
Mat Ghani Hilmi
Institute of Tropical Aquaculture and Fisheries, Universiti Malaysia Terengganu, Kuala Nerus, 21030 Terengganu, Malaysia
Mhd Ikhwanuddin
Institute of Tropical Aquaculture and Fisheries, Universiti Malaysia Terengganu, Kuala Nerus, 21030 Terengganu, Malaysia
LiveDNA: 60.4241
Pakistan Journal of Biological Sciences
Year: 2020 | Volume: 23 | Issue: 5 | Page No.: 708-714







ABSTRACT
Background and Objective: The accumulation of lipofuscin (LF) is an alternative technique to identify age of crustacean species. However, the exact sites and the level of the LF concentration were unknown especially for different sexes of blue swimming crab, Portunus pelagicus. Thus, the present study was aimed to identify which part of the eyestalk of P. pelagicus contains more LF levels in order to establish a specific target part of samples. Materials and Methods: Thus, crab samples for this study were sampled from the wild habitat at Setiu wetlands, Terengganu, Malaysia. A total of 100 samples of with the same size (80±5 mm carapace width) were sampled and the eyestalk dissected for LF extraction. The determination of LF sites and levels in the eyestalks organ was taken from the area between the distal tangential layer (DTL) and proximal tangential layer (PTL). The lower part of the eyestalk was taken from the PTL until the end of the eyestalk. Results: The upper part of the crab’s eyestalk was higher in the males crabs compared to the females crabs. LF index also shown that the upper part of crab’s eyestalk have higher concentration compared to the lower part. Conclusion: The left crab’s eyestalk had the higher LF index especially in males compared to females but the total concentration was higher in female crabs. Knowing which part has highly dense accumulation of LF helps in LF detection of the tissue and further helps for age determination for this species.
PDF Abstract XML References Citation
Copyright: © 2020. This is an open access article distributed under the terms of the creative commons attribution License, which permits unrestricted use, distribution and reproduction in any medium, provided the original author and source are credited.
How to cite this article
Mat Ghani Hilmi and Mhd Ikhwanuddin, 2020. Lipofuscin Level in Eyestalk Blue Swimming Crab, Portunus pelagicus at Different Sexes. Pakistan Journal of Biological Sciences, 23: 708-714.
DOI: 10.3923/pjbs.2020.708.714
URL: https://scialert.net/abstract/?doi=pjbs.2020.708.714
DOI: 10.3923/pjbs.2020.708.714
URL: https://scialert.net/abstract/?doi=pjbs.2020.708.714
INTRODUCTION
The accumulation of lipofuscin (LF) in the eyestalk organ is among an alternative technique to identify age of crustacean. One of the biggest problems in the biochemical analysis of LF (aging pigments) is the lack of standardization in methodologies. Various techniques have been done in the past to determine the age of crustaceans. Another approach that involves fluorescence staining and spectrophotometry was used to associate LF with crab age1. Lipofuscin (LF) is formed during lipid peroxidation and sugar glycosylation by carbonyl-amino crosslinks with biomacrolecules and accumulates slowly within postmitotic cells2. LF is comprised of lipids (30-70%), proteins (20-50%), carbohydrates (4-7%) and metals, primarily iron and copper3. Few researchers compared histology sections of crab tissue with varying LF concentrations for age identification4. LF is characterized by three main properties: (1) it is contained within intracellular lysosomal bodies, (2) it has yellow autofluorescent emission when excited with ultraviolet or blue light and (3) it accumulates in postmitotic tissues with age5. These properties, along with its unique biochemical and morphological characteristics, have led researchers to consider LF as a “hallmark in aging”. Because concentration has been shown to increase with increasing age in animals, it has served as an age biomarker in many invertebrate aging studies where morphological measurements do not correlate to age6.
The complex chemical structure of LF allow it to auto-fluorescence but, peak emission wavelengths vary in regards to species, tissue types and concentrations7. Currently, this technique is applied to identify age of many other crustaceans such as European lobster, Homarus gammarus8, tiger prawn, Penaus monodon9 and the crayfish species, Cherax quadricarinatus and Pacifastacus leniusculus10,11. It is important that a standard be created for replication and verification of results. This becomes especially important when analysis is used in fishery management for a species with a significant economic and ecological value. Furthermore, most of the studies on LF so far have focused on the brain and the olfactory lobe globuli cell masses (OLCM) where LF are likely obvious. Sheehy et al.12 stated that if LF accumulation correlates with functionality or regionalizations of metabolic activity within tissues, it could be that patterns of accumulation in the eyestalk ganglia are quite different from those in the olfactory lobe of the brain. The use of eyestalk ganglia is more practical then the brain because it makes sample collection easier and keeps the samples alive.
The blue swimming crabs, Portunus pelagicus is among the most important species for further climate change and pre-commercialization studies13-20. Previous studies by some authors have focused on the comparison of LF accumulation on the right and left side of the eyestalks. Ju et al.21 and Puckett et al.22 looked at on blue crab, Callinectes sapidus while Maxwell et al.6 the spiny lobster, Panulirus argus. All those studies have the same conclusion that right and left eyestalks contain statistically similar concentrations of extractable age pigments. However, no study has been done on P. pelagicus and this species may have different concentration of LI between right and left eyestalks.
This study aimed to identify which part of the eyestalk of P. pelagicus contains more LF levels in order to establish a specific target part of samples. The difference of LF accumulation in the right and left eyestalks was also observed to confirm and provide a basis of LF levels data for future researchers.
MATERIALS AND METHODS
Sampling technique of the population: Crab samples for this study were sampled from the wild habitat at Setiu wetlands, Terengganu, Peninsular Malaysia (Fig. 1). The samples of P. pelagicus at Setiu Wetlands areas were sampled from series of bio-sampling trips using two method using gill net and second is hunting during the night23-28. The study was carried out at Crustacean Hatchery, Institute of Tropical Aquaculture and Fisheries laboratory from January, 2015-March, 2018.
A total of 100 samples of each sex with the same size (80 mm CW) were sampled and the eyestalk dissected for LF extraction. After obtaining CW and BW measurements, the crabs were anesthetised in an ice bath, the eyestalks removed and tissue from the external portion of the eyestalk was excised from the structural chitin. All the samples chosen were active crabs without any physical damaged. Dissection was done carefully in order to keep the lower part of the eyestalk in a good condition. The eyestalk of the samples was divided into two parts, the upper part and lower part (Fig. 2). The segmentation of the part was based on neuron morphologies and eyestalk anatomy previously studied by Sztarker et al.29 on bayshore crab, Hemigrapsus oregonensis.
 | |
| Fig. 1: | Present study sampling site of Setiu wetlands, Terengganu, Malaysia |
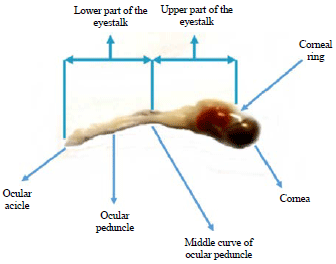 | |
| Fig. 2: | Segmentation of Portunus pelagicus eyestalk |
The upper part of the eyestalk was taken from the area between the distal tangential layer (DTL) and proximal tangential layer (PTL). The lower part of the eye stalk was taken from the PTL until the end of the eyestalk.
Experimental design and lipofuscin extraction: Overall, the samples taken from wild habitat were measured for their biological features such as CW, BW and sexes. Twenty five pairs of samples then performed with lipofuscin analysis for differentiation of ‘right and left’ and ‘upper part and lower part’ experiments. As for the comparison of LI between right and left eyestalk, 50 eyestalk samples from another 25 pairs of both right and left eyestalk with random size for each sex were sampled. The conditions of the samples were perfect and without physical damage. LF extractions of all crab specimens from wild and cage were carried out following the protocol adapted from Ju et al.30.
Retinal tissue from the eye were excluded, to overlapping of fluorescence readings (excitation range of 325-340 nm and emission of 475 nm) with the LF compounds fluorescence (excitation of 340 nm and emission of 405 nm). Eyestalk tissues were placed in a 4 mL amber vial containing 2 mL of a 2:1 dichloromethane-methanol mixture (CH2Cl2-MeOH). Vials were set on ice and sonicated at 20 watts for 2 min to extract LF pigment from the solvent. This followed by high power centrifugation for 10 min. All the supernatant was then transferred by pipetting to a clean 4 mL amber vial. The supernatant were dried completely under pressurized nitrogen gas. Upon completion of drying, the crab LF pellets were re-dissolved in 4 mL of methanol (MeOH) and vortexed, followed by removal of 1 mL for protein analysis. The remaining 3 mL samples were measured (max excitation 340 and max emission 405) using a Cary eclipse routine fluorescence spectrophotometer.
Calibration of fluorescence readings were carried out using quinine sulphate (quinine sulfate dissolved in 0.1 N sulfuric acid) at concentrations of 0-0.3 μg mg–1. In this method, 1 mL of sample was dried using N2, as protocol to remove the remaining methanol. Protein concentration was assayed in order to normalize the fluorescence reading to protein concentration of the extracted sample. Individually, all samples were prepared for analysis using bicinchoninic acid protein assay kit (BCA), from concentrations of 0-400 μg mL–1 (400, 200,100, 50, 25,12.5 and 0 μg L–1). The dried proteins were re-dissolved in 200 μL of DI water. As much as 100 μL sample were added with 2 mL BCA Working Reagent and vortexed at medium speed for two seconds. All samples were incubated at 60°C for 15 min, before cooling at room temperature. The absorbance of standards and samples were measured at a wavelength of 562 nm using the Thermo Scientific UV-1800-240 V spectrophotometer.
Statistical analysis: A paired-samples student t-test was used for statistical analyses of different eyestalk segmentation studies. Fisher’s r to z transformation was done for the right and left eyestalks comparison so that the z-score can be compared and analyzed for statistical significance by determining the observed z-test statistic. For all statistical analysis, α = 0.05.
RESULTS
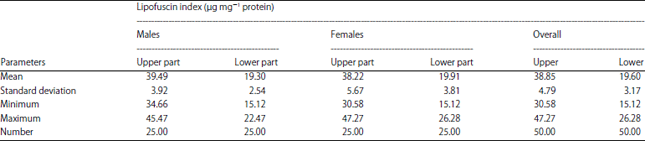
The upper part of male’s eyestalk showed a mean of 39.49±3.92 μg mg–1 (protein) and 19.3±2.54 μg mg–1 (protein) for lower part (Table 1). The maximum (Max) and minimum (Min) of LI was 45.57 and 34.66 μg mg–1 (protein) respectively for upper part and 22.47 and 15.12 μg mg–1 (protein) for lower part. As for females, the upper parts shows a mean 38.22±5.67 22 μg mg–1 (protein) and mean 19.91±3.81 μg mg–1 (protein) for lower part (Table 1). The maximum and minimum of LI was 47.27 and 30.58 μg mg–1 (protein) respectively for upper part and 26.28 and 15.12 μg mg–1 (protein) for lower part.
Paired-samples student-t test analysis showed that the LI in the upper part were significantly higher than the LI in the lower part of males eyestalks (p<0.001). All samples showed the LF in upper part were 100% or above higher compare to lower part. There was not even one sample showed LF in lower part was higher than upper part. Paired-samples student t-test analysis for female’s eyestalk showed that the LI in the upper part were also significantly higher than the LI in the lower part (p<0.001). A comparison of LI between upper and lower part were shown in Fig. 1. All samples showed at least 50% difference of LI between lower part and upper part. As for sexes combined result, the upper part of eyestalk shows a mean 38.85±4.79 μg mg–1 (protein) and mean 19.60±3.17 μg mg–1 (protein) for lower part (Table 1). The maximum and minimum of LI was 47.27 and 30.58 μg mg–1 (protein) respectively for upper part and 26.28 and 15.12 μg mg–1 (protein) for lower part.
As for the comparison between right and left eyestalks for males, the correlation coefficient of LI was not significantly different at p = 0.05. The z observed was less than critical value and the probability value was greater than 0.05. The correlation analysis has showed that there are no differences at all between the LI of left and right eyestalks of males P. pelagicus. The right eyestalks LI shows a mean 19.86±10.48 μg mg–1 (protein) and mean 24.12±10.67 μg mg–1 (protein) for left eyestalks (Table 2). A coefficient of LI for females was also not significantly different at p = 0.05. The z observed was less than critical value and the probability value was greater than 0.05. The correlation analysis has showed that there are no differences at all between the LI of left and right eyestalks of female’s P. pelagicus. The right eyestalks LI shows a mean 22.48 μg mg–1 (protein) ±10.75 and mean 22.52±9.20 μg mg–1 (protein) for left eyestalks (Table 2).
The relationship of LI and CW for right and left males was LI = 9.1922CW-46.126, R2 = 0.8935 and LI = 8.0812CW-36.981, R2 = 0.7595, respectively (Fig. 3a). As for females, the relationship of LI and CW for right and left was LI = 6.0038, CW-25.488, R2 = 0.6643 and LI = 5.5653CW-21.939, R2 = 0.672, respectively (Fig. 3b).
| Table 1: | Overall mean, standard deviation, minimum and maximum, of LI of males and females Portunus pelagicus from Setiu wetlands, Terengganu, Malaysia |
 | |
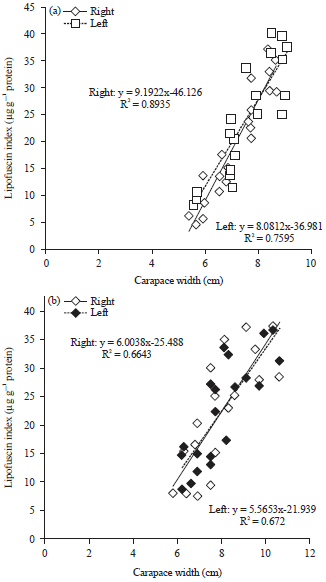 | |
| Fig. 3(a-b): | A linear regression of LI and carapace width of (a) Males and (b) Females Portunus pelagicus from Setiu wetlands, Terengganu, Malaysia |
| Table 2: | Overall mean, standard deviation, minimum and maximum, of LI of right and left eyestalks of males and females Portunus pelagicus from Setiu wetlands, Terengganu, Malaysia |
 | |
DISCUSSION
This study has also showed that there are no significant differences between LF accumulation in both right and left eyestalks. The slope of linear regression line of LI and CW relationship for left and right eyestalk was not significantly different for both sexes. This study has validated the findings of previous studies by Maxwell et al.6, Ju et al.21 and Puckett et al.22. Ju et al.21 stated that extractable LF content between left and right eye-stalks of individual crabs were significantly correlated (r = 0.86, p<0.005, n = 10). Maxwell et al.6, their study showed LF concentrations of replicate left and right eye samples from Dry Tortugas lobsters were not different (t = 2.29, df = 105, p>0.05). All of these results show that mostly crustacean having similar LF accumulation in left and right eyestalks and P. pelagicus also included.
Sheehy et al.12 states that LF accumulation correlates with functionality or regionalization of metabolic activity within tissues, thus the LF in the left and right eyestalks was having the same concentration of LF since both left and right eyestalks are relatively having same functionality and same regions. But in other cases, after comparing the concentration of LF on different part of eyestalk, they are significantly different. This showed that even the samples taken from the same organs or regions, but the different part of the organs may contain different massive of tissue. In this study case, the neuron tissue from the lower part of the eyestalk seems to be lower than the upper part. That is why the concentration of the LF in the upper part was higher compared to the lower part of eyestalks. This is the first study operated on differences of LF in the different part of eyestalks, thus no other finding from previous study that can be compared to. Based on the result, the maximum value of LI for lower part not even reaches the minimum value of upper part. There is explanation on LI occur higher at the upper part compared to lower part. The upper part that is close to the retina contains more LF compared to lower part of the eyestalk. Based on neuronal tissue image from Sztarker et al.29 study, the neuronal tissue was more densely accumulated in the upper part of eyestalk compared to the lower part. The variation of LI in individual samples shows that these crabs with similar sizes are not from the same age. However, age differences do not affect the pattern of high LI in the upper compared to lower part of the eyestalks.
This study has demonstrated the specific site of LF accumulation in the eyestalks of P. pelagicus in order to help future researchers know which part is the best for LF extraction. This study has concluded that there is significant difference of LF accumulation in the upper part and lower part of the eyestalk. This result has indicated that the best part for LF extraction is from the upper part of the eyestalk. Knowing which part has highly dense accumulation of LF helps in LF detection of the tissue and is more accurately and easily detected by any methods used. Knowing the similarity of LF accumulation between right and left eyestalks helps in conserving the samples for replication. Both right and left eyestalks can be used for LF extraction comparison in different states of the samples for crustaceans including P. pelagicus. There are no arguments since this data on crustaceans has been previously validated by several authors as well as from this latest study.
CONCLUSION
This study have successfully identified that both right and left eyestalks of P. pelagicus contains LF extraction for further age determination analysis however, the male left eyestalk have higher LF index compared to female eyestalk.
SIGNIFICANCE STATEMENT
This study discovers the LF extraction of P. pelagicus eyestalk which can be beneficial for further age determination analysis. This study will help the researcher to uncover the critical areas of related-age determination studies that many researchers were not able to explore. Thus a new theory on LF detection of the tissue of other crustacean may be arrived at.
REFERENCES
- Kilada, R. and J.G. Driscoll, 2017. Age determination in crustaceans: A review. Hydrobiologia, 799: 21-36.
CrossRefDirect Link - Cai, S., H. Yang, K. Zeng, J. Zhang and N. Zhong et al., 2016. EGCG inhibited lipofuscin formation based on intercepting amyloidogenic β-sheet-rich structure conversion. PLoS ONE, Vol. 11, No. 3.
CrossRefDirect Link - Brunk, U.T. and A. Terman, 2002. Lipofuscin: Mechanisms of age-related accumulation and influence on cell function. Free. Radic. Biol. Med., 33: 611-619.
CrossRefDirect Link - Sheehy, M.R.J., 2008. Questioning the use of biochemical extraction to measure lipofuscin for age determination of crabs: Comment on Ju et al. (1999, 2001). Mar. Ecol. Progr. Ser., 353: 303-306.
CrossRefDirect Link - Katz, M.L. and W.G. Robison Jr., 2002. What is lipofuscin? Defining characteristics and differentiation from other autofluorescent lysosomal storage bodies. Arch. Gerontol. Geriatr., 34: 169-184.
CrossRefDirect Link - Maxwell, K.E., T.R. Matthews, M.R.J. Sheehy, R.D. Bertelsen and C.D. Derby, 2007. Neurolipofuscin is a measure of age in Panulirus argus, the Caribbean spiny lobster, in Florida. Biol. Bull., 213: 55-66.
CrossRefDirect Link - Li, G., Y. Liao, X. Wang, S. Sheng and D. Yin, 2006. In situ estimation of the entire color and spectra of age pigment-like materials: Application of a front-surface 3D-fluorescence technique. Exp. Gerontol., 41: 328-336.
CrossRefDirect Link - Sheehy, M.R.J. and R.C. Bannister, 2002. Year-class detection reveals climatic modulation of settlement strength in the European lobster, Homarus gammarus. Can. J. Fish. Aquat. Sci., 59: 1132-1143.
CrossRefDirect Link - Sheehy, M.R.J., E. Cameron, G. Marsden and J. McGrath, 1995. Age structure of female giant tiger prawns Penaeus monodon as indicated by neuronal lipofuscin concentration. Mar. Ecol. Progr. Ser., 117: 59-63.
CrossRefDirect Link - Belchier, M., L. Edsman, M.R.J. Sheehy and P.M. Shelton, 1998. Estimating age and growth in long-lived temperate freshwater crayfish using lipofuscin. Freshwater Biol., 39: 439-446.
CrossRefDirect Link - Fonseca, D.B., M.R.J. Sheehy and P.M.J. Shelton, 2003. Unilateral eyestalk ablation reduces neurolipofuscin accumulation rate in the contralateral eyestalk of a crustacean, Pacifastacus leniusculus. J. Exp. Mar. Biol. Ecol., 289: 277-286.
CrossRefDirect Link - Sheehy, M.R.J., P.M.J. Shelton, J.F. Wickins, M. Belchier and E. Gaten, 1996. Ageing the European lobster Homarus gammarus by the lipofuscin in its eyestalk ganglia. Mar. Ecol. Prog. Ser., 143: 99-111.
CrossRefDirect Link - Azra, M.N., J.C. Chen, T.H. Hsu, M. Ikhwanuddin and A.B. Abol-Munafi, 2019. Growth, molting duration and carapace hardness of blue swimming crab, Portunus pelagicus, instars at different water temperatures. Aquacult. Rep., Vol. 15.
CrossRefDirect Link - Azra, M.N., M.A. Aaqillah-Amr, M. Ikhwanuddin, H. Ma and K. Waiho et al., 2020. Effects of climate-induced water temperature changes on the life history of brachyuran crabs. Rev. Aquacult., 12: 1211-1216.
CrossRefDirect Link - Azra, M.N., J.C. Chen, M. Ikhwanuddin and A.B. Abol-Munafi, 2018. Thermal tolerance and locomotor activity of blue swimmer crab Portunus pelagicus instar reared at different temperatures. J. Thermal Biol., 74: 234-240.
CrossRefDirect Link - Ikhwanuddin, M., A.B. Abol-Munafi and M.N. Azra, 2019. Data on the molting duration and time of hardening of instar crab at different culture temperatures. Data Brief, Vol. 25.
CrossRefDirect Link - Ikhwanuddin, M.H.D., M.N. Azra, N.F. Noorulhudha, A. Siti-Aishah and A.B. Abol-Munafi, 2016. Embryonic development and hatching rate of blue swimming crab, Portunus pelagicus (Linnaeus, 1758) under different water salinities. Turk. J. Fish. Aquat. Sci., 16: 669-677.
CrossRefDirect Link - Abol-Munafi, A.B., N. Pilus, R.M. Amin, M.N. Azra and M. Ikhwanuddin, 2017. Digestive enzyme profiles from foregut contents of blue swimming crab, Portunus pelagicus from Straits of Johor, Malaysia. J. Assoc. Arab Univ. Basic Applied Sci., 24: 120-125.
CrossRefDirect Link - Abol-Munafi, A.B., M.S. Mukrim, R.M. Amin, M.N. Azra, G. Azmie and M. Ikhwanuddin, 2016. Histological profile and fatty acid composition in hepatopancreas of blue swimming crab, Portunus pelagicus (Linnaeus, 1758) at different ovarian maturation stages. Turkish J. Fish. Aquat. Sci., 16: 251-258.
CrossRefDirect Link - Taufik, M., Z. Bachok, M.N. Azra and M. Ikhwanuddin, 2016. Effects of various microalgae on fatty acid composition and survival rate of the blue swimming crab Portunus pelagicus larvae. Indian J. Geo-Mar. Sci., 45: 1512-1521.
Direct Link - Ju, S.J., D.H. Secor and H.R. Harvey, 2003. Demographic assessment of the blue crab (Callinectes sapidus) in Chesapeake Bay using extractable lipofuscins as age markers. Fish. Bull., 101: 312-320.
Direct Link - Puckett, B.J., D.H. Secor and S.J. Ju, 2008. Validation and application of lipofuscin-based age determination for Chesapeake Bay blue crabs Callinectes sapidus. Trans. Am. Fish. Soc., 137: 1637-1649.
CrossRefDirect Link - Azra, M.N. and M. Ikhwanuddin, 2015. Larval culture and rearing techniques of commercially important crab, Portunus pelagicus (Linnaeus, 1758): Present status and future prospects. Songklanakarin J. Sci. Technol., 37: 135-145.
Direct Link - Ikhwanuddin, M., M.N. Azra, H. Siti-Aimuni and A.B. Abol-Munafi, 2012. Fecundity, embryonic and ovarian development of blue swimming crab, Portunus pelagicus (Linnaeus, 1758) in Coastal Water of Johor, Malaysia. Pak. J. Biol. Sci., 15: 720-728.
CrossRefDirect Link - Ikhwanuddin, M., A.N. Liyana, M.N. Azra, Z. Bachok and A.B. Abol-Munafi, 2014. Natural diet of blue swimming crab, Portunus pelagicus at strait of Tebrau, Johor, Malaysia. Sains Malaysiana, 43: 37-44.
Direct Link - Ikhwanuddin, M., M.N. Azra, M.A.D. Talpur, A.B. Abol-Munafi and M.L. Shabdin, 2012. Optimal water temperature and salinity for production of blue swimming crab, Portunus pelagicus 1st day juvenile crab. AACL Bioflux, 5: 4-8.
Direct Link - Ikhwanuddin, M., S. Mohamed, A.I. Abdul Rahim, M.N. Azra, S.A. Jaaman, A.M.A. Bolong and N.M. Noordin, 2015. Observations on the effect of natural diets on ovarian rematuration in blue swimming crab Portunus pelagicus (Linnaeus, 1758). Indian J. Fish., 62: 124-127.
Direct Link - Ikhwanuddin, M., M.N. Azra, A. Redzuari, Z.A. Aizam and A.B. Abol-Munafi, 2012. Ingestion rates of Brachionus sp. and Artemia sp. nauplii by blue swimming crab, Portunus pelagicus (Linnaeus, 1758) larvae. J. Fish. Aquat. Sci., 7: 402-411.
CrossRefDirect Link - Sztarker, J., N. Strausfeld, D. Andrew and D. Tomsic, 2009. Neural organization of first optic neuropils in the littoral crab Hemigrapsus oregonensis and the semiterrestrial species Chasmagnathus granulatus. J. Comp. Neurol., 513: 129-150.
CrossRefDirect Link - Ju, S.J., D.H. Secor and H.R. Harvey, 1999. Use of extractable lipofuscin for age determination of blue crab Callinectes sapidus. Mar. Ecol. Progr. Ser., 185: 171-179.
CrossRefDirect Link