
Elsayed K. Bakhiet
Department of Botany and Microbiology, Faculty of Sciences, Al-Azhar University, Assiut 71524, Egypt
Hosni A.M. Hussien
Department of Botany and Microbiology, Faculty of Sciences, Al-Azhar University, Assiut 71524, Egypt
LiveDNA: 20.38291
Mohamed Elshehaby
Department of Zoology, Faculty of Sciences, Al-Azhar University, Assiut 71524, Egypt
Pakistan Journal of Biological Sciences
Year: 2022 | Volume: 25 | Issue: 10 | Page No.: 875-884







ABSTRACT
Background and Objective: Bacterial and fungal infections are major public health problems. Emerging of drug-resistant microbial strains urges the need for the development of alternative untraditional antimicrobial agents. Bee venom is a rich source of secondary metabolites and antimicrobial agents. In this study, the antimicrobial and antifungal potential of Apis mellifera BV (AmBV) against some medically important bacterial and fungal pathogens was investigated. Materials and Methods: Broth microdilution method and Colony Forming Unit (CFU) assay were used to screen the antibacterial potential of AmBV. Similarly, the antifungal activity of AmBV was evaluated using the agar-well diffusion assay. Moreover, the minimum inhibitory concentration (MIC) values of AmBV against tested microorganisms were determined. Results: AmBV significantly inhibited bacterial and fungal growth. The MIC values of AmBV were 15.625, 31.25, 7.8, 7.8 μg mL–1 against Escherichia coli ATCC 8739, Staphylococcus aureus ATCC 6538P, Serratia marcescens AUH 98 and Streptococcus mutans ATCC 25175, respectively. Similarly, AmBV at concentrations of 300 and 600 μg mL–1 significantly inhibited the growth of Aspergillus niger ATCC 16404, Alternaria alternata MLBM09, Fusarium oxysporum MLBM212 and Aspergillus flavus. Conclusion: These results indicated that AmBV could be used in future preclinical and clinical studies to develop cost-effective and efficient antibacterial and antifungal agents. Moreover, this study presents AmBV as an efficient alternative antimicrobial agent against medically important pathogens.
PDF Abstract XML References Citation
Copyright: © 2022. This is an open access article distributed under the terms of the creative commons attribution License, which permits unrestricted use, distribution and reproduction in any medium, provided the original author and source are credited.
How to cite this article
Elsayed K. Bakhiet, Hosni A.M. Hussien and Mohamed Elshehaby, 2022. Apis mellifera Venom Inhibits Bacterial and Fungal Pathogens in vitro. Pakistan Journal of Biological Sciences, 25: 875-884.
DOI: 10.3923/pjbs.2022.875.884
URL: https://scialert.net/abstract/?doi=pjbs.2022.875.884
DOI: 10.3923/pjbs.2022.875.884
URL: https://scialert.net/abstract/?doi=pjbs.2022.875.884
INTRODUCTION
Bee’s venom (BV) has been used in traditional medicine applications to treat a variety of diseases1. There are many pharmacologically and biologically active components in BV, including there are many pharmacologically and biologically active components in BV including melittin, adolapin, apamin, hyaluronidase, phospholipase A2, histamine, epinephrine, lipids, carbohydrates and minerals2,3. The BV is released by the poison glands of honeybee workers as a defence mechanism4.
Natural products such as BV are bioactive compound-rich products and have been shown to mediate a wide range of effects against several diseases5,6. Although BV is poisonous to bee predators, it has been developed as a medicinal tool by humans through time. The use of BV for therapeutic purposes dates back to Ancient Egypt (4000 BC), Hippocrates, Aristotle and Galen used it during the Greek and Roman periods7,8. Inflammatory disorders such as rheumatoid arthritis, tendinitis, fibrosis, lupus and multiple sclerosis were treated with BV in traditional Chinese medicine and other historical traditions8,9. Melittin is the main active polypeptide in BV and has anti-cancer, anti-inflammatory, antibacterial and antiviral properties10,11. In their study, Yu et al.12 demonstrated that BV has potent antifungal properties against Trichophyton mentagrophytes and Trichophyton rubrum, which are far more potent than fluconazole, a commercial antifungal used to treat superficial and systemic fungal infections.
Multidrug-resistant bacteria (MDRB) like Escherichia coli (E. coli ) are becoming more common globally, this in part owing to the expansion of bacterial mobile resistant genetic components like plasmids13. Similarly, MDRB strains of S. marcescens cause a wide spectrum of clinical features including pneumonia, meningitis, conjunctivitis, sepsis, urinary tract infections and surgical wound infections14. S. aureus is one of the major opportunistic human pathogens. S. aureus evades the immune system and causes a variety of human illnesses ranging from minor skin irritations to life-threatening sepsis15,16. Streptococcus mutans (S. mutans) colonize the oral cavity and are responsible for dental caries and periodontal diseases. The antimicrobial resistance of many bacterial and fungal strains has limited the efficacy of available commercial antimicrobial agents. Thus, new untraditional antimicrobial agents are highly in need to control infection of MDRB. The current study aimed to evaluate the antimicrobial activity of Apis mellifera BV (AmBV) against some important bacterial and fungal pathogens.
MATERIALS AND METHODS
Study area: This study was conducted in 2021 at the Botany and Microbiology Department, Faculty of Science, Al-Azhar University, Assiut Governorate, Egypt.
Microbial strains: For this study, the American Type Culture Collection (ATCC) reference strains (Streptococcus mutans ATCC 25175 (S. mutans ATCC 25175), Staphylococcus aureus ATCC 6538P (S. aureus ATCC 6538P), Escherichia coli ATCC 8739 (E. coli ATCC8739) and Aspergillus niger ATCC 16404 (A. niger ATCC 16404) and clinical isolates (Serratia marcescens AUH 98 (S. marcescens AUH 98), Alternaria alternata MLBM09 (A. alternata MLBM09), Fusarium oxysporum MLBM212 (F. oxysporum MLBM212) and Aspergillus flavus (A. flavus)) were used17. Bacterial strains were grown on Mueller Hinton Broth (MHB) at 37°C, while the fungal strains were grown in potato dextrose agar (PDA) medium and incubated at 28°C. Bacterial suspensions were freshly prepared before each experiment and adjusted to the turbidity of 0.5 McFarland standards (OD630 ~ 0.08 in MHB). Bacterial suspensions were used within 30 min of preparation.
Collection of Apis mellifera Bee Venom (AmBV): The venom of Apis mellifera forager honeybees descended from naturally mated queens was used to study its antimicrobial activity as previously described by Surendra et al.18. These bees were derived from a colony pool and collected from the Department of Bees, The Institute of Plant Protection, the Center for Agriculture and the Ministry of Agriculture Egypt. They were rendered immobile by rapid freezing at -20°C. The stinging apparatuses of honeybees were dissected at 4°C and the venom reservoirs were extracted and stored at -20°C till used in subsequent assays. Venom sacs were resuspended in Milli-Q water and whole bee venom (WBV) was extracted by disrupting the reservoir using a glass rod under rapid defrosting and light pressure. Extracted venom samples were centrifuged at 10,000 g for 5 minutes at 4°C and the supernatants were used as protein and enzyme sources before being lyophilized. Lyophilized Bee venom was dissolved in Milli-Q water and filtered through a 0.22 m syringe filter before using it in further experiments.
Determination of the antibacterial activity of AmBV
Broth microdilution method: Serially diluted AmBV was transferred into wells of 96-well plate in 90 μL volume. The final concentrations of AmBV in wells were 7.6-4000 μg mL–1. Next, 10 μL of bacterial suspension was added to each well. The plate was incubated at 37°C for 24 hrs before reading the optical density (OD) at 630 nm using a microplate reader19. Untreated (UT) bacterial suspension was used as a growth control.
Colony Forming Unit (CFU) assay: Antibacterial activity of AmBV was assessed by counting Colony Forming Units (CFUs)20-22. Briefly, 1 mL of serially diluted AmBV (250 to 4000 μg mL–1) was inoculated with 10 μL of bacterial suspension and incubated at 37°C for 6 hrs. Next, 10-fold serial dilutions were prepared from each treatment and 10 to 100 μL of each dilution was streaked on Muller-Hinton agar (MHA) plates. Bacterial colonies were counted after overnight incubation at 37°C. The MHA plates inoculated with untreated bacteria were used as growth control.
Determination of minimum inhibitory concentration (MIC): A broth microdilution susceptibility test was used to detect MIC as previously described20,23-25. Briefly, two-fold serially diluted AmBV was transferred into wells of 96-well plate in 90 μL volume. The serially diluted concentrations of AmBV that used to determine MIC were 4000, 2000, 1000, 500, 250, 250, 125, 62.5, 31.25, 15.625 and 7.8 μg mL–1. Next, 10 μL of bacterial suspension was added to each well and the plate was incubated at 37°C for 24 hrs before reading the optical density (OD) at 630 nm using a microplate reader. The MIC value was determined as the minimum concentration of AmBV that significantly decrease the OD630 value with no visible bacterial growth.
Antifungal activities of AmBV: The antifungal activities of AmBV were evaluated through the agar-well diffusion assay as previously described26-28. The solution has been tested at different concentrations (600, 300 and 150 μg mL–1).
Statistical analysis: The growth reduction was calculated relative to the growth control. Data were shown as Means±Standard Deviation (SD) of at least three independent experiments. Comparisons between various treatments were performed by t-test and one-way ANOVA. The p<0.05 was regarded as statistically significant.
RESULTS
Antibacterial activity of AmBV
Broth microdilution methods: Results from Broth microdilution methods demonstrated that AmBV significantly inhibits the growth of the tested bacterial strains in a concentration-dependent manner. AmBV was significantly inhibited the growth of E. coli ATCC8739 at concentration of 15.625 μg mL–1 (p<0.05) in Fig. 1a.
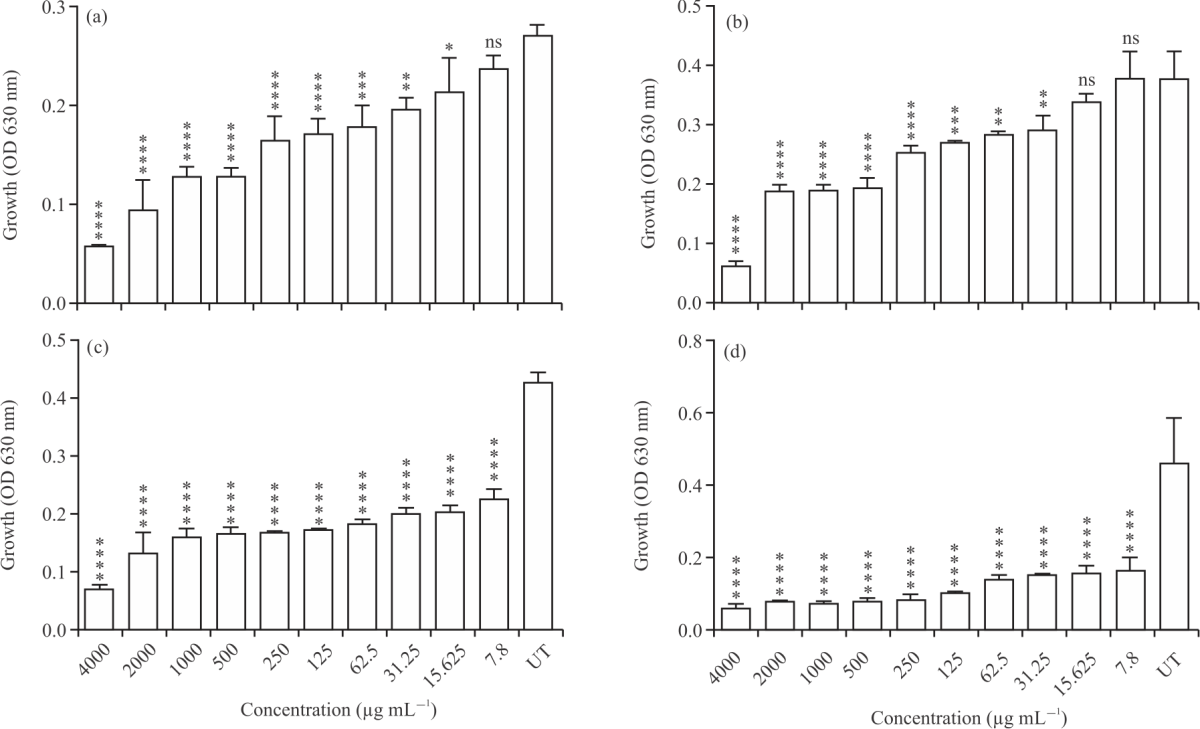 |
| Fig. 1(a-d): | Antibacterial activity of different concentrations of AmBV against bacterial species, (a) E. coli ATCC 8739, (b) S. aureus ATCC 6538P, (c) S. marcescens AUH 98 and (d) S. mutans ATCC 25175 Results are shown as Means±SD of three independent experiments,* p<0.05, **p<0.01, ***p<0.001 and ****p<0.0001 vs. untreated control |
| Table 1: | Antibacterial activity of AmBV | |||
| Concentration (μg mL–1) | E. coli ATCC 8739 | S. marcescens AUH 98 | S. aureus ATCC 6538P | S. mutans ATCC |
| UT | 0.2707±0.009815 | 0.4250±0.01803 | 0.3770±0.04480 | 0.4573±0.1249 |
| 4000 | 0.0580±0.002646 | 0.06767±0.008963 | 0.0630±0.006245 | 0.0620±0.008000 |
| 2000 | 0.09433±0.02977 | 0.1303±0.03661 | 0.1877±0.01102 | 0.07967±0.007638 |
| 1000 | 0.1283±0.008963 | 0.1583±0.01557 | 0.1890±0.008718 | 0.0774±0.006773 |
| 500 | 0.1287±0.007767 | 0.1633±0.01222 | 0.1940±0.01652 | 0.08067±0.008505 |
| 250 | 0.1647±0.02346 | 0.1670±0.001000 | 0.2537±0.01002 | 0.08633±0.01222 |
| 125 | 0.1710±0.01442 | 0.1693±0.002517 | 0.2700±0.004359 | 0.1033±0.007024 |
| 62.5 | 0.1787±0.02065 | 0.1800±0.009165 | 0.2830±0.005000 | 0.1407±0.01172 |
| 31.25 | 0.1967±0.01007 | 0.1990±0.0110 | 0.2910±0.02330 | 0.1523±0.006506 |
| 15.625 | 0.2140±0.03318 | 0.2017±0.01193 | 0.3387±0.01250 | 0.1583±0.01815 |
| 7.8 | 0.2377±0.01185 | 0.2233±0.01893 | 0.3770±0.04480 | 0.1663±0.03213 |
The growth of E. coli ATCC8739 was further inhibited at AmBV concentrations of 31.25, 62.50 and >125 μg mL–1 (p<0.05) (Fig. 1a). The OD value of E. coli ATCC 8739 growth was decreased from 0.2707±0.009815 at untreated control to 0.2140±0.03318, 0.1967±0.01007, 0.1787±0.02065, 0.1710 ±0.01442, 0.1647±0.02346, 0.1287±0.007767, 0.1283±0.008963, 0.09433±0.02977, 0.0580±0.002646 when treated with AmBV concentrations of 15.625, 31.25, 62.5, 125, 250, 500, 1000, 2000 and 4000 μg mL–1, respectively in Fig. 1a and Table 1. Similarly, the growth of S. aureus ATCC 6538P was significantly inhibited by AmBV at concentrations of 31.25, 62.50, 125 and >250 μg mL–1 (p<0.05) in Fig. 1b. The OD value of bacterial growth was decreased from 0.3770±0.04480 in untreated culture to 0.2910±0.02330, 0.2830±0.005000, 0.2700±0.004359, 0.2537±0.01002, 0.1940±0.01652, 0.1890±0.008718, 0.1877±0.01102, 0.0630±0.006245 when treated with AmBV concentrations of 31.25, 62.5, 125, 250, 500, 1000, 2000 and 4000 μg mL–1, respectively in Fig.1b and Table 1. Interestingly, the growth of both S. marcescens AUH 98 in Fig. 1c and S. mutans ATCC 25175 in Fig. 1d were significantly inhibited at all used AmBV concentrations (4000-7.8 μg mL–1 (p<0.05)). The OD value of the growth of S. marcescens AUH 98 was decreased from 0.4250±0.01803 at untreated control to 0.2233±0.01893, 0.2017±0.01193, 0.1990±0.0110, 0.1800±0.009165, 0.1693±0.002517, 0.1670±0.001000, 0.1633±0.01222, 0.1583±0.01557, 0.1303±0.03661, 0.06767±0.008963 at AmBV concentrations of 7.8, 15.625, 31.25, 62.5, 125, 250, 500, 1000, 2000 and 4000 μg mL–1, respectively (Fig.1c and Table 1). While, the OD value in case of S. mutans ATCC 25175 was decreased from 0.4573±0.1249 in untreated control to 0.1663 ±0.03213, 0.1583±0.01815, 0.1523±0.006506, 0.1407±0.01172, 0.1033±0.007024, 0.08633±0.01222, 0.08067±0.008505, 0.0774±0.006773, 0.07967±0.007638, 0.0620 ±0.008000 at AmBV concentrations of 7.8, 15.625, 31.25, 62.5, 125, 250, 500, 1000, 2000 and 4000 μg mL–1, respectively in Fig.1d and Table 1. Bacterial growth inhibition by AmBV was concentration-dependent. In case of E. coli ATCC 8739, the percentages of growth inhibition were 78.56, 65.26, 52.58, 52.44, 39.26, 36.86, 34.04, 27.35, 21.13 and 16.02% at 4000, 2000, 1000, 500, 250, 125, 62.5, 31.25, 15.625 and 7.8 μg mL–1 of AmBV, respectively in Fig. 2a. For S. aureus ATCC 6538P, the growth inhibition was 82.14, 46.35, 45.90, 44.58, 27.74, 22.94, 19.12, 17.2 and 3.216% at 4000, 2000, 1000, 500, 250, 125, 62.5, 31.25 and 15.625 μg mL–1 of AmBV, respectively in Fig. 2b. Interestingly, all the used concentrations of AmBV inhibited the growth of S. marcescens AUH 98 in Fig. 2c. and S. mutans ATCC 25175 in Fig. 2d by more than >47.4 and >60.55%, respectively.
CFU assay: AmBV significantly reduced the CFUs of the treated bacteria. At concentrations of 250-4000 μg mL–1 of AmBV, CFUs of E. coli ATCC 8739 were significantly decreased (p<0.0001) compared to untreated control in Fig. 3a. The CFUs of E. coli ATCC 8739 were 453 CFU mL–1 at untreated control while at AmBV concentrations of 250, 500, 1000, 2000 and 4000 μg mL–1, CFUs of E. coli ATCC 8739 were decreased to 353, 308, 219, 165 and 145 CFU mL–1, respectively (Fig. 3a). Similarly, AmBV concentrations of 250-4000 μg mL–1 were significantly decreased the growth of S. aureus ATCC 6538P (p<0.0001) in Fig. 3b. The CFUs of S. aureus ATCC 6538P were decreased from 505 at untreated control to 360-265, 155, 48 and 30 CFU mL–1 at AmBV concentrations of 250, 500, 1000, 2000 and 4000 μg mL–1, respectively (Fig. 3b). The CFUs of S. marcescens AUH 98 were significantly reduced at AmBV concentrations of 250 μg mL–1 (510 CFU mL–1) (p<0.0001), 500 μg mL–1 (389 CFU mL–1) (p<0.0001), 1000 μg mL–1 (362 CFU mL–1) (p<0.0001), 2000 μg mL–1 (306 CFU mL–1) (p<0.0001) and 4000 μg mL–1 (205 CFU mL–1) (p<0.0001) compared to untreated control in Fig. 3c. Similarly, the CFUs of S. mutans ATCC 25175 were significantly reduced from 750 CFU mL–1 at untreated control to 535, 260, 210, 163 and 19 CFU mL–1 when treated with AmBV concentrations of 250, 500, 1000, 2000 and 4000 μg mL–1, respectively in Fig. 3d.
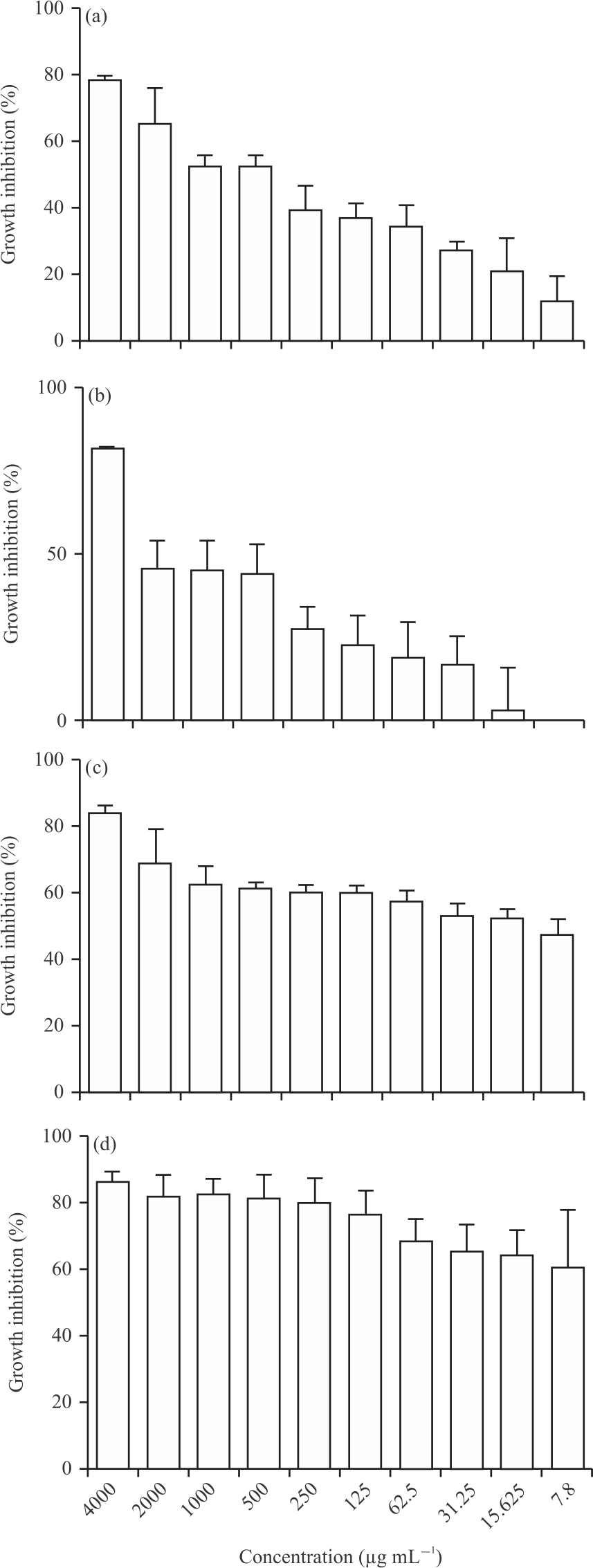 |
| Fig. 2(a-d): | Percentage of bacterial inhibition by different concentrations of AmBV against bacterial species (a) E. coli ATCC 8739, (b) S. aureus ATCC 6538P, (c) S. marcescens AUH 98 and (d) S. mutans ATCC 25175 Results are shown as Means±SD of three independent experiments |
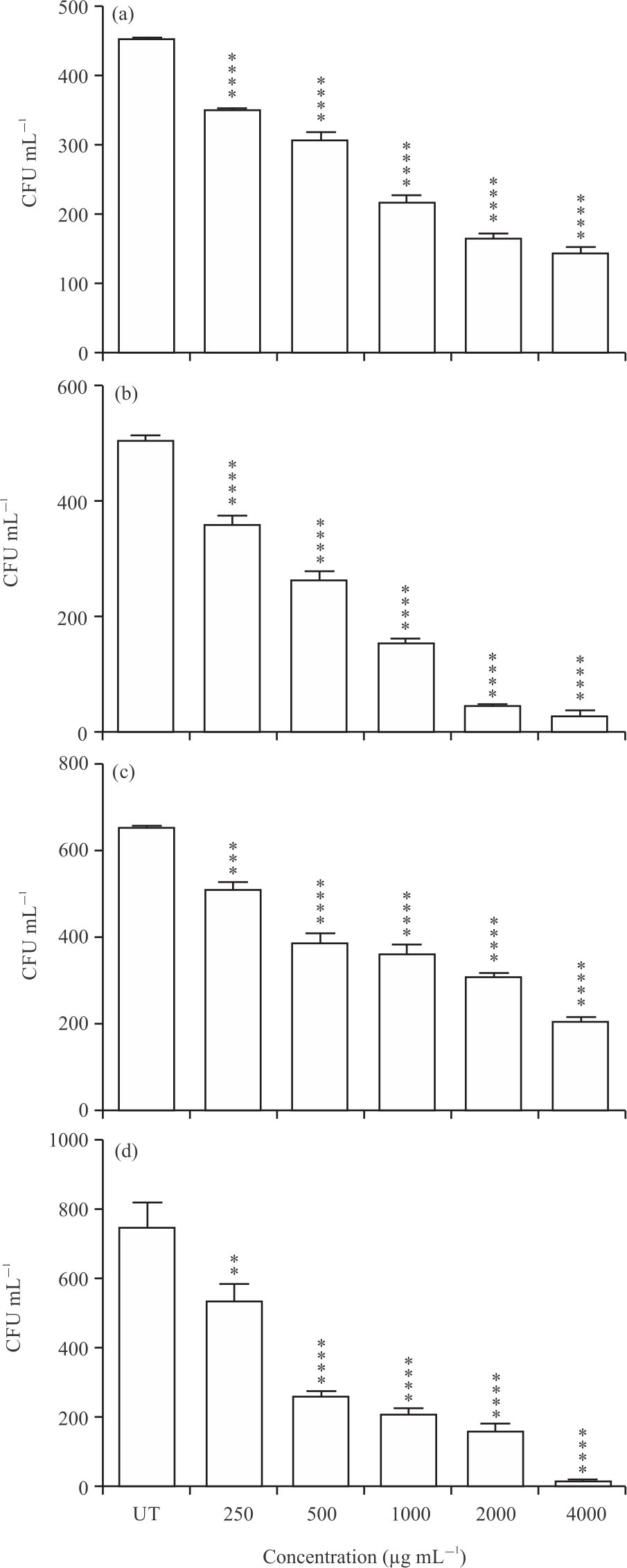 |
| Fig. 3(a-d): | Effect of different concentrations of AmBV on CFUs of bacterial species, (a) E. coli ATCC 8739, (b) S. aureus ATCC 6538P, (c) S. marcescens AUH 98 and (d) S. mutans ATCC 25175 Results are shown as Means±SD of three independent experiments,* p<0.05, **p<0.01, ***p<0.001 and ****p<0.0001 vs. untreated control |
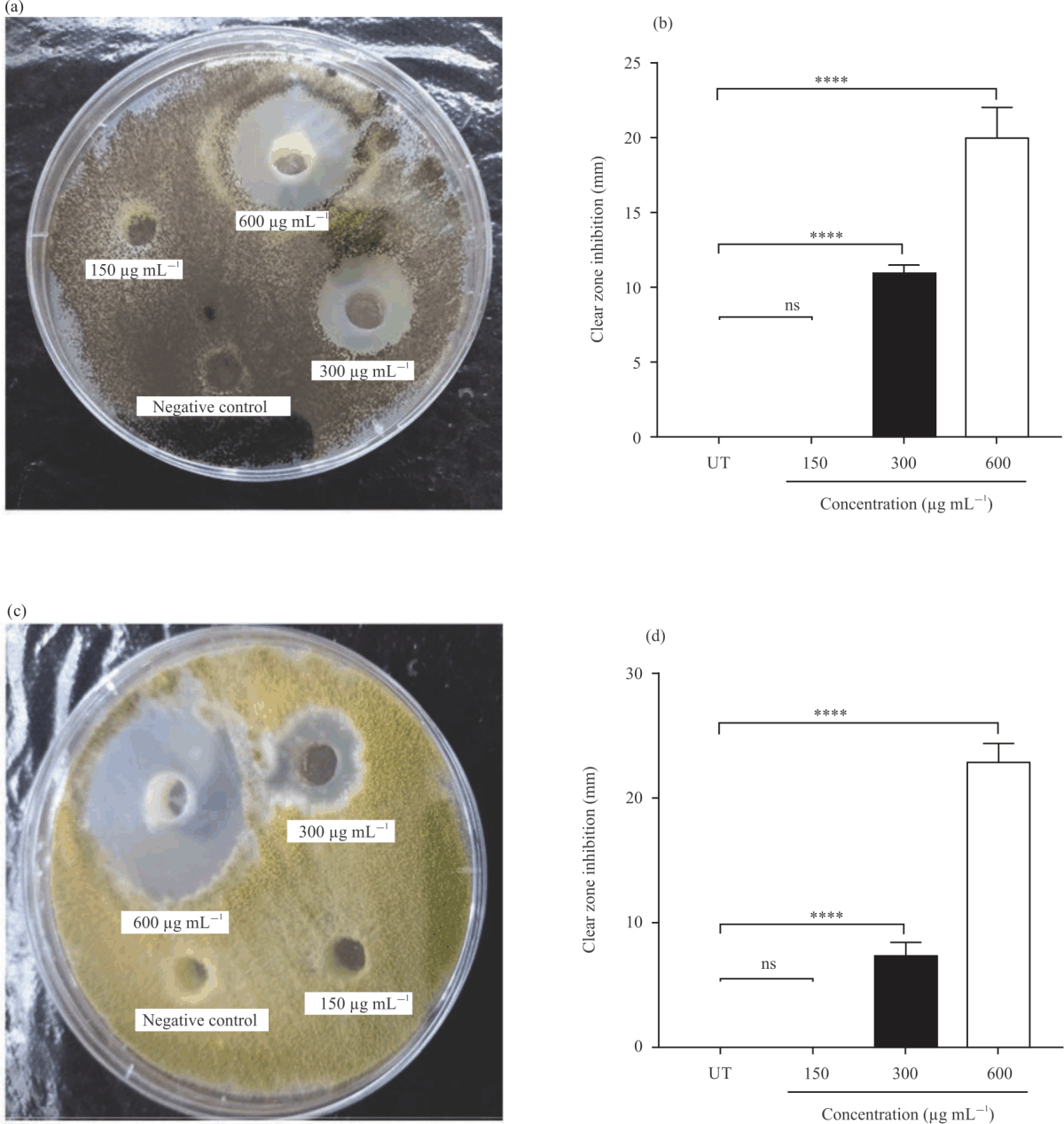 |
| Fig. 4(a-d): | Antifungal activity of different concentrations of AmBV against bacterial species, (a) Representative photographs of antifungal assays of A. niger ATCC 16404, (b) A. niger ATCC 16404 diameter of clear zone inhibition in millimeter (mm), (c) Representative photographs of antifungal assays of A. flavus and (d) A. flavus diameter of clear zone inhibition in millimeter (mm) Results are shown as Means ±SD of three independent experiments and p-value: ****p<0.0001 vs. negative control |
MIC values of AmBV: There were variations in MIC values of AmBV between treated bacterial strains. MIC values of AmBV were 15.625 μg mL–1 (OD: 0.2140±0.03318), 31.25 μg mL–1 (OD: 0.2910±0.02330), 7.8 μg mL–1 (OD: 0.2233±0.01893) and 7.8 μg mL–1 (OD: 0.1663±0.03213) against E. coli ATCC 8739, S. aureus ATCC 6538P, S. marcescens AUH 98 and S. mutans ATCC 25175, respectively in Table 2.
Antifungal activity of AmBV: AmBV exhibited a remarkable antifungal activity at concentrations of 300 and 600 μg mL–1. Concentrations of 300 and 600 μg mL–1 significantly (p<0.0001) inhibited the growth of A. niger ATCC 16404 in Fig. 4a. The diameter of clear zone inhibition of A. niger ATCC 16404 in the agar-well diffusion assay was 19.97±2.040 and 10.97±0.5033 mm at concentrations of 300-600 μg mL–1 of AmBV, respectively in Fig. 4b.
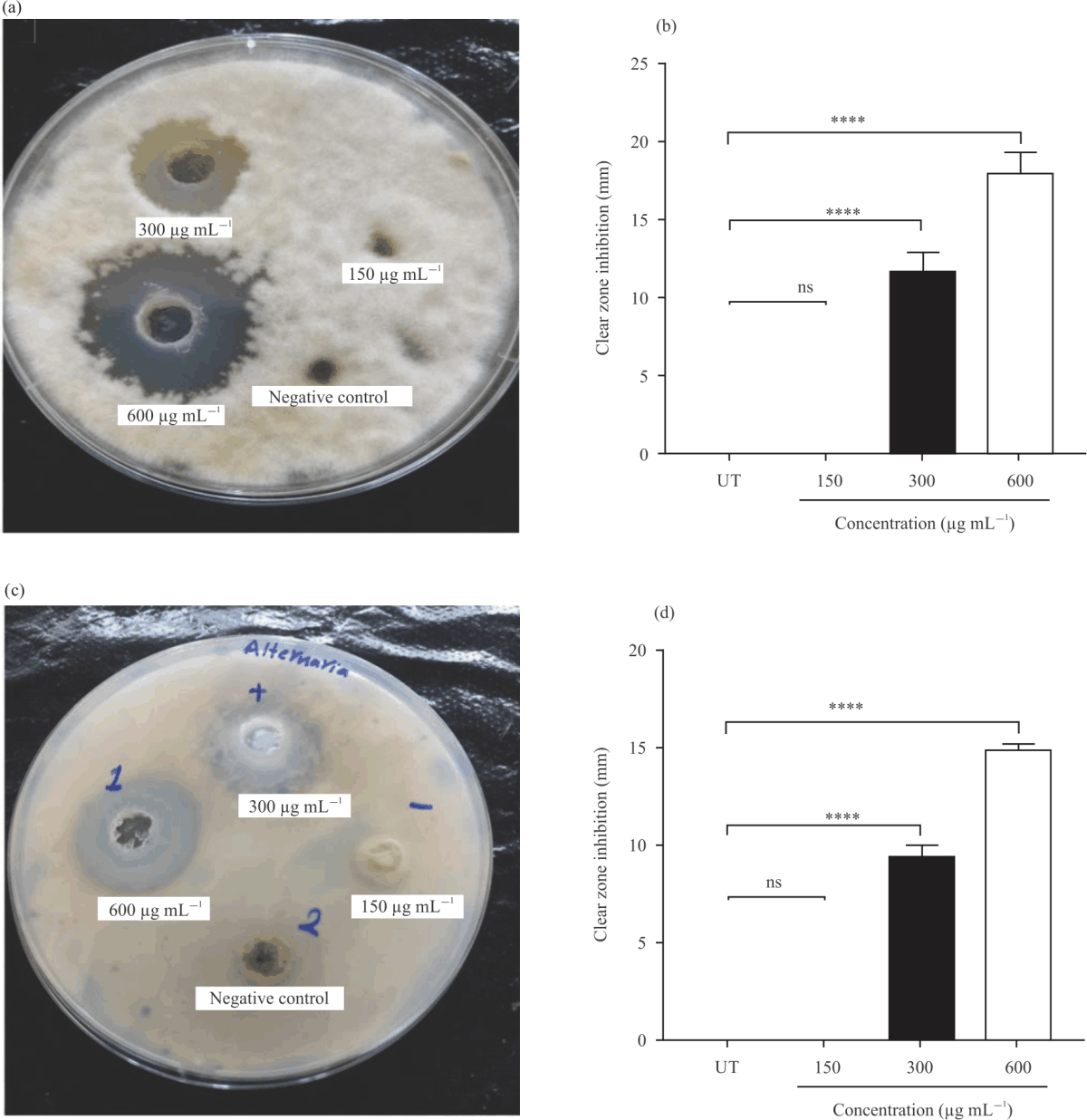 |
| Fig. 5(a-d): | Antifungal activity of different concentrations of AmBV against bacterial species, (a) Representative photographs of antifungal assays of F. oxysporum MLBM212, (b) F. oxysporum MLBM212 diameter of clear zone inhibition in millimeter (mm), (c) Representative photographs of antifungal assays of A. alternata MLBM09, (d) A. alternata MLBM09 diameter of clear zone inhibition in millimeter (mm) Results are shown as Means±SD of three independent experiments and p-value: ****p<0.0001 vs. negative control |
| Table 2: | MIC Values of AmBV |
| Bacterial isolates | MIC (μg mL–1) |
| Escherichia coli ATCC 8739 | 15.625 |
| Staphylococcus aureus ATCC 6538P | 31.25 |
| Serratia marcescens AUH 98 | 7.8 |
| Streptococcus mutans ATCC | 7.8 |
Similarly, AmBV significantly decreased the growth of A. flavus in Fig. 4c. The clear zone inhibition was 22.83±1.528 mm at a concentration of 300 μg Ml–1 of AmBV and 7.333±1.041 mm
at a concentration of 300 μg mL–1 of AmBV in Fig. 4d. Moreover, AmBV inhibited the growth of F. oxysporum MLBM212 in Fig. 5a and A. alternata MLBM09 in Fig. 5c. At a concentration of 300 μg mL–1 of AmBV, the diameters of clear zone inhibition of F. oxysporum MLBM212 in Fig. 5b and Alternaria alternata MLBM09 in Fig. 5d were 11.67±1.258 and 9.47±0.5508 mm, respectively. Whereas at a concentration of 600 μg mL–1 of AmBV, the diameters of clear zone inhibition of F. oxysporum MLBM212 (Fig. 5b) and Alternaria alternata MLBM09 (Fig. 5D) were 18.0±1.323 mm and 14.88±0.34 mm, respectively. There was no antifungal activity for AmBV against all tested fungi at a concentration of 150 μg mL–1.
DISCUSSION
New untraditional antibacterial and antifungal agents are highly required to tackle the problem of microbial resistance to available commercial antimicrobial substances. BV contains a variety of bioactive molecules including amino acids (aa), peptides, proteins, enzymes, sugars, biogenic amines, volatile compounds, phospholipids and pheromones. These biomolecules could be harnessed to develop effective antibacterial and antifungal agents, yet the antimicrobial potential of BV was not fully explored. Thus, in this work, the antimicrobial effect of Apis mellifera BV (AmBV) against some medically important and common bacterial and fungal pathogens was elucidated.
Results demonstrated that AmBV is highly potent in inhibiting the growth of the tested bacterial and fungal strains. Using broth microdilution method and CFU assays, it was discovered that AmBV significantly inhibits the growth of E. coli ATCC8739, S. aureus ATCC 6538P, S. marcescens AUH 98 and S. mutans ATCC 25175 in a concentration-dependent manner. Interestingly, in this study MIC values of AmBV against all tested bacteria were markedly low compared to the majority of the reported MIC values, which indicated the efficacy of AmBV as an antibacterial agent. The MIC value of AmBV for S. aureus was 31.25 μg mL–1 which is markedly lower than the MIC value reported by Samy et al.29 and Al-Ani et al.9. On the other hand, the MIC values for S. aureus were higher than what was demonstrated30,31. Similarly, the MIC values of AmBV were 15.625 μg mL–1 for E. coli ATCC8739 and 7.8 μg mL–1 for both S. marcescens AUH 98 and S. mutans ATCC 25175 which was much lower than the previously reported MIC values9,32-35.
Like the antibacterial activity of AmBV, antifungal activity was very prominent in the agar well diffusion assay. AmBV significantly inhibited the growth of A. niger ATCC 16404, F. oxysporum MLBM212, A. flavus and A. alternata MLBM09. Inhibition of the fungal growth required higher concentrations of AmBV compared to concentrations that were required for the inhibition of bacterial strains. Study detected a potent antifungal activity at a concentration of >300 μg mL–1 from AmBV. There was a significant increase in the clear zone diameter when fungi were treated with AmBV at concentrations of 300 and 600 μg mL–1. Similar to this study, the antifungal activity of AmBV has been evaluated by many other groups. Yu et al.12, demonstrated that AmBV inhibited the growth of Trichophyton mentagrophytes and Trichophyton rubrum by 92 and 32%, respectively. The antifungal inhibitory concentration reported in their study was 0.63 ppm which was higher than the current study. Similarly, BV has been reported to inhibit the growth of C. albicans and C. krusei at concentration of 60 to 125 μg mL–1 9,36.
Although, AmBV significantly decreased the growth of the tested bacteria and fungi, there were variations in antimicrobial potency and the inhibitory concentrations of AmBV between different bacterial and fungal strains. AmBV was more efficient in inhibiting the growth of E. coli ATCC8739, S. marcescens AUH 98 and S. mutans ATCC 25175 compared to S. aureus ATCC 6538P. As noted in the antibacterial activity of AmBV, there were also variations in the antifungal activities of AmBV. A. flavus and A. niger ATCC 16404 were the most sensitive to AmBV treatment followed by F. oxysporum MLBM212 and finally A. alternata MLBM09.
Variations in the antibacterial and antifungal potency of AmBV between the tested bacterial and fungal strains could be attributed to the nature of each microbial strain or the mechanism by which AmBV inhibits the growth of each microbial strain. AmBV contains a wide range of bioactive molecules that could exploit different strategies to inhibit each microbial strain37,38. Similarly, melittin which is one of the major components of AmBV is more active against gram-positive bacteria than gram-negative ones39. Similarly, PLA2 of AmBV inhibited the growth of Lactobacillus casei at a MIC value of 400 mg mL–1, however, it didn’t yield any satisfactory antibacterial effect against the growth of Streptococcus salivarius, S. sobrinus, S. mutans, S. mitis, S. sanguinis and Enterococcus faecalis32. Taken together variations in the antibacterial and antifungal potency of AmBV and its bioactive molecules could be exploited to develop a microbe-specific antimicrobial agent to avoid the side effects associated with the use of broad-spectrum antimicrobial agents on the beneficial normal flora.
CONCLUSION
Microbial diseases are the most prevalent health challenge particularly in developing countries due to drug resistance, high cost and high risk of synthetic and semisynthetic antibiotics. Therefore, new antimicrobial agents are in need to control microbial diseases. Natural products like BV are considered efficient alternatives to synthetic and semisynthetic antibiotics and a promising source for the discovery of new antimicrobial agents as they contain a variety of antimicrobial bioactive compounds. In this study, the antibacterial and antifungal activities of AmBV against medically important bacterial and fungal strains were demonstrated. AmBV significantly inhibited the growth of the bacterial and fungal pathogen, in vitro. There were variations in antimicrobial potency of AmBV between microbial strains that may use in a future study to invent new antimicrobial agents that have no side effects on the beneficial normal flora. This study provides a perspective on new antibacterial and antifungal therapeutic approaches using AmBV. Results from this study may, thus, could be utilized to develop cost-effective and efficient antibacterial and antifungal therapeutic agents.
SIGNIFICANCE STATEMENT
This study discovered that AmBV has strong antibacterial and antifungal activities against many medically important pathogens. This study will help researchers to exploit the antimicrobial activity of AmBV in future preclinical and clinical studies to develop cost-effective and nontraditional antibacterial and antifungal agents.
REFERENCES
- Kim, H., S.Y. Park and G. Lee, 2019. Potential therapeutic applications of bee venom on skin disease and its mechanisms: A literature review. Toxins, Vol. 11.
CrossRefDirect Link - Carpena, M., B. Nuñez-Estevez, A. Soria-Lopez and J. Simal-Gandara, 2020. Bee venom: An updating review of its bioactive molecules and its health applications. Nutrients, Vol. 12.
CrossRefDirect Link - Tu, W.C., C.C. Wu, H.L. Hsieh, C.Y. Chen and S.L. Hsu, 2008. Honeybee venom induces calcium-dependent but caspase-independent apoptotic cell death in human melanoma A2058 cells. Toxicon, 52: 318-329.
CrossRefDirect Link - Baracchi, D., S. Francese and S. Turillazzi, 2011. Beyond the antipredatory defence: Honey bee venom function as a component of social immunity. Toxicon, 58: 550-557.
CrossRefDirect Link - Han, S.M., J.M. Kim, I.P. Hong, S.O. Woo, S.G. Kim, H.R. Jang and S.C. Pak, 2016. Antibacterial activity and antibiotic-enhancing effects of honeybee venom against methicillin-resistant Staphylococcus aureus. Molecules, Vol. 21.
CrossRefDirect Link - Shin, S.H., M.K. Ye, S.Y. Choi and K.K. Park, 2017. The effects of melittin and apamin on airborne fungi-induced chemical mediator and extracellular matrix production from nasal polyp fibroblasts. Toxins, Vol. 9.
CrossRefDirect Link - Pak, S.C., 2016. An introduction to the toxins special issue on “bee and wasp venoms: Biological characteristics and therapeutic application”. Toxins, Vol. 8.
CrossRefDirect Link - Lin, T.Y. and C.L. Hsieh, 2020. Clinical applications of bee venom acupoint injection. Toxins, Vol. 12.
CrossRefDirect Link - AL-Ani, I., S. Zimmermann, J. Reichling and M. Wink, 2015. Pharmacological synergism of bee venom and melittin with antibiotics and plant secondary metabolites against multi-drug resistant microbial pathogens. Phytomedicine, 22: 245-255.
CrossRefDirect Link - Rady, I., I.A. Siddiqui, M. Rady and H. Mukhtar, 2017. Melittin, a major peptide component of bee venom, and its conjugates in cancer therapy. Cancer Lett., 402: 16-31.
CrossRefDirect Link - Falco, A., E. Barrajón-Catalán, M.P. Menéndez-Gutiérrez, J. Coll, V. Micol and A. Estepa, 2013. Melittin-loaded immunoliposomes against viral surface proteins, a new approach to antiviral therapy. Antiviral Res., 97: 218-221.
CrossRefDirect Link - Yu, A.R., J.J. Kim, G.S. Park, S.M. Oh, C.S. Han and M.Y. Lee, 2012. The antifungal activity of bee venom against dermatophytes. J. Appl. Biol. Chem., 55: 7-11.
CrossRefDirect Link - Allocati, N., M. Masulli, M.F. Alexeyev and C.D. Ilio, 2013. Escherichia coli in Europe: An overview. Int. J. Environ. Res. Public Health, 10: 6235-6254.
CrossRefDirect Link - Cristina, M.L., M. Sartini and A.M. Spagnolo, 2019. Serratia marcescens infections in neonatal intensive care units (NICUs). Int. J. Environ. Res. Public Health, Vol. 16.
CrossRefDirect Link - Kebriaei, R., K.L. Lev, R.M. Shah, K.C. Stamper and D.J. Holger et al., 2022. Eradication of biofilm-mediated methicillin-resistant Staphylococcus aureus infections in vitro: Bacteriophage-antibiotic combination. Microbiol. Spectr., Vol. 10.
CrossRefDirect Link - Azizi, A., Z. Amirzadeh, M. Rezai, S. Lawaf and A. Rahimi, 2016. Effect of photodynamic therapy with two photosensitizers on Candida albicans. J. Photochem. Photobiol. B: Biol., 158: 267-273.
CrossRefDirect Link - Al Mousa, A.A., H. Mohamed, A.M.A. Hassane and N.F. Abo-Dahab, 2021. Antimicrobial and cytotoxic potential of an endophytic fungus Alternaria tenuissima AUMC14342 isolated from Artemisia judaica L. growing in Saudi Arabia. J. King Saud Univ. Sci., Vol. 33.
CrossRefDirect Link - Surendra N.S., G.N. Jayaram and M.S. Reddy, 2011. Antimicrobial activity of crude venom extracts in honeybees (Apis cerana, Apis dorsata, Apis florea) tested against selected pathogens. Afr. J. Microbiol. Res., 5: 2765-2772.
CrossRefDirect Link - Abdelgaleil, S.A.M., M.M.G. Saad, N.R. Ariefta and Y. Shiono, 2020. Antimicrobial and phytotoxic activities of secondary metabolites from Haplophyllum tuberculatum and Chrysanthemum coronarium. South Afr. J. Bot., 128: 35-41.
CrossRefDirect Link - Krasnodembskaya, A., Y. Song, X. Fang, N. Gupta, V. Serikov, J.W. Lee and M.A. Matthay, 2010. Antibacterial effect of human mesenchymal stem cells is mediated in part from secretion of the antimicrobial peptide LL-37. Stem Cells, 28: 2229-2238.
CrossRefDirect Link - Bergsson, G., E.P. Reeves, P. McNally, S.H. Chotirmall and C.M. Greene et al., 2009. LL-37 complexation with glycosaminoglycans in cystic fibrosis lungs inhibits antimicrobial activity, which can be restored by hypertonic saline. J. Immunol., 183: 543-551.
CrossRefPubMedDirect Link - Sung, D.K., Y.S. Chang, S.I. Sung, H.S. Yoo, S.Y. Ahn and W.S. Park, 2016. Antibacterial effect of mesenchymal stem cells against Escherichia coli is mediated by secretion of beta-defensin-2 via toll-like receptor 4 signalling. Cell. Microbiol., 18: 424-436.
CrossRefDirect Link - Bhattacharya, D., D.S. Sayi, R. Thamizhmani, H. Bhattacharjee, A.P. Bharadwaj, A. Roy and A.P. Sugunan, 2012. Emergence of multidrug-resistant Vibrio cholerae O1 biotype El Tor in Port Blair, India. Am. Soc. Trop. Med. Hyg., 86: 1015-1017.
CrossRefDirect Link - Chen, L., E.E. Tredget, P.Y.G. Wu and Y. Wu, 2008. Paracrine factors of mesenchymal stem cells recruit macrophages and endothelial lineage cells and enhance wound healing. PLoS ONE, Vol. 3.
CrossRefDirect Link - Chung, H.S., S.K. Kim, C. Hahm and M. Lee, 2022. Performance evaluation of the VITEK2 and sensititre systems to determine colistin resistance and MIC for Acinetobacter baumannii. Diagnostics, Vol. 12.
CrossRefDirect Link - Gonelimali, F.D., J. Lin, W. Miao, J. Xuan, F. Charles, M. Chen and S.R. Hatab, 2018. Antimicrobial properties and mechanism of action of some plant extracts against food pathogens and spoilage microorganisms. Front. Microbiol., Vol. 9.
CrossRefDirect Link - Daoud, A., D. Malika, S. Bakari, N. Hfaiedh, K. Mnafgui, A. Kadri and N. Gharsallah, 2019. Assessment of polyphenol composition, antioxidant and antimicrobial properties of various extracts of date palm pollen (DPP) from two Tunisian cultivars. Arabian J. Chem., 12: 3075-3086.
CrossRefDirect Link - Mosquera, O.M., Y.M. Correa and J. Nino, 2004. Antibacterial activity of some Andean Colombian plants. Pharm. Biol., 42: 499-503.
CrossRefDirect Link - Samy, R.P., P. Gopalakrishnakone, M.M. Thwin, T.K.V. Chow, H. Bow, E.H. Yap and T.W.J. Thong, 2007. Antibacterial activity of snake, scorpion and bee venoms: A comparison with purified venom phospholipase A2 enzymes. J. Appl. Microbiol., 102: 650-659.
CrossRefPubMedDirect Link - Choi, J.H., A.Y. Jang, S. Lin, S. Lim and D. Kim et al., 2015. Melittin, a honeybee venom-derived antimicrobial peptide, may target methicillin-resistant Staphylococcus aureus. Mol. Med. Rep., 12: 6483-6490.
CrossRefDirect Link - Pereira, A.F.M., M. Albano, F.C.B. Alves, B.F.M.T. Andrade and A. Furlanetto et al., 2020. Influence of apitoxin and melittin from Apis mellifera bee on Staphylococcus aureus strains. Microb. Pathogen., Vol. 141.
CrossRefDirect Link - Leandro, L.F., C.A. Mendes, L.A. Casemiro, A.H.C. Vinholis, W.R. Cunha, R. de Almeida and C.H.G. Martins, 2015. Antimicrobial activity of apitoxin, melittin and phospholipase A2 of honey bee (Apis mellifera) venom against oral pathogens. An. Acad. Bras. Ciênc., 87: 147-155.
CrossRefDirect Link - Zolfagharian, H., M. Mohajeri and M. Babaie, 2016. Bee venom (Apis mellifera) an effective potential alternative to gentamicin for specific bacteria strains. J. Pharmacopuncture, 19: 225-230.
CrossRefDirect Link - Tanuwidjaja, I., L. Svečnjak, D. Gugić, M. Levanić, S. Jurić, M. Vinceković and M.M. Fuka, 2021. Chemical profiling and antimicrobial properties of honey bee (Apis mellifera L.) venom. Molecules, Vol. 26.
CrossRefDirect Link - Maitip, J., W. Mookhploy, S. Khorndork and P. Chantawannakul, 2021. Comparative study of antimicrobial properties of bee venom extracts and melittins of honey bees. Antibiotics, Vol. 10.
CrossRefDirect Link - Lee, S.B., 2016. Antifungal activity of bee venom and sweet bee venom against clinically isolated Candida albicans. J. Pharmacopuncture, 19: 45-50.
CrossRefDirect Link - El-Seedi, H., A.A. El-Wahed, N. Yosri, S.G. Musharraf and L. Chen et al., 2020. Antimicrobial properties of Apis mellifera's bee venom. Toxins, Vol. 12.
CrossRefDirect Link - El Mehdi, I., S.I. Falcão, M. Harandou, S. Boujraf and R.C. Calhelha, 2021. Chemical, cytotoxic, and anti-inflammatory assessment of honey bee venom from Apis mellifera intermissa. Antibiotics, Vol. 10.
CrossRefDirect Link - Čujová, S., L. Bednárová, J. Slaninová, J. Straka and V. Čeřovský, 2014. Interaction of a novel antimicrobial peptide isolated from the venom of solitary bee Colletes daviesanus with phospholipid vesicles and Escherichia coli cells. J. Pept. Sci., 20: 885-895.
CrossRefDirect Link