
Yanar Yusuf
Department of Plant Protection, Agricultural Faculty, Gaziosmanpasa University, Tokat 60250, Turkey
Miller Sally
Department of Plant Pathology, Ohio State University, OARDC, Wooster, OH, 44691, USA
Plant Pathology Journal
Year: 2004 | Volume: 3 | Issue: 1 | Page No.: 52-55







ABSTRACT
This study was carried out to determine the effect of CGA 245704 (a Benzothiadiazole derivative) and two non-pathogenic Xanthomonas campestris strains on white mold disease of pepper caused by Sclerotinia sclerotiorum. Pretreatment of pepper seedlings with the bacterial strains did not reduce the disease severity of white mold. Application of CGA 245704 followed by a subsequent inoculation of the plants with the fungus resulted in significant decrease in disease severity. No inhibition of ascospore germination of Sclerotinia sclerotiorum isolate Ss6 but reduced germ tube length, radial growth of mycelium and number of sclerotia on culture medium amended with CGA 245704 was observed. The decrease in disease severity following treatment with CGA 245704 might be the result of both resistance induction and fungistatic effect.
PDF Abstract XML References
How to cite this article
Yanar Yusuf and Miller Sally, 2004. Effect of Treating Pepper with Benzothiadiazole Derivative
and Non-pathogenic Xanthomonas campestris Pathovars on the Level of Disease
Caused by Sclerotinia sclerotiorum. Plant Pathology Journal, 3: 52-55.
DOI: 10.3923/ppj.2004.52.55
URL: https://scialert.net/abstract/?doi=ppj.2004.52.55
DOI: 10.3923/ppj.2004.52.55
URL: https://scialert.net/abstract/?doi=ppj.2004.52.55
INTRODUCTION
The phenomenon of systemic acquired resistance (SAR), in which resistance to disease is enhanced in tissues distant from the site of the prior inducing treatment, has been extensively reported for a number of plant pathogen systems[1,5]. Biochemical studies have shown that SAR responses are correlated with the accumulation of some pathogenesis related (PR) proteins including β-1,3-glucanases, chitinases, cysteine rich proteins related to thaumatin and the PR-1 proteins[6,7]. Accumulation of these compounds in plants is stimulated by bacteria[7]. Bacterial induction of systemic resistance by plant growth promoting rhizobacteria (PGPR) has been demonstrated in several plant species[8-11]. Liu et al.[9] reported that two PGPRs, Pseudomonas putida strain 8913-27 and Serrotia marcescens strain 90-166, induced systemic resistance in cucumber against Colletotrichum orbiculare (causal agent of cucumber anthracnose). Induction of SAR in cucumber and rice by pathogenic and non-pathogenic bacteria was demonstrated[4]. Inoculation of the first leaf of rice plant with Pseudomonas syringae pv. syringae induced systemic resistance in the second leaf of the plant against the fungal pathogen Pyricularia oryzae[4]. Infiltration of the first leaf of cucumber plants with a bacterial suspension of P. syringae pv. syringae (nonpathogenic, elicit hypersensitive response in cucumber) or P. syringae pv. Lachrymans (cucumber pathogen) induced systemic resistance in cucumber plants against a subsequent inoculation by Colletotrichum lagenarium[3].
The treatment of plants with certain synthetic chemical agents can also induce the SAR response [7,12-15]. Many compounds provide good protection against fungal and bacterial pathogens of many crops in greenhouse as well as in field conditions. The synthetic chemicals Benzothiadiazole (BTH) and 2,6-dichloroisonicotinic acid (INA) have been used for induction of SAR in wheat, bean, soybean and barley against fungal and bacterial pathogens[16-19]. Kogel et al.[19] reported that INA (CGA 41396) induced resistance in susceptible barley (Hordeum vulgare L.) cultivars to powdery mildew (Erysiphe graminis f.sp hordei). BTH, which was developed by Ciba-Geigy Corp. (Novartis) (Basel, Switzerland), has been tested under the experimental name CGA 245704 (benzo (1,2,3) thiadiazole-7-carbothioic acid S methyl ester), triggers SAR in several crops[18,20,21]. The effect of this compound on induction of SAR to Sclerotinia sclerotiorum has not been reported for pepper. However, Dann et al.[22] reported that three or four applications of INA or BTH reduced white mold severity and increased yield in two highly susceptible soybean cultivars under greenhouse and field conditions. In the present study the effects of this compound and of two non-pathogenic Xanthomonas spp., when applied as foliar spray for protecting pepper against Sclerotinia sclerotiorum were investigated.
MATERIALS AND METHODS
Ascospore production: Sclerotia produced on autoclaved carrot discs were surface sterilized in 95 % ethanol for 90 s as described by Huang and Kozub[23] and air dried for 15 min in a laminar flow hood. Twenty sclerotia were placed in 9 cm-diameter glass petri plates containing 15 mL sterile distilled water. Plates were wrapped with parafilm and conditioned at 4°C for 6 wk in the dark. They were then incubated at 15°C under fluorescent light (47.5 E s-1 m-2) for 14 h followed by 10 h dark/day. Stipes formed on sclerotia 4 weeks after transferring to 15°C and mature apothecia developed after 1 to 2 weeks. Ascospores were collected from mature apothecia directly on nucleopore polycarbonate filters (type GS, 47 μm, Thomas Scientific, Swedesboro, NJ) as described by Hunter and Cigna[24] and Steadman and Cook[25]. Filters with ascospores were stored in glass jars containing calcium chloride (CaCl2) as a desiccant at -19°C or at 4°C until used.
Pepper cultivar, growing conditions and induction treatments: The susceptible bell pepper cultivar Marengo was used in this study. Ascospores of Sclerotinia sclerotiorum isolate Ss6 were used for inoculation. Seeds were sown directly into 10 cm pots filled with sterilized soil mixture consisting of 0.43 m3 silty-loam, 0.43 m3 muck soil, 0.14 m3 peatmoss and 1,2 kg lime m-3 and allowed to germinate on greenhouse benches where the air temperature was maintained 28±2°C during day and night. After 3 weeks, pots were thinned to contain six seedlings that had one true leaf each. Plants were fertilized with peter’s 20-20-20 soluble N-P-K (454 g 378 L-1) solution weekly and watered once a day. A strain of Xanthomonas campestris (Xc 19), isolated from pepper (non-pathogenic), a necrotizing strain of Xanthomonas campestris pv. raphani isolated from tomato and the synthetic chemical CGA 245704 were tested for their SAR inducing activity in pepper against Sclerotinia sclerotiorum. Treatments were: 1) plant sprayed with CGA 245704 or each of the bacterial strains alone; 2) plants sprayed with CGA 245704 or each of the bacterial strains plus an ascospore suspension of the fungus 3) plants sprayed with an ascospore suspension of the pathogen alone and plants sprayed with deionized water alone.
Pepper seedlings were sprayed with CGA 245704 (125 mg a.i. L-1 water) or bacterial suspensions (108 cfu mL-1) 5 weeks after sowing. One week later the sprays were repeated. Seven days after the second spray, seedlings were sprayed with an ascospore suspension of the isolate Ss6 (5x104 ascospores mL-1) or distilled water.
| Table 1: | Development of Sclerotinia stem rot in pepper seedlings after foliar application of CGA 245704 or inoculation with nonpathogenic Xanthomonas campestris strain Xc19 or Xanthomonas campestris pv. raphani strain DC-91-1 |
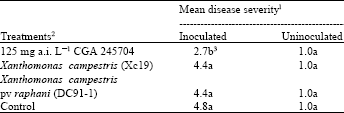 | |
| 1Disease severity was estimated 15 days after inoculation on a scale of 1 to 4 where 1= no symptoms, 2= water-soaking of petioles and leaves or restricted lesions on leaves, 3= small arrested lesions on main stem, 4= lesions encircled the main stem, and 5= lesions on the main stem resulting in stem collapse and plant death 2Initial treatment applied 5 weeks after seeding, second treatment applied one week after initial one. 3Means within a column followed by the same letter are not significantly different according to Fisher’s least significant difference test,P= 0.05 | |
| Table 2: | Effects of CGA 245704 treatment on the germ tube length, mycelial growth and sclerotia production of Sclerotinia sclerotiorum |
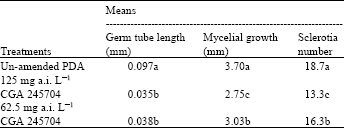 | |
| Means with in a column followed by the same letter are not significantly different according to Fisher’s least significant difference test, P=0.05` | |
Pots were placed on greenhouse bench at 20±2°C. Each treatment had five replicates. Pots were arranged in a completely randomized design. Relative humidity was maintained near 100% by a mist system that sprayed plants for 5 min h-1 during the incubation period. Plants were rated for Sclerotinia stem rot disease severity 15 days after inoculation using a 1-5 scale, in which 1= no symptoms, 2= water soaking of petioles and leaves or restricted lesions on leaves, 3= small arrested lesions on the main stem, 4= lesion encircling the main stem and 5= lesion on the main stem resulting in stem collapse and/or plant death.
Mycelial growth and sclerotia production on amended PDA: Sclerotinia sclerotiorum isolate Ss6 were assessed for mycelial growth and sclerotia production on non-amended and medium amended with 125 and 62.5 mg a.i. L-1 of CGA 245704. CGA 245704 was added at 0, 125, 62.5 mg a.i. L-1 to autoclave-sterilized potato dextrose agar (PDA) medium cooled to 50°C. The plates were inoculated in the center with a 0.5 mm diameter plug of mycelium removed from the actively growing edge of the culture (isolate Ss6) previously grown for 2 days at 24°C in darkness and radial growth was then measured. Each treatment contained 4 replicates. The test was performed three times.
The effect of CGA245704 on sclerotia production was determined by counting the number of sclerotia produced on 25 day old cultures on PDA amended with CGA 245704 and on unamended PDA.
Ascospore germination: To test the percentage of germination of ascospores, nucleopore filters containing ascospores of Sclerotinia sclerotiorum isolate Ss6 were back washed with sterile distilled water and the final ascospore concentration was adjusted to 5x104 spores mL-1, a drop of ascospore suspension was placed on to the surfaces of non-amended PDA and PDA amended with CGA 245704 (125 and 0.63 mg a.i. L-1). After 10 h incubation at room temperature (21-23°C), 100 ascospore per plate were examined for germination. An ascospore was considered germinated if the length of the germ tube was greater than the width of the ascospore. Germ tube length of ascospores was measured at the end of incubation period. The experiment was conducted three times with four replicates per treatment and data were analyzed by analysis of variance (ANOVA) procedures (SAS Institute, Cary, NC). Means were compared using Fisher’s least significant difference.
RESULTS AND DISCUSSION
Development of Sclerotinia stem rot in pepper seedlings after foliar applications of CGA 245704 or inoculation with the nonpathogenic Xanthomonas campestris strain Xc19 or X. campestris pv. raphani strain DC-91-1 is shown in Table 1. Pre-treatment of pepper seedlings with CGA 245704 reduced disease severity (2.73 mean disease severity) significantly compared to non-treated control (mean disease severity of 4.8). None of the bacterial strains tested reduced disease severity in pepper.
At a concentration of 125 and 62.5 mg a.i. L-1 CGA 245704 25% WG significantly inhibited mycelial growth of Sclerotinia sclerotiorum isolate Ss6. The number of sclerotia produced by this isolate in the presence of CGA245704 was significantly lower than the control (Table 2), but no clear difference was detected in the size of sclerotia formed at concentrations of 125 and 62.5 mg a.i. L-1 CGA245704. Sclerotia of isolate Ss6 were quite variable in size and shape, which made the detection of changes in size difficult.
The CGA245704 treatment did not inhibit ascospore germination of Sclerotinia sclerotiorum isolate Ss6 at 125 and 62.5 mg a.i. L-1 concentrations. The number of germinating ascospore was similar to the one observed on un-amended PDA. However, 90-95% of ascospores on amended medium germinated from both ends while ascospores on un-amended PDA germinated from one end. Germ tube length was measured after 10 hours of incubation showed that there was a significant difference between the length measured on amended and un-amended medium (Table 2).
Sclerotinia sclerotiorum disease severity was reduced when pepper seedlings were inoculated one week after spraying with 125 mg a.i. L-1 CGA 245704 and a strong local and systemic protection was observed. The result is in agreement with findings of Dann et al.(17) that showed significant control of Sclerotinia sclerotiorum in soybean. In the PDA medium amended with CGA 245704 at concentrations of 125 and 62.5 mg a.i. L-1 a significant reduction in germ tube length , radial growth of mycelium and sclerotia number occurred. Therefore, it was concluded that CGA 245704 had some fungistatic effect on Sclerotinia sclerotiorum isolate Ss6 at the concentrations used in this study. The results are different from those obtained by other groups(15,22,26) showing that the compound had no antifungal activity. This is possibly due to a difference in the concentrations of the inducer and the pathogen tested, suggesting that the activity of CGA 245704 could differ from one plant-pathogen system to another according to the dose used.
REFERENCES
- Van Loon, L.C., P.A.H.M. Bakker and C.M.J. Pieterse, 1998. Systemic resistance induced by rhizosphere bacteria. Annu. Rev. Phytopathol., 36: 453-483.
CrossRefPubMedDirect Link - Kessmann, H., T. Staub, C. Hofmann, T. Maetzke and J. Herzog et al., 1994. Induction of systemic aquired resistance in plants by chemicals. Ann. Rev. Phytopathol., 32: 439-459.
CrossRef - Leeman, M., J.A. van Pelt, F.M. Denouden, M. Heinsbroek, P.A.H.M. Bakker and B. Schippers, 1995. Induction of systemic resistance against fusarium wilt of radish by lipopolysaccharides of P. fluorescens. Phytopathology, 85: 1021-1027.
Direct Link - Liu, L., J.W. Kloepper and S. Tuzun, 1995. Induction of systemic resistance in cucumber against fusarium wilt by plant growth-promoting rhizobacteria. Phytopathology, 85: 695-698.
CrossRefDirect Link - Van Peer, R., G.J. Niemann and B. Schippers, 1991. Induced resistance and phytoalexin accumulation in biological control of Fusarium wilt of carnation by Pseudomonas sp. strain WCS417r. Phytopathology, 81: 728-734.
CrossRefDirect Link - Wei, G., J.W. Kloepper and S. Tuzun, 1991. Induction of systemic resistance of cucumber to Colletotrichum orbiculare by select strains of plant growth-promoting rhizobacteria. Phytopathology, 81: 1508-1512.
CrossRefDirect Link - Brisset, N.M., S. Cesbron, S.V. Thomson and J.P. Paulin, 2000. Acibenzolar-S-methyl induces the accumulation of defense-related enzymes in apple and protects from fire blight. Eur. J. Plant Pathol., 106: 529-536.
CrossRef - Tosi, L., R. Luigetti and A. Zazzerini, 1999. Benzothiadiazole induces resistance to Plasmopara helianthi in sunflower plants. J. Phytopathol., 147: 365-370.
CrossRef - Dann, E.K. and B.J. Deverall, 1995. Effectiveness of systemic resistance in bean against foliar and soilborne pathogens as induced by biological and chemical means. Plant Pathol., 44: 458-466.
CrossRef - Gorlach, J., S. Volrath, G. Knauf-Beiter, G. Hengy and U. Becknove et al., 1996. Benzothiadiazole a noval class of inducers of systemic acquired resistance, activates gene expression and disease resistance in wheat. Plant Cell, 8: 629-643.
PubMed - Kogel, K.H., U. Beckhove, J. Dreschers, S. Munch and Y. Romme, 1994. Acquired resistance in barley the resistance mechanism induced by 2 6 dichloroisonicotinic acid is a phenocopy of a genetically based mechanism governing race specific powdery mildew resistance. Plant Physiol., 106: 1269-1277.
Direct Link - Dann, E., B. Diers, J. Byrum and R. Hammerschmidt, 1998. Effect of treating soybean with 2,6-dichloroisonicotinic acid (INA) and benzothiadiazole (BTH) on seed yields and the level of disease caused by Sclerotinia sclerotiorum in field and greenhouse studies. Eur. J. Plant Pathol., 104: 271-278.
CrossRef - Huang, H.C. and G.C. Kozub, 1989. A simple method for production of apothecia from sclerotia of Sclerotinia sclerotiorum. Plant Prot. Bull., 31: 333-345.
Direct Link - Hunter, J.E. and J.A. Cigna, 1982. Preservation of ascospores of Sclerotinia sclerotiorum on membrane filters. Phytopathology, 72: 650-652.
Direct Link - Bigirimana, J. and M. Hofte, 2002. Induction of systemic resistance to Colletotrichum lindemuthianum in bean by a benzothiadiazole derivative and rhizobacteria. Phytoparasitica, 30: 159-168.
Direct Link - Metraux, J.P., P. Ahl-Goy, T. Staub, J. Speich, A. Steinemann, J. Ryals and E. Ward, 1991. Induced Resistance in Cucumber in Response to 2, 6-dichloroisonicotinic Acid and Pathogens. In: Advances in Molecular Genetics of Plant-Microbe Interactions, Henneke, H. and D.P.S. Verma (Eds.). Kluwer Dordecht, The Netherlands, pp: 432.
CrossRef