
Sri Darmawati
Master Program of Clinical Laboratory Science, Postgraduate School, Universitas Muhammadiyah Semarang, Jl. Kedungmundu Raya No. 18, Semarang, Jawa Tengah 50273, Indonesia
LiveDNA: 62.39768
ORCID: 0000-0002-9415-1955
I. Agus Adi Gunawan
Master Program of Clinical Laboratory Science, Postgraduate School, Universitas Muhammadiyah Semarang, Jl. Kedungmundu Raya No. 18, Semarang, Jawa Tengah 50273, Indonesia
Nastiti Wijayanti
Faculty of Biology, Universitas Gadjah Mada, Jl. Teknika Selatan, Sekip Utara, Bulaksumur, Sleman, Yogyakarta 55281, Indonesia
LiveDNA: 62.32258
ORCID: 0000-0001-6483-2816
Muchamad Dafip
Department of Biology, Faculty of Mathematics and Natural Sciences, Universitas Negeri Semarang, Jl. Taman Siswa, Kampus Sekaran, Gunung Pati, Semarang, Central Java 50229, Indonesia
LiveDNA: 62.35020
ORCID: 0000-0001-8059-8193
Pakistan Journal of Biological Sciences
Year: 2022 | Volume: 25 | Issue: 12 | Page No.: 1085-1093







ABSTRACT
Background and Objective: The flagellin of Salmonella typhi is potentially developed as an identifying antigen in a rapid diagnostic test instrument that may be more accurate than conventional serological tests. Therefore, this study aims to analyze the immunogenicity of flagellin S. typhi as the basis for developing a typhoid fever diagnostic. Materials and Methods: Flagellin was isolated from the bacterial culture of S. typhi serovar Semarang and used as the primary antigen for vaccine assembly. Native flagellin antigen was immunized in Balb/C mice with injection doses of 2, 3, 4, 5 and 6 g/100 L in each group (K0-K5), respectively, via intraperitoneal cavity. Blood serum was collected to ELISA based-measurement for IL-6 and TNF-a titers. Then, specific immunoglobulin (Ig) of anti-flagellin was detected using in-house ELISA and western blotting. Results: The findings in this study showed that immunization at the dose of 4-5 g/100 L significantly decreased the IL-6 titer, i.e., 8.33±0.87 pg mL–1, compared to control. The antibody titer test analysis showed the highest Ig-G anti-flagellin was found in K4 mice after immunization using a dose of 5 g/100 L with an average absorbance of Ig-G reaching 1.19 ±0.32. Conclusion: The results indicated that the flagellin protein of S. typhi serovar Semarang induces adaptive immune responses and produces specific antibodies against flagellin. The immunogenic properties of the flagellin protein of S. typhi serovar Semarang potentially developed as a specific diagnostic marker. Further research may also focus on a beneficial feature of flagellin as a vaccine candidate.
PDF Abstract XML References Citation
Copyright: © 2022. This is an open access article distributed under the terms of the creative commons attribution License, which permits unrestricted use, distribution and reproduction in any medium, provided the original author and source are credited.
How to cite this article
Sri Darmawati, I. Agus Adi Gunawan, Nastiti Wijayanti and Muchamad Dafip, 2022. Immunogenicity Response of Mus musculus var. Balb/C after Immunization using Flagellin Salmonella typhi Serovar Semarang. Pakistan Journal of Biological Sciences, 25: 1085-1093.
DOI: 10.3923/pjbs.2022.1085.1093
URL: https://scialert.net/abstract/?doi=pjbs.2022.1085.1093
DOI: 10.3923/pjbs.2022.1085.1093
URL: https://scialert.net/abstract/?doi=pjbs.2022.1085.1093
INTRODUCTION
Salmonella typhi (S. typhi ) is an opportunistic bacterial pathogen that causes typhoid fever which is transmitted through contaminated food or drink (food-born disease). The pathogenicity of S. typhi is influenced by the adhesion ability to the small intestinal mucosa, penetrating into lymphoid tissue and spreading in the body through the bloodstream1,2. Under unfavorable conditions, S. typhi produces exotoxins and endotoxins to protect against the body’s immune system. S. typhi endotoxin is a robust immune generator that triggers an infection that is manifested as fever, nausea, fatigue, organ illness and other symptoms of infection3,4. In particular conditions, Salmonella can silence the immune response that increases in cases of carriers and resistance becomes a problem in controlling typhoid fever5.
In some cases, the characteristic symptoms of typhoid fever cannot be distinguished from other bacterial or viral infections (underdiagnosis). It contributes to misinformation during the diagnosis procedure, resulting in inappropriate antibiotic treatment and increasing the risk of pathogenic multidrug-resistant S. typhi6,7. Therefore, non-specific clinical symptoms must be supported by accurate laboratory tests.
Currently, the Widal Test is a common laboratory diagnosis to identify typhoid fever because it is a fast result, cheap and straightforward, although less specific8,9. Furthermore, the diagnostic test is improved to detect specific antibodies of S. typhi and used as a foundation for serology-based rapid tests, such as Tubex and Typhidot. Tubex Test detects immunoglobulin-M (Ig-M) against S. typhi O9-lipopolysaccharide (LPS) antigen, while Typhidot, a dot-enzyme immunoassay, is used to detect IgM and IgG outer membrane protein antigens10-12. However, serology-based diagnosis has fluctuating sensitivity and specificity because of LPS cross-reaction to other bacterial infections. Therefore, an effective diagnostic test should be developed using specific features or components of S. typhi, which are highly immunogenic.
One of the protein features recognized in most S. typhi various sub-species that potentially can be developed as an immunogenic antigen is flagellin, a complex part of flagella. Flagellin protein is an external filament consisting of repeating copies of several structural proteins13. Salmonella typhi flagella is responsible for host cell invasion and considered as virulence properties that induce infection14,15. Recent studies also reveal that flagella involve in an escape mechanism against immune cells5,16,17. Due to its vital function, the flagella proteins, including flagellin, is conserved and become specific characteristic of S. typhi markers. Several studies have shown that flagellin is an essential ligand for the Toll-Like Receptor 5 (TLR5) protein and is involved in immune system activation, including the production of inflammatory-related cytokines, adaptive immune responses and produces specific antibodies14. Therefore, this study aims to analyze the immunogenicity of S. typhi flagellin as the basis for developing a typhoid fever diagnostic.
MATERIALS AND METHODS
This study was an experimental research: Post-only control group design to determine the ability of the flagellin protein of S. typhi serovar Semarang to induce an adequate immune response. The research was carried out at the Laboratory of Microbiology and Molecular Biology, Department of Health Analyst, Faculty of Nursing and Health Sciences, Universitas Muhammadiyah Semarang and Biochemistry Laboratory, Center for Biotechnology Studies, Universitas Gadjah Mada. Animal concern has been approved ethically by the Health Research Ethics Commission, Faculty of Public Health, Universitas Muhammadiyah Semarang, No: 578/KEPK-FKM/UNIMUS/2021.
Salmonella typhi sample isolate and culture: The S. typhi serovar Semarang isolate was obtained from the Semarang Center for Health Laboratory of Semarang City. Salmonella typhi colony stock was inoculated on MacConkey Agar (MCA) media, Cat. No.: A5306 (Merck, Sigma-Aldrich: Jakarta, Indonesia) and then planted in 50 mL of Brain-Heart Infusion (BHI) liquid media, Cat. No.: 53286 (Merck, Sigma-Aldrich: Jakarta, Indonesia), then incubated at 37°C for 24 hrs, the culture was stored and used as a starter. The bacteria starter was then mixed well with 500 mL of BHI liquid media until homogenous, after that, incubated at 37°C. The cultured S. typhi was ready to be harvested after 48 hrs of culture.
Isolation of flagellin Salmonella typhi serovar Semarang: Bacterial cultures aged 48 hrs were centrifuged at 8000 rpm at 4°C for 30 min. The supernatant was discarded and the pellet was compacted into a thick suspension by adding 2 mL of PBS 1X. The suspension was acidified by adding 1M HCl until it reached pH 2, then homogenized with a stirrer for 30 min. The suspension was centrifuged again at 8000 rpm at 4°C for 30 min, the supernatant was taken and centrifuged again at 12,000 rpm at 4°C for 30 min. The supernatant was added with 1M NaOH to pH 7.2 (a low concentration of flagella was obtained). Then, added ammonium sulfate, as much as 40% of the suspension volume and stirred until dissolved. The flagellin protein was centrifuged at 12,000 rpm for 15 min. The pellet obtained was added with 0.5 mL of PBS 1×, suspended and dialyzed against a PBS solution of 0.5X pH 7.4 following the protocol from Senevirathne et al.18 with modification.
Preparation of Balb/C mice: A total of 36 female Balb/C mice aged 8-10 weeks-old weighing ±30 g were divided into six groups, namely the control group (K0), which was given placebo treatment. In contrast, the five treatment groups received antigen immunization with different concentrations (K1-K5) of 2, 3, 4, 5 and 6 μg/100 μL, respectively. Each group was kept in cages measuring 50×40×15 cm and placed in a maintenance room at a temperature of 26-28°C with lighting for 12 hrs. Feeding and drinking were done ad libitum and refilled every morning and evening.
Immunization of Balb/C mice with flagellin to induce IL-6 and TNF-α and antibody: The vaccine was made with antigen emulsion at 2, 3, 4, 5 and 6 μg of flagellin in 50 μL PBS 1× for all three immunization stages. Each vaccine dose was added with complete Freund’s adjuvant solution in a ratio of 1:1 (v/v), then re-emulsified to get a final volume of up to 100 μL/head/injection. The first dose was administered intraperitoneally on the first day of treatment. Approximately 1.5 mL of blood was drawn through the hearts of mice for each mouse on the 7th day from the first immunization. The serum was separated by centrifugation and used for IL-6 and TNF-α titers analysis.
The vaccination process continued for the second immunization carried out on day 14th, then the booster or the third stage was carried out on day 28th. On day 32, approximately 1.5 mL of mouse blood was taken from the heart for each mouse, the serum was separated by centrifugation, then the serum was checked for antibody titer by in-house ELISA based on the absorbance value using an ELISA reader and specificity by western blotting.
Measurement of IL-6 and TNF-α levels: The serum obtained from each mouse in each group was measured for levels of IL-6 and TNF-α. A total of 100 L for the measurement of cytokine levels was carried out using the sandwich ELISA method and the Elikine Mouse IL-6 ELISA Kit reagent kit, Cat. No.: KET7009TNF and Elikine Mouse TNF-α ELISA Kit, Cat. No.: KET7015 (Abbkine: Wuhan, China) according to manufacturer’s procedure. The optical density (OD) of the reaction between horseradish peroxidase (HRP)-conjugated Streptavidin and the sample substrate was read with a MaxSignal™ 6000 Bio Microplate ELISA reader (Bioo Scientific: Austin, Texas, USA) at a wavelength of 450 nm. The IL-6 levels were then calculated using a standard curve, which showed the OD value and IL-6 levels.
Flagellin antibody identification and measurement
In-house ELISA: Antigen preparation for coating was carried out by dissolving 5 g mL–1 of antigen in a coating buffer. One hundred microliters of coating antigen were added to each well and incubated for 24 hrs at 37°C water bath. Then washed three times with a washing buffer of 200 L/well for each wash. Blocking was carried out using 1% BSA in 1X PBS buffer by adding 100 L of blocking solution to each well, incubating for ±2 hrs at room temperature. Then washed three times with a washing buffer of 200 L/well for each wash.
In the dry well was added 100 L/well of primary antibody (anti-flagellin antibody) with 700X dilution (optimized result), incubated in a water bath at 37°C for 1 hr. The well was washed thrice with a washing buffer of 200 L/well for each wash. The cleaned wells were added with 100 L/well of secondary antibody (conjugate anti-mouse IgG (whole molecule) alkaline phosphatase antibody produced in goat SIGMA) and incubated for 1 hr in a 37°C water bath. Then washed three times with a washing buffer of 200 L/well for each wash. Cleaned wells were added 100 L substrate solution to each well and incubated for 15-30 min in a 37°C water bath in the dark. Then read the absorbance with an ELISA reader at a wavelength of 405 nm.
Antibody specificity test by western blotting: Identifying proteins recognized by antibodies was carried out using the western blotting method. Protein isolates were run on polyacrylamide gel with a voltage of 100 V for 120 min. Then the running polyacrylamide gel was put in a container containing the transfer buffer. Prepared apparatus for transfer of protein from polyacrylamide gel to nitrocellulose. Protein transfer was carried out by placing a polyacrylamide gel between four layers of Whatman paper No. 41 on the bottom and four layers on the top. Each layer was moistened with transfer buffer and removed air bubbles, then electrified with a current of 500 mA for 60 min. Furthermore, the nitrocellulose membrane was immersed in 0.5% TBS-Tween for 30 min by shaking on a shaker. After that, the solution was discarded and continued with immersion using a blocking buffer at 4°C for 24 hrs.
The membrane that had passed the blocking stage was washed three times using 0.05% TBS-Tween in a shaker for 5 min for each wash. Furthermore, the membrane was immersed in the primary antibody solution at room temperature for 1 hr. The membrane was washed thrice with 0.05% TBS-Tween in a shaker for 5 min for each wash. The membrane was immersed in a conjugate solution consisting of 25 mL of incubation buffer, 0.25 g of BSA and 25 L of the conjugate and incubated at room temperature for 1 hr. Furthermore, the membrane was washed two times with 0.05% TBS-Tween for 5 min and the last washing step was conducted without Tween. The staining step was carried out in a dark room by immersing the nitrocellulose membrane in a combination of nitro-blue tetrazolium chloride and 5-bromo-4-chloro-3-indolyl phosphate p-toluidine salt (NBT-BCIP) until a band was formed and added aquabides when the band was visible.
Data analysis: Primary data in the 6, TNF-titers and antibody absorbance of mice were statistically analyzed using SPSS, version 22, with a 95% confidence level and a significance level of 0.050. The assumption of data normality was met based on the Shapiro-Wilk normality test, then continued with the different tests using One-way ANOVA and the least significant difference (LSD) test. The western blot specificity visualization data were analyzed narratively by comparing the presence of protein bands at 104, 64 and 50 kDa.
RESULTS
Post-immunization IL-6 and TNF-α levels: The Balb/C mice’s immune response after immunization was performed by decreased IL-6 titer and detected antibody production against flagellin. However, the decrease in IL-6 titer was not accompanied by a change in TNF-α titer. The concentration of IL-6 titer experienced a downward trend alongside the increased dose of the flagellin vaccine (Table 1).
An immunization dose of 4 μg/100 μL flagellin significantly reduced IL-6 levels and is considered the optimum dose for enhancing the immune response in mice. Based on these data, 4 and 5 g/100 L became the threshold doses for changes in IL-6 titer compared to the control and low-dose immunization groups. It shows that the more concentration of antigen dose for immunization, the more IL-6 is suppressed. However, it did not change TNF-α titer.
Antibody absorbance of Balb/C mice and detection of Ig-G against flagellin: In addition to inducing changes in the serum IL-6 titer of Balb/C mice, flagellin immunization also triggered a humoral response indicated by immunoglobulin-G (Ig-G) production. Anti-flagellin Ig-G detection showed that the antibody absorbance was significantly increased at doses of 4, 5 and 6 g/100 L of vaccine injection (Fig. 1).
Antibody absorbance indicates that anti-flagellin Ig-G was significantly increased in the K3-K5 group after complete immunizations. The highest average of antibody absorbance was obtained from a dose of 5 g/100 L vaccine, even though it was not significantly different from the K3 group. This result was relevant to the IL-6 production, which may relate to the humoral response in mice. Furthermore, increasing the vaccine dose up to 6 g/100 L in K5 decreases the absorbance of anti-flagellin Ig-G compared to the K4 group but is higher than the control group. Decreased Ig-G in higher vaccine doses indicates an immunogenic tolerance effect that suppresses antibody production. On the other hand, increasing the vaccine dose may not always be proportional to an increase in antibody titer.
The anti-flagellin Ig-G synthesis was confirmed by the presence of protein bands at sizes of 104, 64 and 50 kDa by SDS-Page and western blotting methods (Fig. 2). In the result, the protein bands observed in all treatment groups represented by K2 for low doses and K4 for optimum dose. The observed protein was validated using flagellin protein flagellin for positive control and K0 for negative control.
| Table 1: | IL-6 and TNF-α levels in the blood of Balb/C mice seven days after immunization with the flagellin of S. typhi serovar Semarang | ||
| Group | Flagellin (μg/100 μL) | IL-6 (pg mL–1) | TNF-α (pg mL–1) |
| K0 | 0.00 | 13.22±1.94a | 23.93±2.73 |
| K1 | 2.00 | 14.12±3.94a | 19.98±2.30 |
| K2 | 3.00 | 12.05±3.46a | 20.25±2.23 |
| K3 | 4.00 | 8.34±0.36bc | 29.56±1.98 |
| K4 | 5.00 | 6.08±0.76c | 22.74±4.24 |
| K5 | 6.00 | 5.59±0.37cd | 21.62±0.63 |
| p-value | 0.005 | 0.286 | |
| a-dSignificant difference between treatment groups based on One-way ANOVA at 95% confidence level and significance level (p-value) 0.050 | |||
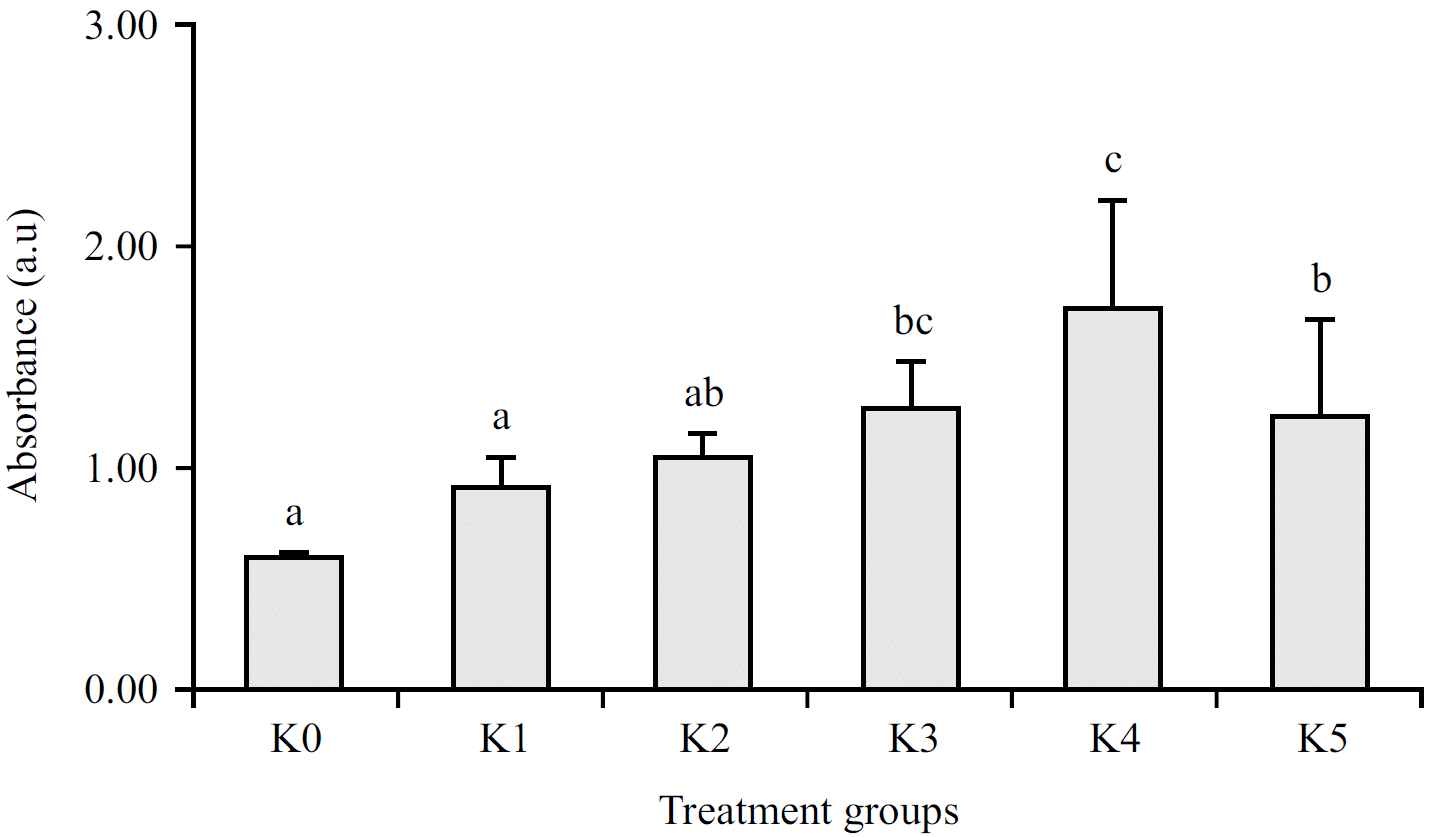 |
| Fig. 1: | Antibody absorbance of Balb/C mice after immunization with flagellin protein S. typhi at OD 405 Alphabet above the bar shows a significant difference between treatment groups based on one-way ANOVA at 95% confidence level and significance level = 0.416 (p<0.05) |
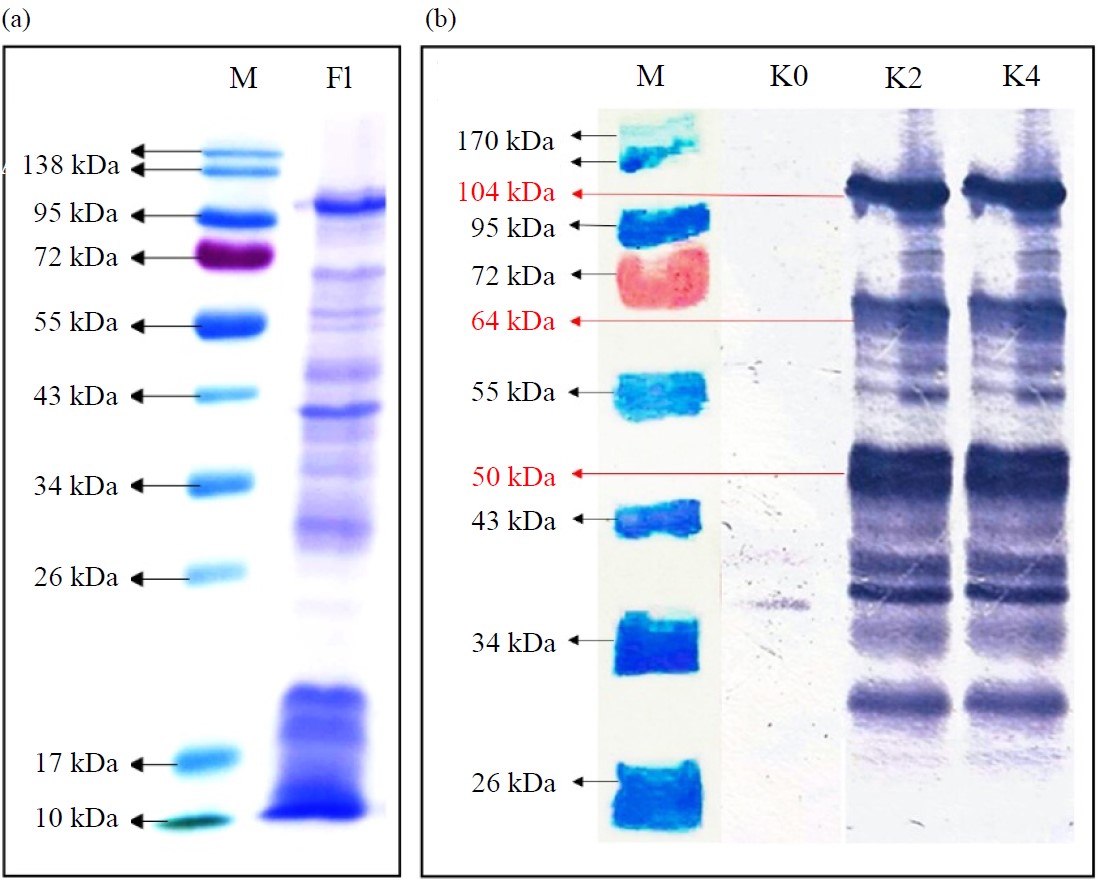 |
| Fig. 2(a-b): | Visualization of protein S. typhi serovar Semarang (a) SDS-PAGE result and (b) Western blotting result (a) M: Markers, F1: Flagellin protein samples and (b) M: Marker, K0: Control, K2 and K4 sample |
The visualization results show the presence of anti-flagellin Ig-G, which recognizes flagellin protein and sub-unit bands. The anti-flagellin Ig-G is indicated by the appearance of blue reaction color as the reaction product of flagella sub-unit proteins with the Ig-G anti-flagellin in mice. Furthermore, the strongest interactions were formed at the sub-unit proteins of 104, 64 and 50 kDa, which is observed by the dark blue color. This result indicates that flagellin protein subunits 104, 64 and 50 kDa are the main immunogenic protein subunits. However, the polypeptide structure and amino acid sequence of the flagellin sub-unit proteins at each identified molecular weight still need further study to determine the helix and sheet structure exposed as the epitope.
DISCUSSION
Salmonella typhi flagella are composed of thousands of copies of a single protein flagellin forming an external filament for motility, attachment and penetration into host cells19-21. Invasion of S. typhi resulted in flagellin recognition as an antigen, thereby inducing activation of the adequate immune system. Furthermore, in recent research, vaccine-based flagellin has also been reported to cause the activation of humoral and cellular immune systems22.
The basic structure of flagella consists of a ring, a GOLD ring, a P ring and an L ring located in the cytoplasm, cytoplasmic membrane, peptidoglycan layer and outer membrane, respectively23. During MS ring formation, FlhA, FlhB, fLIp, FliQ and FliR converge to one with the help of FliO to become a transmembrane export gate. FliM and FliN will bind to FliG to form ring C in the cytoplasm24. The FlgK dan FlgL form a splice structure so that FliD can form a filament cap at the end of the junction to induce FliC assembly into the filament25.
Circular dichroism spectroscopy revealed that flagellins derived from various species and Salmonella serovars have at least a secondary structure riches with α-helix synthesized by conserved domains D0 and D1 genes encoding both N and C terminals26. A conserved structure at each terminal protects the FljB ring and FLiC filament protein structure, which is dominated by β-sheets, from protein degradation27. Furthermore, the conserved helix structure in flagellin also plays an essential function in stimulating the TLR5 receptor of the host cell28. It is caused by pathogen-associated molecular pattern (PAMP), a locus in FLiC protein synthesized by the fliC gene for TLR5 recognition14,29.
In mammals, the ability of flagellin to induce the host’s immune responses may occur via three different signaling pathways. First, the binding complex formed by the interaction of TLR5 with PAMP site triggers the activation of the phosphorylated signaling pathway30. Immune cells, especially macrophages and neutrophils involve in innate immunity and respond to extracellular monomeric flagellin via the TLR5 homodimer complex on the cell surface31. This flagellin binding complex induces TLR5 dimerization, then recruits myeloid differentiation primary response 88 (MyD88) and Mal/TIRAP, which forwards the signal to the Mitogen-Activated Protein Kinase (MAPK) cascade, thereby inducing transcription factor Activator Protein-1 (AP-1)32,33. Furthermore, MyD88 recruitment activates the Interleukin-1 Receptor-Associated Kinase 4 (IRAK4) component complex, IRAK1, TNF Receptor-Associated Factors-6 (TRAF6) and IκB kinase34,35. Activation of IκB kinase causes localization of phosphorylation of proteins 65 and 50 (P65/P50), which then induces nuclear factor kappa-light-chain-enhancer of activated B cells (NF-κB) towards the nucleus and acts as an initiation factor for the synthesis of inflammatory-related cytokines. In addition, NF-κB then induces the expression of various genes involved in canonical and innate proinflammatory pathways and adaptive immune responses36.
In the early stage, TNF-α binds to TNF-Receptor Type 1-Associated DEATH Domain (TRADD) protein as the adaptor molecule that initiates signaling to activate NF-κB. Further process, TRADD recruits TRAF2, TRAF5 and receptor-interacting protein kinases (RIP)37. These adaptor protein complex then inducing kinase (NIK) and delivers the signal to the multicomponent protein, including IκB kinase (IKK), enabling the formation of IKKα/IKKβ complex. The complex then activates NF-κB (p65/p50 heterodimer) by replacing ubiquitin (Ub) inhibitory. The Ub protein which normally binds to NF-κB, is phosphorylated by IκK kinase and then degraded together with releasing IκBα. The NF-κB then moved into the nucleus to bind with transcription factors to initiate protein inflammation gene expression, including TNF-α and IL-6 production37,38. According to Khani et al.39, IL-6 production in mice significantly increased 24 hrs after flagellin immunization. Furthermore, Barkhordari et al.29 explained that S. typhi flagellin induces and enhances immune response by increasing TNF-α expression.
Second, in some cases, flagellin initiates infectious action via TLR5/TLR4 heterodimer and induces the IRF3 pathway without involving MyD88 to synthesize interferon-β (IFN-β)40. Furthermore, IFN-β induces inducible nitric oxide synthase (iNOS) gene transcription by activating the Signal Transducer and Activator of Transcription-1 (STAT1), which increases nitric oxide production. Third, antigen recognition occurs during flagellin penetration into the host cell’s cytoplasm via the Type III Secretion System (T3SS)41. Then it is detected by the nod-like receptor family CARD domain-containing protein 4 (NLRC4) inflammasome complex and transmits signaling pathways to the caspase-1 (cas-1), cas-7 and NLRP3 inflammasome, resulting in the synthesis of IL-1 family42.
In this study, decreased IL-6 and no change in TNF-α might indicate that TLR5/TLR4 heterodimers dominate the activated Balb/C mouse immune cell cascade with low affinity to the NF-κB cascade. In addition, the involvement of TLR5 homodimers primarily triggers the MAPK cascade and TIR-domain-containing adapter-inducing interferon-β (TRIF) to synthesize IL-1β, IL-1, IL-12 and IFN-γ. Although the role of IL-6 and TNF-α has not been confirmed in flagellin-based immunization, considering that finding these two cytokines might act simultaneously as pro and anti-inflammatory43. Therefore, it requires a comprehensive understanding of cytokine variations and action in synergistic and antagonistic interactions. Furthermore, several studies have also shown that the interaction of TLR5 with flagella potentially has different responses depending on the exposed cell type and host species44. For example, A higher IL-8 production is observed when the flagellar protein binds to TLR5 in epithelial cells, whereas, the interaction between monocytes and dendritic cells induces TNF-α secretion in humans39. Another example shows that TLR5 of the small intestinal epithelial cells interacting with flagellin triggers the secretion of pro-inflammatory IL-6, IL-12 and TNF-α and induces the production of anti-flagellin Ig-G. Suppression of NF-κB may also occur due to a Salmonella defensive mechanism through SpvB protein. This Salmonella-specific cytotoxic protein prevents NF-κB activation by targeting IκB kinase-b (IKKb)45. However, this condition was neglected in this study because only the small flagellum part was used as a vaccine antigen. Although there was no different TNF-α titer among groups, the flagellin S. typhi serovar Semarang immunization activated the humoral immune system and produced specific anti-flagellin Ig-G antibodies. Specifically, the resulting antibody binds to flagellin protein fragments used as attachment sites for both SDS-Page and western blotting. This result was relevant to the previous studies where the profile of immunogenic flagellin-specific protein fragments producing antibodies was found at 64 and 50 kDa3,29. The binding site fitness promotes flagellin protein fragments as a recognition site to develop of S. typhi serovar Semarang diagnostic kit or vaccine candidate.
CONCLUSION
Immunization with a 4-5 g/100 L flagellin S. typhi serovar Semarang triggered a specific immune response in Balb/C mice by decreasing IL-6 secretion. Even though the TNF-α titers were not significantly different between the control and treatment groups, S. typhi flagellin protein immunized in Balb/C mice could induce specific antibody production. The highest average antibody absorbance results were obtained from Balb/C mice, immunized using 5 g/100 L vaccine, reaching 1.19±0.32 at OD 405. The visualization results showed that a bond was formed between the antibody and primary antigen on the tape strip. The observed protein is sized at 104, 64 and 50 kDa. These findings confirmed the potential of flagellin as an immune generator and potentially be used in the development of diagnostic tests or vaccine candidates. Further research may focus on identifying and developing epitope-potential protein sequences and a retest to determine its specifications and sensitivity in recognizing S. typhi infection.
SIGNIFICANCE STATEMENT
This study aims to understand the immunogenicity reaction caused by flagellin protein in the flagella of S. typhi serovar Semarang. The flagellin protein is a promising potential feature to be developed as a specific vaccine against typhi fever and a marker for early Rapid Diagnostic Tests (RDT). Further research may also focus on protein sequence and structure to discover specific epitope regions of flagellin for vaccine candidates or RDT development.
ACKNOWLEDGMENTS
Special thanks to the Ministry of Education and Culture of the Republic of Indonesia for fully funding this research through Lembaga Layanan Perguruan Tinggi Wilayah IV, grant number: 072/E5/PG.02.00.PT/2022.
REFERENCES
- Habte, L., E. Tadesse, G. Ferede and A. Amsalu, 2018. Typhoid fever: Clinical presentation and associated factors in febrile patients visiting Shashemene Referral Hospital, Southern Ethiopia. BMC Res. Notes, Vol. 11.
CrossRefDirect Link - Kuijpers, L.M.F., P. Chung, M. Peeters, M.F. Phoba and C. Kham et al., 2018. Diagnostic accuracy of antigen-based immunochromatographic rapid diagnostic tests for the detection of Salmonella in blood culture broth. PLoS ONE, Vol. 13.
CrossRefDirect Link - Darmawati, S. and A.I. Kartika, 2019. The immunogenicity of flagellin protein of Salmonella typhi of Semarang and Salatiga isolates. Third Int. Seminar Global Health, 3: 367-374.
Direct Link - Britto, C.D., V.K. Wong, G. Dougan and A.J. Pollard, 2018. A systematic review of antimicrobial resistance in Salmonella enterica serovar Typhi, the etiological agent of typhoid. PLoS Negl. Trop. Dis., Vol. 12.
CrossRefDirect Link - Wang, M., I.H. Qazi, L. Wang, G. Zhou and H. Han, 2020. Salmonella virulence and immune escape. Microorganisms, Vol. 8.
CrossRefDirect Link - Andrews, J.R., F.N. Qamar, R.C. Charles and E.T. Ryan, 2018. Extensively drug-resistant typhoid-Are conjugate vaccines arriving just in time? N. Engl. J. Med., 379: 1493-1495.
CrossRefDirect Link - Aziz, S. and L. Malik, 2018. Emergence of multi-resistant enteric infection in a paediatric unit of Karachi, Pakistan. J. Pak. Med. Assoc., 68: 1848-1850.
Direct Link - Salama, R.I. and N.M. Said, 2019. A comparative study of the typhidot (Dot-EIA) versus widal test in diagnosis of typhoid fever among Egyptian patients. Open J. Gastroenterol., 9: 91-98.
CrossRefDirect Link - Kumar, M., G. Velvizhi, G. Sucilathangam and C. Revathy, 2019. A comparative study of blood culture, widal test and immunochromatographic assay for rapid diagnosis of typhoid fever in a tertiary care centre. Int. J. Curr. Microbiol. Appl. Sci., 8: 500-508.
CrossRefDirect Link - Mehmood, K., A. Sundus, I.H. Naqvi, M.F. Ibrahim, O. Siddique and N.F. Ibrahim, 2015. Typhidot-A blessing or a menace. Pak. J. Med. Sci., 31: 439-443.
CrossRefDirect Link - Bundalian, Jr. R., M. Valenzuela and R.E. Tiongco, 2019. Achieving accurate laboratory diagnosis of typhoid fever: A review and meta-analysis of TUBEX® TF clinical performance. Pathog. Global Health, 113: 297-308.
CrossRefDirect Link - Andrews, J.R., F. Khanam, N. Rahman, M. Hossain and I.I. Bogoch et al., 2019. Plasma immunoglobulin a responses against 2 Salmonella typhi antigens identify patients with typhoid fever. Clin. Infect. Dis., 68: 949-955.
CrossRefDirect Link - Horstmann, J.A., M. Lunelli, H. Cazzola, J. Heidemann and C. Kühne et al., 2020. Methylation of Salmonella typhimurium flagella promotes bacterial adhesion and host cell invasion. Nat. Commun., Vol. 11.
CrossRefDirect Link - Hajam, I.A.,P.A Dar, I. Shahnawaz, J.C. Jaume and J.H. Lee, 2017. Bacterial flagellin-A potent immunomodulatory agent. Exp. Mol. Med., Vol. 49.
CrossRefDirect Link - Das, J.K., R. Hasan, A. Zafar, I. Ahmed and A. Ikram et al., 2018. Trends, associations, and antimicrobial resistance of Salmonella typhi and paratyphi in Pakistan. Am. J. Trop. Med. Hyg., 99: 48-54.
CrossRefDirect Link - Islam, K., M.A. Sayeed, E. Hossen, F. Khanam and R.C. Charles et al., 2016. Comparison of the performance of the tptest, tubex, typhidot and widal immunodiagnostic assays and blood cultures in detecting patients with typhoid fever in Bangladesh, including using a Bayesian latent class modeling approach. PLoS. Negl. Trop. Dis., Vol. 10.
CrossRefDirect Link - Gibani, M.M., C. Britto and A.J. Pollard, 2018. Typhoid and paratyphoid fever. Curr. Opin. Infect. Dis., 31: 440-448.
CrossRefDirect Link - Senevirathne, A., C. Hewawaduge and J.H. Lee, 2020. Salmonella enterica serovar enteritidis ghosts displaying a surface FliC adjuvant elicit a robust immune response and effective protection against virulent challenge. Vet. Microbiol., Vol. 243.
CrossRefDirect Link - Hatta, M., A.R. Sultan, R. Pastoor and H.L. Smits, 2011. New flagellin gene for Salmonella enterica serovar typhi from the East Indonesian archipelago. Am. J. Trop. Med. Hyg., 84: 429-434.
CrossRefDirect Link - Hajam, I.A., J.H. Kim and J.H. Lee, 2018. Incorporation of membrane-anchored flagellin into Salmonella Gallinarum bacterial ghosts induces early immune responses and protection against fowl typhoid in young layer chickens. Vet. Immunol. Immunopathol., 199: 61-69.
CrossRefDirect Link - Das, C., C. Mokashi, S.S. Mande and S. Saini, 2018. Dynamics and control of flagella assembly in Salmonella typhimurium. Front. Cell. Infect. Microbiol., Vol. 8.
CrossRefDirect Link - Eom, J.S., J.S. Kim, J.I. Jang, B.H. Kim and S.Y. Yoo et al., 2013. Enhancement of host immune responses by oral vaccination to Salmonella enterica serovar typhimurium harboring both FliC and FljB flagella. PLoS ONE, Vol. 8.
CrossRefDirect Link - Johnson, S., Y.H. Fong, J.C. Deme, E.J. Furlong, L. Kuhlen and S.M. Lea, 2020. Symmetry mismatch in the MS-ring of the bacterial flagellar rotor explains the structural coordination of secretion and rotation. Nat. Microbiol., 5: 966-975.
CrossRefDirect Link - Fukumura, T., F. Makino, T. Dietsche, M. Kinoshita and T. Kato et al., 2017. Assembly and stoichiometry of the core structure of the bacterial flagellar type III export gate complex. PLoS Biol., Vol. 15.
CrossRefDirect Link - Minamino, T., 2018. Hierarchical protein export mechanism of the bacterial flagellar type III protein export apparatus. FEMS Microbiol. Lett., Vol. 365.
CrossRefDirect Link - Song, W.S., Y.J. Jeon, B. Namgung, M. Hong and S.I. Yoon, 2017. A conserved TLR5 binding and activation hot spot on flagellin. Sci. Rep., Vol. 7.
CrossRefDirect Link - Yamaguchi, T., S. Toma, N. Terahara, T. Miyata and M. Ashihara et al., 2020. Structural and functional comparison of Salmonella flagellar filaments composed of FljB and FliC. Biomolecules, Vol. 10.
CrossRefDirect Link - Côté-Cyr, M., L. Gauthier, X. Zottig, S. Bourgault and D. Archambault. 2022. Recombinant Bacillus subtilis flagellin Hag is a potent immunostimulant with reduced proinflammatory properties compared to Salmonella enterica serovar Typhimurium FljB. Vaccine, 40: 11-17.
CrossRefDirect Link - Barkhordari, M., M. Bagheri, S. Irian, M.H. Khani, M.M. Ebrahimi, A. Zahmatkesh and S. Shahsavandi, 2021. Comparison of flagellin and an oil-emulsion adjuvant in inactivated newcastle disease vaccine in stimulation of immunogenic parameters. Comp. Immunol. Microbiol. Infect. Dis., Vol. 75.
CrossRefDirect Link - Vijayan, A., M. Rumbo, C. Carnoy and J.C. Sirard, 2018. Compartmentalized antimicrobial defenses in response to flagellin. Trends Microbiol., 26: 423-435.
CrossRefDirect Link - Mcheik, S., N.S. Al-Akl and A.M. Abdelnoor, 2018. The effect of denatured flagellin on toll-like receptor-5 (TLR-5) in mice. Endocr. Metab. Immune Disord. Drug Targets, 18: 412-416.
CrossRefDirect Link - Behzadi, P., H.A. García-Perdomo and T.M. Karpiński, 2021. Toll-like receptors: General molecular and structural biology. J. Immunol. Res., Vol. 2021.
CrossRefDirect Link - Mitchell, J., S.J. Kim, A. Seelmann, B. Veit, B. Shepard, E. Im and S.H. Rhee, 2018. Src family kinase tyrosine phosphorylates Toll-like receptor 4 to dissociate MyD88 and Mal/Tirap, suppressing LPS-induced inflammatory responses. Biochem. Pharmacol., 147: 119-127.
CrossRefDirect Link - Kuang, P., H. Guo, H. Deng, H. Cui and J. Fang et al., 2019. Sodium fluoride impairs splenic innate immunity via inactivation of TLR2/MyD88 signaling pathway in mice. Chemosphere, Vol. 237.
CrossRefDirect Link - Zhang, J., S. Pei, Y. Ding, Y. Peng, J. Chen, H. Wang and H. Wang, 2020. IRAK-M down-regulates the activation of microglial NLRP3 inflammasome and GSDMD-mediated pyroptosis in microglial through inhibiting IRAK1/TRAF6 combination during experimental autoimmune encephalomyelitis. J. Immunol., Vol. 204.
CrossRefDirect Link - Parween, F., J. Yadav and A. Qadri, 2019. The virulence polysaccharide of Salmonella typhi suppresses activation of rho family GTPases to limit inflammatory responses from epithelial cells. Front. Cell. Infect. Microbiol., Vol. 9.
CrossRefDirect Link - Al-Sadi, R., S. Guo, D. Ye, M. Rawat and T.Y. Ma, 2016. TNF-α modulation of intestinal tight junction permeability is mediated by NIK/IKK-α axis activation of the canonical NF-κB pathway. Am. J. Pathol., 186: 1151-1165.
CrossRefDirect Link - Sun, S.C., 2017. The non-canonical NF-κB pathway in immunity and inflammation. Nat. Rev. Immunol., 17: 545-558.
CrossRefDirect Link - Khani, M.H., M. Bagheri, A. Zahmatkesh and S.M. Bidhendi, 2019. Immunostimulatory effects of truncated and full-length flagellin recombinant proteins. Microb. Pathogen., 127: 190-197.
CrossRefDirect Link - Stacey, M., D. Talbot, J. Buchanan and D. Mayer, 2020. The development of an Australian teacher performance assessment: Lessons from the international literature. Asia-Pac. J. Teach. Educ., 48: 508-519.
CrossRefDirect Link - Park, D., M. Lara-Tejero, M.N. Waxham, W. Li, B. Hu, J.E. Galán and J. Liu, 2018. Visualization of the type III secretion mediated Salmonella-host cell interface using cryo-electron tomography. BioRxiv.
CrossRefDirect Link - Bergeron, J.R., 2016. Structural modeling of the flagellum MS ring protein FliF reveals similarities to the type III secretion system and sporulation complex. PeerJ, Vol. 4.
CrossRefDirect Link - Huang, F.C., 2021. The interleukins orchestrate mucosal immune responses to Salmonella infection in the intestine. Cells, Vol. 10.
CrossRefDirect Link - Senevirathne, A., C. Hewawaduge and J.H. Lee, 2021. Immunization of chicken with flagellin adjuvanted Salmonella enteritidis bacterial ghosts confers complete protection against chicken salmonellosis. Poult. Sci., Vol. 100.
CrossRefDirect Link - Yang, S., Q. Deng, L. Sun, Y. Zhu and K. Dong et al., 2021. Salmonella effector SpvB inhibits NF-κB activity via KEAP1-mediated downregulation of IKKβ. Front. Cell. Infect. Microbiol., Vol. 11.
CrossRefDirect Link