
Kiros Meles
Tigrai Agricultural Research Institute, P.O. Box 492, Mekele, Ethiopia
M. Hulluka
Alemaya University of Agriculture, P.O. Box 138, Dire Dawa, Ethiopia
M.M. Abang
Cereal Pathology Laboratory, Germplasm Program,
International Center for Agricultural Research in the Dry Areas (ICARDA), P.O. Box 5466 Aleppo, Syria
Plant Pathology Journal
Year: 2004 | Volume: 3 | Issue: 1 | Page No.: 26-34







ABSTRACT
Knowledge of Rhynchosporium secalis variability and host response to scald is important in the context of breeding for resistance. The variation in R. secalis isolates from barley in different agroecological zones of Ethiopia was investigated with respect to colony and conidial morphology, colony growth rate, sporulation and virulence spectrum on a set of ten barley differentials. The R. secalis isolates differed markedly in several cultural characteristics but no correlation was found between these characteristics and isolate virulence. Five R. secalis pathotypes were selected from a total of 19 pathotypes and used to screen 35 barley lines in a glasshouse and also under natural infection in the field. Discriminant analysis revealed considerable divergence in host response, as well as in R. secalis virulence. Percent leaf area affected (PLAA) and area under the disease progress curve (AUDPC) were negatively correlated with grain yield and 1000-grain weight and more closely reflected host resistance than apparent infection rate (r). High yielding barley lines such as HB-100 that showed resistance to all five pathotypes in the glasshouse and had low AUDPC, r and PLAA values in the field appear promising as donors of quantitative resistance genes in scald resistance breeding in the high altitude zones of Ethiopia.
PDF Abstract XML References
How to cite this article
Kiros Meles, M. Hulluka and M.M. Abang, 2004. Phenotypic Diversity in Rhynchosporium secalis from Ethiopia and Host
Response to Barley Scald. Plant Pathology Journal, 3: 26-34.
DOI: 10.3923/ppj.2004.26.34
URL: https://scialert.net/abstract/?doi=ppj.2004.26.34
DOI: 10.3923/ppj.2004.26.34
URL: https://scialert.net/abstract/?doi=ppj.2004.26.34
INTRODUCTION
Barley (Hordeum vulgare L.) is the most important food crop in Ethiopia after tef (Eragrostis tef [Zucc] Trotter) and sorghum (Sorghum bicolor [L.] Moench). It is a traditional crop in the highlands of Ethiopia grown over a wide range of climatic conditions. Scald, caused by Rhynchosporium secalis (Oud) Davis, is the most widely distributed and destructive disease of barley in Ethiopia[1], which is one of the centres of diversity of cultivated barley. The disease is most severe in the highlands (above 2000 m) where precipitation is high and temperature is low during the cropping season. Reported losses in yield due to scald vary between 21-67% depending on season and cultivar[2].
Cultural measures to control the disease are usually ineffective when disease pressure is high and chemical control strategies are generally inappropriate for the mostly resource-poor farmers. Genetic resistance could potentially form the basis of sustainable management strategies for barley scald; however, screening and selection of barley genotypes for resistance to the disease is currently hampered by the dearth of knowledge on the variability of the pathogen in Ethiopia. Pathogenic variation in R. secalis and corresponding variability in host resistance have been studied by workers in several countries[3-5] but little is known about the variability of the pathogen or host response to R. secalis pathotypes in Ethiopia. Yitbarek[6] observed that some scald-resistant barley lines were also high- yielding across locations in Ethiopia, whereas other scald-resistant lines were low- yielding. This suggests that interactions between foliar diseases of barley may have a considerable influence on the field performance of scald-resistant cultivars. Although studies have been conducted on the effect of scald on yield and yield parameters, little is known about the relative merits of parameters to characterize severity levels of scald observed during the growing season on individual barley genotypes in Ethiopia.
R. secalis is a deuteromycete fungal pathogen with uninucleate hyphal and conidial cells and no known sexual stage[7]. The lack of a recognized sexual stage precludes standard Mendelian analysis of the genetics of host-pathogen interaction. The pathogen apparently posseses limited mechanisms for generation of variability but morphological and pathological characterization[8], as well as population genetic analyses using molecular markers[9] have revealed high genetic diversity within the pathogen. Genetic diversity has been found to be high within a small spatial scale[9] and up to 74% of genetic variability was distributed within collection areas of approximately 1 m2 [10]. The source of this high level of genetic diversity is not known, although asexual recombination[11], spontaneous mutation[12] and sexual reproduction[10] have been proposed as possible mechanisms responsible for the high diversity.
The virulence structure of R. secalis populations may change over a relatively short period of time[13] and major resistance genes deployed in barley to control scald have frequently exhibited a finite life span due to the breakdown of resistance associated with selection for increased virulence in the pathogen[14,15]. Yitbarek et al.[1] reviewed work on barley disease surveys in Ethiopia and noted that, “more studies on variability within pathogen populations are needed to refine the breeding of barley for higher levels of disease resistance.” This is particularly true in the case of barley scald, where little is known concerning the variability in morphology, virulence and pathogenic specialization that exists within populations of R. secalis. The pathogenic variation of R. secalis presents a risk to the use of single gene resistance in barley cultivars. It is, therefore, important to identify and/or develop lines carrying as many different genes for resistance as possible in order to provide stable resistance against a broad spectrum of the fungal pathogen.
The aim of this study was to characterize isolates of R. secalis collected from barley in different agro-ecological zones of Ethiopia according to their morphological, cultural and pathogenic variation. Barley lines were also screened for scald resistance using single spore isolates representative of the resultant virulence phenotypes, in order to identify sources of resistance and to provide a basis for the future selection of isolates for use as inoculum in breeding programmes for scald resistance. The relative merits of different parameters to characterize severity levels of scald on individual barley genotypes in the high altitude, high rainfall zone of Bekoji, Ethiopia were also assessed. A preliminary report on this work has been published[16].
MATERIALS AND METHODS
Isolates of R. secalis: Naturally infected barley leaves were collected from various locations in Bale, Arsi and Shoa regions. Twenty-four single-spore isolates were obtained using the method of Jackson and Webster[17] and maintained in 2% Lima bean agar (LBA) at 17°C. To prevent pathogenicity changes in culture, there was minimal sub-culturing of isolates between isolation and pathogenicity. For short-term storage, spore suspensions were prepared in sterile distilled water, transferred to sterile culture tubes and stored at 5°C.
Morphological and cultural characteristics: Single colonies of each isolate were initiated by inverting a 3 mm mycelial plug from a 10-14-day-old culture onto each of four plates of LBA. The plates were incubated at 17°C in the dark for 10 days. From day 3, colony diameter was recorded every 3 days until no further growth was observed and used to compute the radial growth rates of the isolates[18]. On day 10, colony colour was described using a Mensell Soil Color Chart (1954). Colony topography (whether the surface of the colony was raised or not) and the nature of the spore masses (slimy or dry) were also described[7]. The sporulation capacity of isolates on LBA was evaluated using the method of Habgood[8].
Conidial suspensions in sterile distilled water were prepared from 10-day-old cultures of all isolates. Slide preparations of the conidial suspensions were subsequently made using cotton-blue lactophenol. Conidial size for each isolate was determined by measuring the length and width (at the broadest point) of 30 randomly chosen conidia using an eyepiece micrometer. The shape was determined by examination of 100 randomly selected conidia, which were then placed in one of two shape categories: beaked or non-beaked. The number of cells in each conidium was also noted.
Pathogenic variation: Pathogen variability of the 24 single spore isolates was evaluated on ten barley accessions, differing in resistant gene or gene combinations (Table 1), as described by Meles et al.[16]. Based on the differential reaction of the isolates and the host accessions, five pathotypes of R. secalis differing in pathogenic complexity (two with complex virulence capability, two with moderate virulence capability and one with simple virulence capability) were selected for use in glasshouse screening of 35 barley lines and a susceptible check cultivar. Glasshouse screening was conducted as described previously[16]. Cultivar-isolate combinations were repeated three times and disease ratings were made 13 and 20 days after inoculation.
| Table 1: | Barley differential varieties and their designated gene or gene combinations conferring resistance to R. secalis |
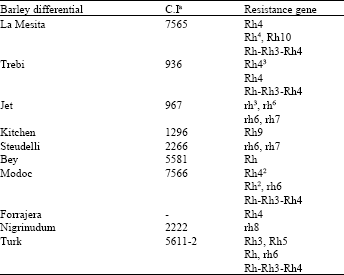 | |
| aAccession number of the Cereal Crops Research Branch, ARS, USDA, Beltsville, Maryland[3] | |
Reaction types were classified on a 0-4 scale. In this study, individual host genotypes commonly displayed two (occasionally three) different infection responses to a pathogen isolate across replicates. Where this occurred, the most severe reaction was taken as a score for that combination. Isolate-cultivar combinations with a reaction type of 0-2 were classified as resistant, while those with types 3 and 4 were considered as susceptible[16].
The relatedness of the 24 R. secalis isolates was determined by analyzing their infection responses on the host differentials using the Cluster Analysis module of the Statistica software v. 6.0[19]. The similarity of the 35 barley cultivars in response to the five selected R. secalis pathotypes was analyzed using the same module. Dissimilarity coefficients were generated using the percent disagreement (simple mismatch coefficient [SM]) calculated as SM = m/n where m = the number of mismatches between paired isolates or cultivars and n = the total number of differential hosts or pathotypes. Values of the dissimilarity coefficient (linkage distance) lie within the range of 0 to 1, where 0 indicates complete similarity and 1 represents complete dissimilarity. Phenograms were constructed based on these data sets using the unweighted pair-group method with arithmetic averaging (UPGMA) from the dissimilarity or distance matrices. Percent disagreement was used as distance measure because the data for the dimensions included in the analysis were categorical in nature.
Field screening of barley lines for resistance to scald: Thirty-five barley lines and a susceptible check cultivar were selected for this experiment. The experiment was conducted at Bekoji (50 km south of Asella), where scald infection was reported to be highly severe. Barley was grown at the experimental site in the previous season and there was enough left-over infested straw to serve as a ready source of inoculum. The field was laid out in a randomized complete block design with three replications. Barley seeds of each test cultivar were drilled on 4.8 m2 plots with a row to row distance of 20 cm. Plots were separated by two rows of the susceptible check cultivar ARDU 12-8°C, which also served as a spreader. Blocks were separated on all sides by four rows of the spreader. Within each randomized block, one plot of ARDU 12-8°C was also planted at intervals of five test cultivar plots. The fertilizers urea and diammonium phospate were each applied at the rate of 100 kg ha-1 at planting.
Disease assessment was made on the top three leaves 42, 48, 62, 75 and 89 days after planting (DAP). On each assessment date, 12 leaves were sampled at random from the middle rows of each plot and assessed for percent leaf area affected (PLAA), as the basis for estimating the area under the disease progress curve (AUDPC). PLAA was computed by comparison with a standard key for scoring blotch area[24]. AUDPC was calculated using the formula:
where xi refers to the per cent leaf area affected at ti (time, day) and n refers to the number of successive leaf area measurements. The apparent infection rate was also calculated using the following formula:
Assessment of number and size of complete lesions formed was also undertaken on the same leaves. A lesion was said to be complete when brown margins developed around the advancing edge of the lesion[22]. Lesion size and number of lesions were recorded at 48 and 75 DAP. Barley lines that had developed coalesced lesions at the time of assessment were excluded if the outline of the individual lesions wasn’t distinct, making it difficult to take lesion counts or measure lesion size.
At growth stage 11.1[23] the flag- and second-leaf (10 each) were randomly picked from each plot and assessed for PLAA using the standard key for blotch area[24]. The mean area under disease progress curve (AUDPC) for each barley line was regressed against grain yield. PLAA on the top two leaves at crop stage 11.1, lesion size and number and apparent infection rate were also tested for correlation with grain yield and 1000-grain weight and with each other. The level of scald infection on the test lines was also compared with the level of other associated diseases that were present in the field at the post-heading stage. The level of infection due to these diseases was recorded using the double digit scoring method[25], where the first digit refers to the relative height of the disease using the 0-9 Saari-Prescot scale and the second digit shows the disease severity on a 0-9 scale. Analysis of 1000-grain weight was done in random samples of grain taken from each plot. Twenty gram grain samples, dried at 35°C for 24 h, were used to determine 1000-grain weight[26]. Grain yield was also determined.
The MSTAT-C statistical package[27] was used for analysis of variance (ANOVA) tests in the laboratory, glasshouse and field experiments. When significant F values were obtained, treatment means were compared using Fisher’s least significant difference (F-LSD).
RESULTS AND DISCUSSION
Morphological and cultural characteristics: Marked and consistent differences were observed among the R. secalis isolates with respect to the colour and appearance of the fungal colonies. Moreover, significant differences existed among isolates in conidial characteristics, as well as in growth rate and spore production in culture (Table 2). More than a quarter of the isolates had spore counts of ≥ 3.0x106 spores ml-1 while half of the isolates had spore counts of ≤ 1.6x106 spores ml-1. Although growth of the isolates was slow, the strains showed significant differences in mycelial growth rate. When viewed from above, two thirds of the isolates had a colour category of 7/4, while the rest of the isolates had colour categories 7/6, 7/8, 8/3 and 8/4. But when viewed from below only colour categories 7/4, 7/6 and 8/4 were observed. Conidial length ranged from 5.0 to 24.2 μm and the mean length varied from 12.1 to 17.8 μm. Conidial width at the broadest point ranged from 1.5 to 7.1 μm while the mean width varied from 2.5 to 4.4 μm among the isolates. The most frequent conidial shape in about 2/3 of the isolates was the beaked type and conidia without beaks were observed in only nine of the isolates (Table 2). Isolate Holetta-32, however, had conidia with distinctly rounded ends. Though conidia typically had two cells, a considerable number of conidia with three cells were observed in isolates Chancho-16, Bekoji-28 and Meraro-13. The pathogenic complexity of isolates in this study appeared unrelated to both colony growth rate (r = -0.283, P=0.18) and the sporulation ability of the isolates. Jackson and Webster[17] found that the relative sporulation ability of isolates on potato dextrose agar was not necessarily indicative of their ability to sporulate on the host and to their pathogenic complexity. In agreement with the results of this study, they concluded that sporulation ability may be an attribute of isolates rather than races. Considerable variation was observed in morphological and cultural characteristics among isolates belonging to the same pathotype (Table 2). Since some of this variation is directly attributable to genic mutation[28], this may be an indication of the high adaptability of the pathogen population in Ethiopia[8].
Pathogenic variation and glasshouse and field response of barley cultivars to scald: The term “pathotype”[4] was used in this study to denote R. secalis isolates that differed in virulence on the barley differentials. Pathotype complexity referred to the ability of a pathotype to cause compatible reactions on several barley genotypes. A graphical presentation of pathotype diversity and variation in host response was obtained by generating dendrograms from the dissimilarity matrices based on the simple mismatch coefficient and the UPGMA clustering method (Fig. 1 and 2). Nineteen pathotypes were identified among the 24 R. secalis isolates based on the infection response of 10 host differentials carrying most of the known scald resistance genes. It is likely that a larger differential set, such as the 24 differentials described by Goodwin et al.[29] would have revealed more pathotypes in this study. Also, the ‘snap-shot’ nature of the present survey means that a larger sample size and an intensive hierarchical sampling strategy will likely reveal greater pathotype diversity. Knowledge of the pathogen population structure at experimental sites will be of great practical importance since evaluation of new barley lines and local and introduced cultivars, takes place at these locations[30]. Pathotype 16 showed the most complex virulence capability since it caused a compatible reaction in all the differential hosts. Interestingly, pathotypes 16, 7, 15 and 17 that had the top four most complex virulence phenotypes, comprised strains that were isolated from experiment stations of the state farms and the Institute of Agricultural Research. Also, all these strains were isolated from improved malting-barley lines. Although pathotype 6 was the most common virulence phenotype (representing 17% of the isolates), it displays a narrow virulence spectrum, being capable of causing susceptible reactions only on Steudelli and Kitchin. Such a pathotype perhaps represents a low level of virulence[31]; therefore, pathotype 6 should be supplemented with other pathotypes of a wide virulence spectrum for general resistance screening in Ethiopia.
The pathotypes were mostly represented by individual isolates, with the exception of pathotypes 6, 7 and 14 that were the only multi-isolate pathotypes (Fig. 1). Contrary to our expectation, none of the multi-isolate pathotypes contained isolates that originated from the same location.
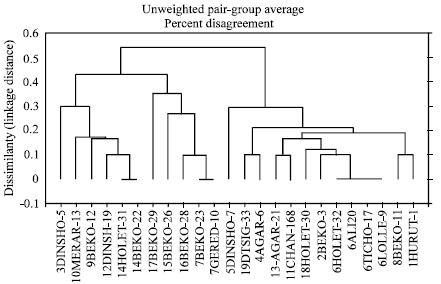 | |
| Fig. 1: | Dendrogram showing pathotype diversity among 24 R. secalis isolates from barley in Ethiopia following virulence phenotyping using ten host differentials. The isolates are designated by a code showing the pathotype number followed by the abbreviated isolate designation (Table 2) |
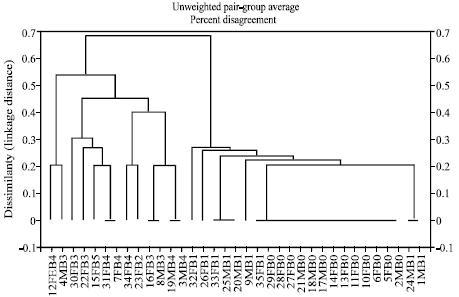 | |
| Fig. 2: | Dendrogram showing relationships among 35 barley lines in response to five selected R. secalis pathotypes following glasshouse screening. The lines are designated by a code showing the cultivar serial number (Table 3) followed by the barley type (food barley, FB or malting barley, MB) and the number of pathotypes to which the cultivar was resistant. |
For instance, pathotype 6 that was the most common pathotype was represented by four isolates collected from different locations in Arsi, Bale and Shoa. Pathogenic differences have been reported among single spore cultures of R. secalis derived from the same geographical location, plant, lesion, or even the same parent isolate[32]. Although no comparisons were made in this study below the sampling scale of a location, the high pathogenic variability within sites such as Bekoji, Dinsho and Holetta clearly suggests a wide virulence spectrum of R. secalis across locations in Ethiopia.
The susceptible check cultivar ARDU 12-8C was susceptible to all the five pathotypes (3, 5, 7, 11 and 16) selected for screening of the 35 Ethiopian barley lines in the glasshouse. The most complex pathotypes 16 and 7 caused susceptible reactions in only 21 and 22 of the 35 barley lines, respectively, in the glasshouse. In contrast, the least complex pathotype 6 caused susceptible reactions in 25 barley lines, while 24 lines were susceptible to pathotype 3 that was of intermediate virulence capability. Cluster analysis of host response in the glasshouse revealed two major clusters, which clearly separated cultivars that were resistant to zero and one pathotype from those that showed resistance to more than one pathotype (Fig. 2).
| Table 2: | Morphological and cultural characteristics of 24 isolates of R. secalis used in this study |
 | |
| aColour hue 5YR 7/4, 8/4 and 8/3 = pink; hue 5YR 7/6 and 7/8 = reddish yellow colony, bColony appearance after 18 days cMean linear growth rate of colony (mm day-1), dIntensity of sporulation in culture (spores ml-1), eLength, width and shape of conidia; B = beaked conidia, NB = non-beaked conidia | |
In the present study, the reaction of the barley lines in the glasshouse differed from that in the field. Seventeen cultivars that were classified in the field as resistant were found to be resistant to only one of the five pathotypes with which they were screened in the glasshouse. The remaining lines were resistant to 2, 3 or 4 pathotypes, with HB-117 alone being resistant to all the five pathotypes. Conversely, barley lines such as KLDN 54/85 and KLDN 209/85 classified as highly susceptible in the field, were able to resist even the most complex pathotype (pathotype 16) in the glasshouse. These findings clearly demonstrate that the cultivar reactions in the glasshouse seedling test were not always expressed in the field. Jackson and Webster[17] and Webster et al.[33] made similar findings and considered that the pathotype complexity observed on differential hosts may not be relevant to host-pathogen interactions involving other cultivars/landraces. The inability of the more complex pathotypes to infect more barley lines may be explained, at least in part, by the possession by these cultivars of novel resistance gene(s) or gene combinations not present in the differential cultivars.
The lack of a well-defined set of differential cultivars for pathotyping of R. secalis makes direct comparison of our results with the results of other pathotyping studies quite difficult[34]. To stem the problems inherent in the use of differential cultivars with widely differing genetic backgrounds and having two or more known or assumed resistance genes, Bjørnstad et al.[34] recently suggested a more appropriate terminology of scald resistance genes and proposed a set of near isogenic lines (NILs) derived from the current differentials for use as a standard. Their NILs are yet to be adopted as a new set of standard differentials and were not available for this study.
AUDPC was used as a measure of the resistance or susceptibility of the barley lines at Bekoji. Significant differences (P≤0.05) were observed in AUDPC, r and PLAA among the test barley lines. Considerable variation was also observed in grain yield and 1000-grain weight (Table 3). Most of the lines with AUDPC values below 133.6 also had grain yield of 66.2 to 166.7% over the yield of the susceptible check; however, a few barley lines with low AUDPC such as HB-116 and HB-68 gave yields that were similar to that of the control. Grain yield and 1000-grain weight were negatively and significantly correlated with both AUDPC and PLAA, but the correlation with apparent infection rate was weak and non-significant (Table 4). Assessment of foliar diseases at the post-heading stage showed that net blotch (Pyrenophora teres Dreschl) was the most important disease associated with scald at Bekoji. There was a negative and significant correlation between scald and net-blotch (r = -0.53, P≤0.05).
| Table 3: | Mean percent leaf area damage on flag and 2nd leaf (PLAA), area under the disease progress curve (AUDPC), double digit score, apparent infection rate (r), grain yield and 1000 grain weight of 35 barley lines at Bekoji |
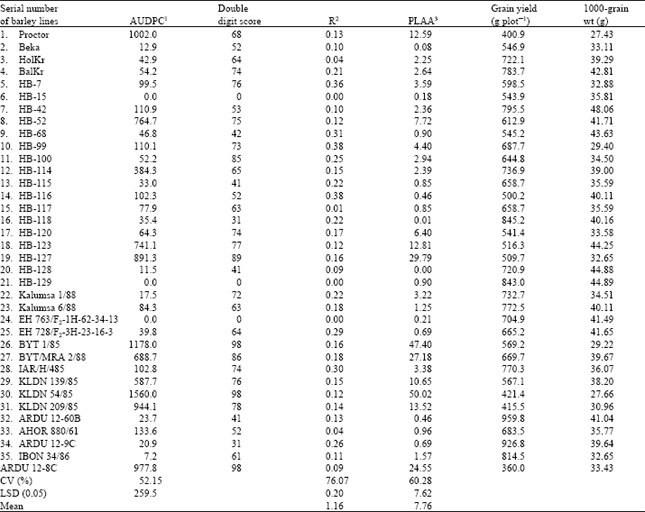 | |
| 1Area under the disease progress curve, 2Apparent infection rate, slope of the logit disease progress curve 3Percentage leaf area affected (only top two leaves) | |
| Table 4: | Correlation analysis between grain yield, 1000 grain weight, area under the disease progress curve (AUDPC), plant leaf area affected (PLAA) and the apparent infection rate “r” |
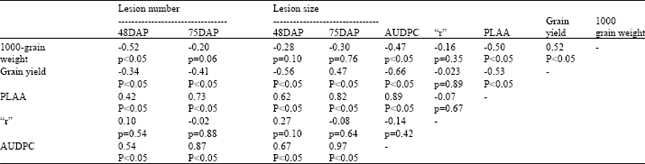 | |
Although somewhat demanding with respect to data collection, computation and interpretation, AUDPC is often favoured as the best parameter to characterize levels of partial resistance within a particular environment[35]. AUDPC revealed marked differences in response to scald among the barley lines compared to r, which agrees with the findings of Wilcoxon et al.[36] and Parlevliet[37]. Parlevliet[37] observed that r wasn’t useful in differentiating resistant from susceptible cultivars while distinct and consistent differences were observed in AUDPC. A very close relationship was observed between the double-digit score (Table 3)[25] and the AUDPC, indicating that this simple yet robust and biologically meaningful parameter may be used to substitute AUDPC in large multi-environment field trials as proposed by Pinnschmidt[35]. Breeding lines such as HB-129 that had AUDPC values of zero possibly carry major resistance gene(s) and may not be promising candidates for use as donors of quantitative resistance to scald in the breeding program. Similarly, lines such as BYT 1/85 and KLDN-54/85 with AUDPC of over 1000 probably possess a very low level of scald resistance. On the other hand, barley lines such as HB-100 possessing resistance to all five pathotypes in the glasshouse and having low AUDPC, r and PLAA values and with high yields appear promising as donors of both quantitative resistance and yield. The finding that some barley lines that were highly resistant to scald gave low yields, may be explained, at least in part, by their susceptibility to net blotch[6]. James et al.[38] and Khan and Crosbie[39] reported that yield loss due to scald was accounted for mainly by a loss in grain weight. The significant positive correlation between grain yield and 1000-grain weight observed in this study supports their finding; however, some lines such as HB-123 and HB-68 had high grain weight but relatively low yield. Low yield in these cases may have been brought about by reduced grain number per ear and/or reduction in the number of tillers per plant[38].
The vast majority of farmers in Ethiopia are still growing local varieties and have little means of increasing output. Although most of the preferred local varieties are susceptible to foliar diseases, particularly scald[30], the resistance against scald remains low. Ethiopia is one of the centres of diversity of barley and is thus a presumed centre of diversity of its pathogen R. secalis. Since the pathogen is expected to show a high level of genetic diversity in countries like Ethiopia[40], host resistance could potentially form the basis of barley scald management strategies. This first study has shown that high yielding barley lines such as HB-100 with resistance to all five pathotypes tested and exhibiting low AUDPC, r and PLAA values in the field are promising as donors of quantitative resistance genes in scald resistance breeding in the high altitude zone of Bekoji. Clearly, pathotypes with complex virulence capability such as pathotypes 16, 7, 15 and 17 will be good candidates for initial screening for scald resistance since they caused a compatible reaction in most of the differential hosts. Since pathotype structure may differ across locations, further research is now warranted to test promising lines such as HB-100 for genotype x environment interaction. There is also need to use molecular markers and a larger sample size to gain more insight into the population biology of R. secalis in Ethiopia.
REFERENCES
- Goodwin, S.B., R.W. Allard, S.A. Hardy and R.K. Webster, 1992. Hierarchical structure of pathogenic variation among Rhynchosporium secalis populations in Idaho and Oregon. Can. J. Bot., 70: 811-817.
CrossRef - McDonald, B.A., J. Zhan and J.J. Burdon, 1999. Genetic structure of Rhynchosporium secalis in Australia. Phytopathology, 89: 639-645.
Direct Link - Newton, A.C., 1989. Somatic recombination in Rhynchosporium secalis Plant Pathol., 38: 71-74.
CrossRef - Goodwin, S.B., R.K. Webster and R.W. Allard, 1994. Evidence for mutation and migration as sources of genetic variation in populations of Rhynchosporium secalis. Phytopathology, 84: 1047-1053.
Direct Link - Jackson, L.F., A.L. Kahler, R.K. Webster and R.W. Allard, 1978. Conservation of scald resistance in barley composite cross populations. Phytopathology, 68: 645-650.
CrossRef - Meles, K., M. Hulluka and M.L. Deadman, 2000. Pathogenic variation in Rhynchosporium secalis on barley in Ethiopia. Sultan Qaboos Univ., 5: 75-78.
Direct Link - Jackson, L.F. and R.K. Webster, 1976. The dynamics of a controlled population of Rhynchosporium secalis, changes in race composition and frequencies. Phytopathology, 66: 726-728.
CrossRef - Baxter, A.P., G.C.A. van der Westhuizen and A. Eicker, 1983. Morphology and taxonomy of South African isolates of Colletotrichum. S. Afr. J. Bot., 2: 259-289.
CrossRefDirect Link - Shtienberg, D., S.N. Bergeron, A.G. Nicholson, W.E. Fry and E.E. Ewing, 1990. Development and evaluation of a general model for yield loss assessment in potatoes. Phytopathology, 80: 466-472.
Direct Link - Williams, R.J. and H. Owen, 1975. Susceptibility of barley cultivars to leaf blotch and aggressiveness of Rhynchosporium secalis races Trans. Br. Mycol. Soc., 65: 109-114.
CrossRef - Skon, J.P. and V. Haahr, 1987. Field screening for response to barley net-blotch. Ann. Applied Biol., 111: 617-627.
CrossRef - McDermott, J.M., B.A. McDonald, R.W. Allard and R.K. Webster, 1989. Genetic variability for pathogenicity, isozyme, ribosomal DNA and colony color variants in populations of Rhynchosporium secalis. Genetics, 122: 561-565.
PubMedDirect Link - Goodwin, S.B., R.W. Allard and R.K. Webster, 1990. A nomenclature for Rhynchosporium secalis pathotypes. Phytopathology, 80: 1330-1336.
Direct Link - Xi, K., J.H. Turkington and C. Bos, 2002. Pathogenic variation of Rhynchosporium secalis in Alberta. J. Plant Pathol., 24: 176-183.
Direct Link - Webster, R.K., L.F. Jackson and C.W. Schaller, 1980. Sources of resistance in barley to Rhynchosporium secalis. J. Plant Dis., 64: 88-90.
Direct Link - Bjornstad, A., V. Patil, A. Tekauz, A.G. Maroy and H. Skinnes et al., 2002. Resistance to scald (Rhynchosporium secalis) in barley (Hordeum vulgare) studied by near-isogenic lines: I. Markers and differential isolates. J. Phytopathol., 92: 710-720.
Direct Link - Wilcoxson, R.D., B. Skovmand and A.H. Atif, 1975. Evaluation of wheat cultivars for ability to retard development of stem rust. J. Ann. Appl. Biol., 80: 275-281.
CrossRefDirect Link - James, D.G., E.E. Jenkins and J.L. Jemmet, 1968. The relationship between leaf blotch caused by Rhynchosporium secalis and losses in grain yield of barley. Ann. Applied Biol., 62: 273-288.
CrossRefDirect Link - Khan, T.N. and G.B. Crosbie, 1988. Effect of scald (Rhynchosporium secalis (Oud.) J. Davis) infection on some quality characteristics of barley. Australian. Aust. J. Exp. Agric., 28: 783-785.
Direct Link - McDonald, B.A. and C. Linde, 2002. Pathogen population genetics, evolutionary potential and durable resistance. J. Annu. Rev. Phytopathol., 40: 349-379.
PubMed