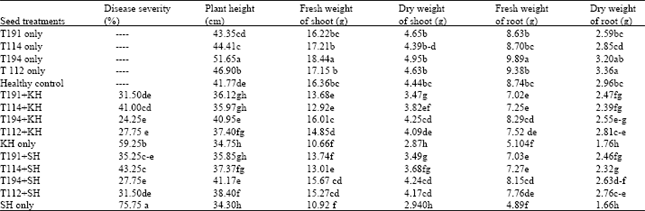 | |
| Data represent the mean of 4 replicates, Significant differences are denoted by different letters within each column according to Multiple Range Test at p<0.01. T112= Trichderma viride, T194= Trichoderma viride (MO), T114=T. harzianum, T191= T. harzianum (M), KH= Bipolaris sorokiniana isolated from wheat field of Khomain, Iran, SH= B. sorokiniana isolated from wheat field of Shahzand, Iran | |
| Table 3: | Effect of Trichoderma isolates on control of Bipolaris sorokiniana at glasshouse condition(soil treatment test) |
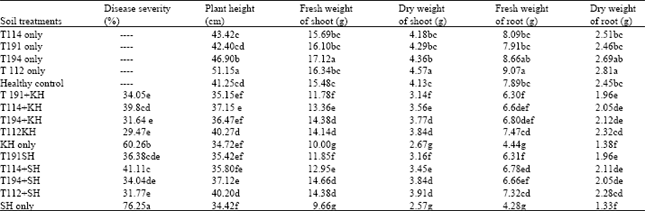 | |
| Data represent the mean of 4 replicates, Significant differences are denoted by different letter(s) within each column according to Multiple Range Test at p<0.01. T112= Trichoderma viride, T194= Trichoderma viride (MO), T114=T. harzianum, T191= T. harzianum (M), KH= Bipolaris sorokiniana isolated from wheat field of Khomain, Iran, SH= B. sorokiniana isolated from wheat field of Shahzand, Iran | |
All isolates reduced mycelial growth of Pathogen by means dual culture, cellophane overlay technique and volatile metabolite test. In most cases the effectiveness of T. viride T112 isolated from soil surrounding of wheat was superior to the other isolates and uniform among the above tests. Biocontrol activity of antagonist fungi and bacteria may partially be associated with production of antibiotic[9,18-20]. For example, the mode of action of T. harzianum appeared to be antagonism by the production Isonitrin, homothallin II, melanoxadin[18,21,22] and the other antibiotics Trichdermin[23], ergokonin[24], viridin[25,26] viridiofungin A, B and C[27] produced by different isolates of Trichoderma viride have been involved in biological control. However it has not been possible to extract these substances from T. harzianum and T. viride tested. The metabolites produced by T. viride T112 and T. viride T194 that provided biocontrol in these experiments is likely similar to metabolite produced by the other isolates tested by the other investigators mentioned above. Further research is needed to identify these metabolites and their properties.
T. viride and T. harzianum reduced disease severity of common root rot of wheat. Some investigator have tried to use Tirichoderma isolate and the other fungi for biocontrol of disease caused by Cochliobolus sativus[11] and Drechslera sorokiniana on wheat and rye leaves[28] and also successful antagonists against seed-borne B. sorokinina were Chaetomium sp., Idriella bolleyi and Gliocladium roseum[29]. Suppression of soil-borne fungi, including B. sorokiniana has been observed in the presence of Isothiocyanates released into soil by Brassica species[30]. Reduced symptoms could also be achieved, although not to be the full extent, after spraying with the bacterial biocontrol agent Pseudomonas chloraphis strain MA 342[31]. Hogdes et al.[3] indicated that Pseudomonas PSD-42 was antagonistic to B. sorokiniana on leaves of Poa pratensis. Plant height, fresh and dry weight of shoots and roots of wheat seedling from soil and seed treated with 2 isolates of Trichoderma viride plus B. sorokiniana were greater than those treated with pathogen alone. This study supports previous results showing that some Trichoderma isolates are capable of increasing plant growth and yield in green house[9,32-34].
Inoculation of soil and seed in the glasshouse with all four isolates of Trichoderma alone increased plant height, fresh and dry weight of shoots and roots compared with controls without Trichoderma. This is in accordance with results showing that some fungal isolates and bacterial strains are capable of promoting plant growth in greenhouse[4,35-37]. One of the mechanisms of disease suppression by Trichoderma isolates is competition with pathogen for infection site on the root surface. The degree of root colonization by the Plant Growth Promoting Fungal (PGPF) isolates depended upon the amount of pathogen inoculum present. The PGPF was found to colonize the epidermal and outer cortical cell layers of roots.
Another possible mechanism for suppression of common root rot could be induced resistance in wheat plants due to root colonization by PGPF isolates[4]. The production of pathogen inhibitory substance by Trichoderma during root colonization cannot be ruled out. Plant growth promotion due to PGPF has been attributed to the increase of mineral nutrient availability to roots during the growth activity of PGPF isolates in the rhizosphere. Induction of local or systemic resistance in wheat to pathogen following PGPF root colonization is yet another possible explanation for the suppression of common root rot. However the role(s) of other mechanisms in disease suppression cannot be rule out.
In conclusion T. viride T112 and T. viride (MO) tested here reduced severity in wheat in the glasshouse. Future research will involve studies of the mechanisms involved. The isolates warrant further investigation for their ability to control common root rot of wheat, especially in commercial situations. An integrated approach using a combination of biocontrol agents and the fungicides which does not affect mycelial growth of these antagonists may allow reduction in the amount of fungicide needed to suppress common root rot.
ACKNOWLEDGMENTS
The research presented here is part of M.Sc Thesis at Department of plant protection, Abourayhan Campus, University of Tehran.
REFERENCES
- Murray, T.D., D.W. Parry and N.D. Catlin, 1998. A Color Book of Disease of Small Grain Cereal Crop. Mannson Publishing, UK., pp: 145.
Direct Link - Di Pietro, A., M. Loro, C.K. Hayes, R.M. Broadway and G.E. Harman, 1993. Endochitinase from Gliocladium virens: Isolation, characterization and synergistic antifungal activity in combination with gliotoxin. Phytopathology, 83: 308-313.
Direct Link - Elad, Y., 1994. Biological control of grape grey mould by Trichoderma harzianum. Crop Prot., 13: 35-38.
CrossRefDirect Link - Harman, G.E., B. Latorre, B. Agosin, R. San Martin and D.G. Riegel et al., 1996. Biological and integrated control of Botrytis bunch rot of grape using Trichoderma sp. Biol. Control, 7: 259-266.
CrossRef - Lumsden, R.D., J.F. Walter and C. Baker, 1996. Development of Gliocladium virens for damping-off disease control. Can. J. Plant Pathol., l8: 463-468.
Direct Link - Biles, C.L. and J.P. Hill, 1988. Effect of Trichoderma harzianum on sporulation of Cochliobolus sativus on excised wheat seedling leaves. Phytopathology, 78: 656-659.
Direct Link - Dennis, C. and J. Webster, 1971. Antagonistic properties of species-groups of Trichoderma: I. Production of non-volatile antibiotics. Trans. Br. Mycol. Soc., 57: 25-39.
CrossRefDirect Link - Fiddaman, P.J. and S. Rossall, 1993. The production of antifungal volatile by Bacillus subtilis. J. Applied Bacteriol., 74: 119-126.
Direct Link - Weller, D.M. and R.J. Cook, 1983. Suppression of take-all of wheat by seed treatments with fluorescent pseudomonads. Phytopathology, 73: 463-469.
CrossRefDirect Link - Little, T.M. and F.J. Hills, 1978. Agricultural Experimentation: Design and Analysis. John Wiley and Sons, Hoboken, New Jersey, ISBN-13: 9780471023524, Pages: 350.
Direct Link - Howell, C.R. and R.D. Stipanovic, 1979. Control of Rhizoctonia solani on cotton seedling with Pseudomonas fluorescens and with an antibiotic produced by bacterium. Phytopathology, 69: 480-482.
Direct Link - Pusey, P.L. and C.L. Wilson, 1984. Postharvest biological control of stone fruit brown rot by Bacillus subtilis. Plant Dis., 70: 587-590.
Direct Link - Lee, C.H., M.C. Chang, H.J., Lee, Y.H. Kho and H. Koshiso, 1995. MR-93A, A new oxazole from Trichoderma harzianum KCTC 0114BP. J. Natl. Prod., 58: 1605-1607.
Direct Link - Lee, C.H., H. Koshino, M.C. Chang, H.J. Lee and Y.H. Kho, 1995. MR-304A, a new melanin synthesis inhibitor from the fungus Trichoderma harzianum. J. Antibiot., 48: 1168-1170.
Direct Link - Grove, J.F., J.S. Moffatt and E.B. Vischer, 1965. Viridin. Part I. Isolation and characterization. J. Chem. Soc., 1965: 3803-3811.
CrossRef - Harris, G.H., E.T.T. Jones, M.S. Meinz, M. Nallin-Omstead and G.L. Bills et al., 1993. Isolation and structure elucidation of viridiofungins A, B and C. Tetrahedron Lett., 34: 5235-5238.
Direct Link - Fokkema, N.J., 1973. The role of saprophytic fungi in antagonism against Drechslera sorokiniana (Helminthosporium sativum) on agar plates and on rye leaves with pollen. Physiol. Plant Pathol., 3: 195-205.
CrossRefDirect Link - Kirkegaard, J.A., P.T.W. Wong and J.M. Desmarchelier, 1996. In vitro suppression of fungal root pathogen of cereal by Brassica tissues. Plant Pathol., 45: 593-603.
Direct Link - Johnseon, L., M. Hokeberg and B. Gerhardson, 1998. Performance of the Pseudomonas chlororaphis biocontrol agent MA 342 against cereal seed-borne diseases in field experiments. Eur. J. Plant Pathol., 104: 701-711.
CrossRefDirect Link - Chang, Y.C., Y.C. Chang, R. Baker, O. Kleifeld and I. Chet, 1986. Increased growth of plants in the presence of the biological control agents Trichoderma harzianum. Plant Dis., 70: 145-148.
Direct Link - Seuk, B.Y., J.S. Sik, P.C. Seuk and K.H. Kyu, 1995. In vitro and greenhouse evaluation of cucumber growth enhanced by rhizosphere microorganisms. Kor. J. Plant Pathol., 11: 292-297.
Direct Link - El-Abyad, M.S., M.A. El-Sayed, A.R. El-Shanshoury and N.H. El-Batanouny, 1993. Inhibitory effects of UV mutants of Streptomyces corchorusii and Streptomyces spiroverticillatus on bean and banana wilt pathogens. Can. J. Bot., 71: 1080-1086.
CrossRefDirect Link - El-Tarabily, K.A., M.L. Sykes, I.D. Kurtboke, G.E.S.J. Hardy and A.M. Barbosa et al., 1996. Synergistic effects of a cellulase-producing Micromonospora carbonacea and an antibiotic-producing Streptomyces violascens on the suppression of Phytophthora cinamomi root rot of Banksia grandis. Can. J. Bot., 74: 618-624.
- Jones, C.R. and D.A. Samac, 1996. Biological control of fungi causing alfalfa seedling damping-off with a disease-suppressive strain of Streptomyces. Biol. Control, 7: 196-204.
CrossRefDirect Link