
Mat Ghani Hilmi
Institute of Tropical Aquaculture and Fisheries, Universiti Malaysia Terengganu, 21030 Kuala Nerus, Terengganu, Malaysia
Mhd Ikhwanuddin
Institute of Tropical Aquaculture and Fisheries, Universiti Malaysia Terengganu, 21030 Kuala Nerus, Terengganu, Malaysia
LiveDNA: 60.4241
Pakistan Journal of Biological Sciences
Year: 2020 | Volume: 23 | Issue: 5 | Page No.: 685-690







ABSTRACT
Background and Objective: Fundamental information on how environmental conditions affect lipofuscin (LF) levels could be useful to those furthering the development of an effective management plan to control certain populations in certain geographical areas. Thus, the effects of salinity upon LF levels were examined using 7 salinities in the range 10-40% (increasing in steps of 5%). Materials and Methods: The impacts of freezing preservation were also examined for 60 crab samples with carapace widths of 7-8 cm randomly allocated for either immediate or post-freezing (-80°C) analysis. Results: The result showed that 25% salinity was optimum for LF accumulation in both males and females crabs. The mean value of LF production was higher for males compared to females. Higher and lower salinity of 10 and 40% were the most stressed conditions for LF accumulation. The LF index (LI) of P. pelagicus decreased in salinities below and above the typical range. Conclusion: For both sexes, the frozen samples exhibited notably higher LI values than the fresh samples (p<0.05). These results will be applicable in numerous respects, for example in the selection of farm locations and for optimising productivity by controlling salinity levels.
PDF Abstract XML References Citation
Copyright: © 2020. This is an open access article distributed under the terms of the creative commons attribution License, which permits unrestricted use, distribution and reproduction in any medium, provided the original author and source are credited.
How to cite this article
Mat Ghani Hilmi and Mhd Ikhwanuddin, 2020. Salinity and Preservation Conditions of Lipofuscin Extracts in Blue Swimming Crab, Portunus pelagicus. Pakistan Journal of Biological Sciences, 23: 685-690.
DOI: 10.3923/pjbs.2020.685.690
URL: https://scialert.net/abstract/?doi=pjbs.2020.685.690
DOI: 10.3923/pjbs.2020.685.690
URL: https://scialert.net/abstract/?doi=pjbs.2020.685.690
INTRODUCTION
The LF analysis has been introduced as one of the most accurate age determination methods for crustaceans. However, since salinity has been shown to affect LF accumulation, further study into the role of salinity in LF accumulation is warranted.
Over the past 30 years, new methods for determining age in crustaceans have been developed1. These include size-frequency analysis2, counting growth bands3,4, evaluating molting activities5 and determining LF levels in the eyestalk1. The latter is known to be one of the best markers of age in crustaceans6.
The pigment LF, produces during cellular metabolism in crustacean6, has been quantitatively studied to generate findings on age determination7 in lobsters8, crayfish9, prawns10 and krill11. The blue swimming crab (Portunus pelagicus) is among the most economically significant seafood species for domestic consumption and exportation, as well as for culture and research12-20. Due to its increasing price in Malaysia, many coastal fishermen are exploiting this species as a food source and for use in produce21,22. This has led to several studies being carried out on diverse topics such as population structure23, fatty acids composition24, reproduction25 and chitin and chitosan extraction26 of the Portunus species.
Since crustacean metabolism is critically influenced by salinity27, it may also influence LF levels. Previous studies of P. pelagicus28 and P. trituberculatus29 have shown that crab metabolism is affected by changes in salinity. Generally, estuarine invertebrates are able to tolerate or control variations in ambient salinity, however, few studies dealing with the effects of salinity upon LF have been published.
Moreover, few studies have examined the impact of freezing preservation upon LF levels. To avoid possible effects of chemicals or freezing, Ju et al.6 examined fresh samples only, while Puckett et al.30 opted to freeze samples at -80°C, so there was no notable difference between fresh and frozen crab flesh.
The present study therefore examined the effects of salinity and preservation process upon LF levels in the eyestalks of P. pelagicus.
MATERIALS AND METHODS
Animals preparations and water management: Gill net was used to collect wild P. pelagicus samples from Setiu Wetlands, in the coastal waters of Terengganu, Malaysia and brought to the crustacean hatchery at the Institute of Tropical Aquaculture and Fisheries, Universiti Malaysia Terengganu, Malaysia. The study was carried out at Crustacean Hatchery, Institute of Tropical Aquaculture and Fisheries laboratory from Jan, 2015-March, 2018. Mature crabs (n = 60) with carapace widths (CW) of 8-9 cm were selected because their high LF levels would facilitate extraction and analysis31-32. At the beginning of the experiment, water was disinfected with 25 ppm of active chlorine and neutralized with 30 ppm sodium thiosulphate. A total of 0.8 t of seawater of the appropriate salinity was created using filtered seawater diluted with distilled water. Crabs were adjusted to their environment and maintained in adequately aerated circular fiberglass holding tank with twice daily feedings (noon and night) of chopped squid and Loligo spp., flesh.
Experimental design: In accordance with the species' tolerance range33, experiments were performed using seven salinities in the range 10-40%, increasing in steps of 5%. A total of 30 crab replicates of each sex for every salinity level was used for this experiment. The culture tank has a volume of 1 t with dimensions of 140 cm diameter and 80 cm height. Salinity measurements were made with a refractometer (ATC brand, model: LB32T-CU). Throughout the study trials (one month), the water temperature of 29±1°C was maintained using a water heater. It took 2-3 weeks for crabs to show significant effects of the salinity and temperature changes, which is why a one-month culture period was established for this present study.
The effects of preservation were investigated for 30 crabs of each sex (CW = 9 cm). The eyestalks were separated into two groups (left and right), the LF levels of the first group were analysed immediately after collection, while the second group was stored for two weeks at -80°C before analysis. There is no significant difference between left and right eyestalks, hence, the effects of freezing could be examined by comparing the LF levels of a pair of eyestalks (one fresh and one frozen) from each crab. The sex, CW and body weight (BW) of each crab was recorded before anesthetization in an ice bath.
Extraction process: Based on the procedure of Ju et al.6, the eyestalk was removed and tissue from its external portion was excised from the structural chitin. Retinal tissue was excluded to avoid overlap of fluorescence readings (excitation at 325-340 nm, emission at 475 nm) with those of LF (excitation at 340nm, emission at 405 nm). The eyestalk tissue was placed in a 2:1 dichloromethane-methanol mixture (2 mL) in an amber vial (4 mL capacity) and sonicated in an ice bath at 20 W for 2 min prior to centrifugation at high power for 10 min. The LF-containing supernatant was pipetted into a clean vial, completely dried under pressurized nitrogen (N2), re-dissolved in methanol (4 mL) and vortexed, after which a 1 mL aliquot was removed for protein determination. The fluorescence of the remainder was measured using a cary eclipse routine fluorescence spectrophotometer (max excitation at 340 nm, max emission 405 nm). A quantitative measure of LF was obtained using a calibration curve (quinine sulphate in 0.1 N sulfuric acid) with concentrations of 0-0.3 μg mg–1. After drying under N2 gas to remove the remaining methanol, the protein concentration was assayed to normalize the fluorescence reading to the protein concentration of the extracted sample. The protein was then assayed using a bicinchoninic acid protein assay kit (BCA, Fisher) with 0-400 μg mL–1 protein standards. After re-dissolving the dried protein sample in deionised water (200 μL), a 100 μL aliquot of sample was added to the BCA Working Reagent (2 mL) and vortexed for 2 sec at medium speed. The samples were incubated for 15 min at 60°C and then cooled to room temperature. The absorbance of standards and samples at 562 nm were measured using the Thermo Scientific UV-1800 240V spectrophotometer and the LI values were calculated. The LI was then calculated as follows:
Statistical analysis: Data sets were tested for normality with the Shapiro-Wilk test and for constant variance using Levene’s test. For data sets that met these assumptions, a one-way analysis of variance (ANOVA) was performed to test for differences in absolute LF concentration in different salinities regimes. A paired-samples student t-test was used for statistical analysis of the effects of the freezing studies. A 95% confident level was set for this analysis.
RESULTS
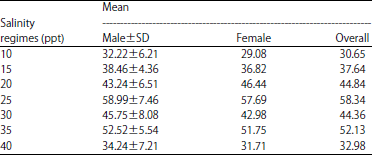
Effects of salinity: The ANOVA significance test revealed a significant variation in mean LI values with varying salinity (p<0.001). For the male P. pelagicus, a salinity of 25 ppt allowed the maximum amount of LF production with a mean of 58.99±5.01 μg mg–1 (protein) (Table 1). The worst salinity conditions for LF accumulation were 10 and 40 ppt. The mean value of LI for both salinity regimes was below 35 μg mg–1 (protein), 32.22±6.21 μg mg–1 (protein) for 10 ppt and 34.24±6.61 μg mg–1 (protein) for 40 ppt (Table 1). Female P. pelagicus also showed a significant difference of LI in different salinity regimes (p<0.001). The results showed that 25 ppt salinity is optimal for LF accumulation in females, with a mean of 57.69±7.46 μg mg–1 (protein) (Table 1). Samples from 10 and 40 ppt salinity were also the most stressed conditions, compared to other regimes. The mean value for 10 ppt and 40 ppt was very low, at 29.08±7.96 μg mg–1 (protein) and 31.71±7.21 μg mg–1 (protein), respectively (Table 1). Since the males and females showed similar LF levels in different salinity regimes, the overall results combining both sexes follow the same outcome. The optimum salinity regimes for LF levels were 25 ppt, because the value of the mean was the highest, at 58.34±6.22 μg mg–1 (protein). The 10 ppt and 40 ppt regimes had very low LF levels, 30.65±7.13 μg mg–1 (protein) and 32.98±6.86 μg mg–1 (protein), respectively (Table 1).
Effects of freezing: Paired-samples student t-test analysis between fresh and frozen male samples showed mean LI in frozen samples to be significantly higher than the mean LI in fresh samples (p<0.05). A comparison of LI means between fresh and frozen samples is shown in Table 2. Frozen eyestalk samples showed a mean of 59.35±9.28 μg mg–1 (protein) while fresh samples showed 41.62±6.49 μg mg–1 (protein) for males (Table 2). As for females, paired-samples student t-test analysis between fresh and frozen samples showed the mean LI in frozen samples was also significantly higher than mean LI in fresh samples (p<0.05). According to the paired-samples student-t test analysis, mean LI was significantly higher in frozen samples than in fresh samples for both male and female crabs. The LI values were comparable for each sex (p<0.001). For females, frozen eyestalks displayed a mean of 58.12±8.82 μg mg–1 (protein), while the fresh samples displayed 46.91±8.59 μg mg–1 (protein). For males, frozen eyestalks exhibited LI values of 58.74±8.83 μg mg–1 (protein), while fresh sample values were 44.26±7.89 μg mg–1 (protein).
| Table 1: | Mean with±standard deviation of LI in the eyestalk organ of Portunus pelagicus cultured in different salinities regimes (n = 60) |
 | |
| Table 2: | Mean with standard deviation of lipofuscin index of fresh and frozen samples for Portunus pelagicus from Setiu wetlands, Terengganu, Malaysia |
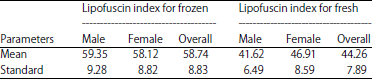 | |
DISCUSSION
This study has showed that the 25% salinity was as optimum level for LF accumulation in both males and females P. pelagicus. There was a significance different found between the fresh and frozen eyestalk samples in P. pelagicus crab. Various impacts of salinity and preservation conditions upon brachyuran crabs have been noted in previous studies. For example, Falconer et al.34, noted decreased oxygen consumption rates in 2 intertidal crab species of Hemigrapsus crenulatus and H. sexdentatus, under cyclic salinity changes relative to that of crabs maintained under constant salinity within a similar range. Since respiring organisms effectively obtain all their metabolic energy by consuming oxygen, this factor provides an indirect measure of metabolic rate. Moreover, since LF is formed during cellular metabolic processes, increased oxygen consumption rates lead to increased cellular levels of LF.
Since salinity affected the oxygen intake of P. pelagicus, LF concentration was closely related to changes in water salinity. From the results, it can be assumed that the optimal metabolic rate is 25 ppt salinity. The LI for 25 ppt salinity was the highest for males and females. This indicates the metabolic rate was at the optimal state, since optimal salinity allowed species to uptake oxygen consumption at their favored condition and produce LF without stress. This finding is different from those of other studies, as most suggest salinity of more than 30 ppt as the most favorable salinity for growth of P. pelagicus. Kangas35 and Anand and Soundarapandian36 have suggested 30-40 and 33-35 ppt salinity, respectively, as the most suitable for P. pelagicus culture. As for juvenile crabs, Romano and Zheng33 have suggested 25 ppt as the optimum salinity for the crabs to grow, which is the same for P. pelagicus in the Setiu Wetlands. P. pelagicus seems to have adapted well to the salinity of the Setiu Wetlands.
The average monthly salinity at the Setiu Wetlands is a low 20-30 ppt throughout the year due to the extensive input of fresh river water37,38. Extreme low (10 ppt) or high (40 ppt) salinity conditions both generate a potentially fatal reduction in LI levels for the crabs; hence, these salinities must be avoided if the crabs are to thrive during commercial crab farming. Although the crabs are tolerant to salinity levels between 15 and 35 ppt, their growth rates are impacted. The following four metabolic responses to changing environmental salinity were described by Kinne39, Type 1: Metabolic rate increasing in below-normal salinities and decreasing in above-normal salinities, Type 2: Metabolic rate increasing in below and above-normal salinities, Type 3: Metabolic rate decreasing in below and above-normal salinities and Type 4: Metabolic rate unaffected by varying salinity levels. The response exhibited by P. pelagicus can therefore be categorized as Type 3.
Salinity clearly affects fitness-determining factors such as lifespan, growth rate, time to maturity, age and size at first reproduction and offspring size and number40. Other studies have attributed decreased growth at high salinities to reduced consumption or assimilation of feed41-43. In the present study, changes in metabolic responses to environmental salinity levels revealed several intriguing patterns. The crab samples cultured at the highest and lowest salinities appeared to be less active when presented with food. The observed effects of salinity upon LF accumulation indicated an immediate and significant effect of salinity upon survival and growth, with salinities between 20 and 35 ppt being recommended for culturing P. pelagicus. The natural distribution of P. pelagicus in the Setiu Wetlands appears to reflect this salinity range. A notable difference in LI levels of fresh and frozen samples was observed, with higher LI in the frozen samples. This contrasts with the findings of Crowley44, who reported notably higher LI levels in fresh samples relative to frozen samples. Crowley's result can be attributed to the impacts of freezing upon cell structure and integrity. Maggi and Riddle45 demonstrated significant changes and damage to the lysosomal membrane during freezing. Similarly, Prento46 indicated that freezing results in rapid deterioration of cell organization and organelles, as well as increased enzyme leakage.
These results will be applicable in numerous respects, for example in the selection of farm locations and for optimising productivity by controlling salinity levels. This information provides solid baseline data that will enhance our understanding of P. pelagicus habitat preferences. The effects of salinity and freezing on LI as shown here may also served as a useful guideline for improved modeling of age estimation in crustaceans.
CONCLUSION
The optimum salinity for growth of P. pelagicus is 25 ppt. This has major implications for aquaculture of P. pelagicus, informing the selection of farm locations and the maximisation of productivity by optimization of salinity levels. Moreover, frozen samples displayed higher LI levels than fresh samples.
SIGNIFICANCE STATEMENT
This study is first to discover the effects of salinity and the preservation process upon lipofuscin levels in the eyestalks especially in P. pelagicus that can be beneficial for further population dynamic and related stock enhancement studies. This study will help the researcher to uncover the critical areas of effects of various environmental affects that many researchers were not able to explore. Thus a new theory on preservation process of lipofuscin extraction may be arrived at.
ACKNOWLEDGMENTS
The present study was supported by the Ministry of Education, Malaysia under the Niche Research Grant Scheme (Vot. No. 53131) to MI. The first author is grateful for the financial support received from this organization for the Postgraduate Scholarship Programme (MyPhD - MyBrain15). Special thanks to Dr. Mohamad N Azra for providing valuable comments and English-language revisions.
REFERENCES
- Kilada, R. and J.G. Driscoll, 2017. Age determination in crustaceans: A review. Hydrobiologia, 799: 21-36.
CrossRefDirect Link - Koeller, P.A., C. Fuentes-Yaco and T. Platt, 2007. Decreasing shrimp (Pandalus borealis) sizes off Newfoundland and Labrador-environment or fishing? Fish. Oceanogr., 16: 105-115.
CrossRefDirect Link - Ismail, N.H., A.A. Safwan, N.F. Fozi, F.H. Megat and H.M. Farouk et al., 2017. Study on carapace width growth band counts relationship of orange mud crab, Scylla olivacea (Herbst, 1796) from Terengganu Coastal waters, Malaysia. Pak. J. Biol. Sci., 20: 140-146.
CrossRefDirect Link - Ikhwanuddin, M., A. Amin-Safwan, N. Hasyima-Ismail and M.N. Azra, 2019. Dataset on body weight, carapace width increment and growth band count of mud crabs, Scylla olivacea. Data Brief, Vol. 26.
CrossRefDirect Link - Fasya, A.H., 2019. Study of patterns in the relationship of ecdysis with the age of freshwater crayfish Cherax quadricarinatus aged 76 days. IOP Conf. Series: Earth Environ. Sci., Vol. 236, No. 1.
CrossRefDirect Link - Ju, S.J., D.H. Secor and H.R. Harvey, 1999. Use of extractable lipofuscin for age determination of blue crab Callinectes sapidus. Mar. Ecol. Progr. Ser., 185: 171-179.
CrossRefDirect Link - Islam, M.S. and H. Kurokura, 2009. Accumulation of lipofuscin and preliminary estimation of age-structure in wild mud crab (Scylla paramamosain) population in tropical mangrove swamps, Thailand. J. Fish. Aquatic Sci., 4: 191-202.
CrossRefDirect Link - Wahle, R.A., O. Tully and V. O'Donovan, 1996. Lipofuscin as an indicator of age in crustaceans: Analysis of the pigment in the American lobster Homarus americanus. Mar. Ecol. Progress Ser., 138: 117-123.
CrossRefDirect Link - Belchier, M., L. Edsman, M.R.J. Sheehy and P.M. Shelton, 1998. Estimating age and growth in long-lived temperate freshwater crayfish using lipofuscin. Freshwater Biol., 39: 439-446.
CrossRefDirect Link - Sheehy, M.R.J., 1992. Lipofuscin age-pigment accumulation in the brains of ageing field-and laboratory-reared crayfish Cherax quadricarinatus (von Martens) (Decapoda: Parastacidae). J. Exp. Mar. Biol. Ecol., 161: 79-89.
CrossRefDirect Link - Nicol, S., M. Stolp and G.W. Hosie, 1991. Accumulation of fluorescent age pigments in a laboratory population of Antarctic krill Euphausia superba Dana. J. Exp. Mar. Biol. Ecol., 146: 153-161.
CrossRefDirect Link - Azra, M.N., M. Ikhwanuddin and A.B. Abol-Munafi, 2019. Behavioural data on instar crab movement at different thermal acclimation. Data Brief, 22: 998-1002.
CrossRefDirect Link - Abol-Munafi, A.B. and M.N. Azra, 2018. Climate change and the crab aquaculture industry: Problems and challenges. J. Sustainability Sci. Manage., 13: 1-3.
Direct Link - Azra, M.N., J.C. Chen, M. Ikhwanuddin and A.B. Abol-Munafi, 2018. Thermal tolerance and locomotor activity of blue swimmer crab Portunus pelagicus instar reared at different temperatures. J. Thermal Biol., 74: 234-240.
CrossRefDirect Link - Abol-Munafi, A.B., M. Mohd-Syafiq, M.A. Roswati, M.N. Azra, G. Azmie and M. Ikhwanuddin, 2016. Histological profile and fatty acid composition in hepatopancreas of blue swimming crab, Portunus pelagicus (Linnaeus, 1758) at different ovarian maturation stages. Turk. J. Fish. Aquatic Sci., 16: 251-258.
CrossRefDirect Link - Azra, M.N. and M. Ikhwanuddin, 2015. Larval culture and rearing techniques of commercially important crab, Portunus pelagicus (Linnaeus, 1758): Present status and future prospects. Songklanakarin J. Sci. Technol., 37: 135-145.
Direct Link - Ikhwanuddin, M., M.N. Azra, H. Siti-Aimuni and A.B. Abol-Munafi, 2012. Fecundity, embryonic and ovarian development of blue swimming crab, Portunus pelagicus (Linnaeus, 1758) in Coastal Water of Johor, Malaysia. Pak. J. Biol. Sci., 15: 720-728.
CrossRefDirect Link - Azra, M.N., J.C. Chen, T.H. Hsu, M. Ikhwanuddin and A.B. Abol-Munafi, 2019. Growth, molting duration and carapace hardness of blue swimming crab, Portunus pelagicus, instars at different water temperatures. Aquacult. Rep., Vol. 15.
CrossRefDirect Link - Ikhwanuddin, M., A.B. Abol-Munafi and M.N. Azra, 2019. Data on the molting duration and time of hardening of instar crab at different culture temperatures. Data Brief, Vol. 25.
CrossRefDirect Link - Taufik, M., Z. Bachok, M.N. Azra and M. Ikhwanuddin, 2016. Effects of various microalgae on fatty acid composition and survival rate of the blue swimming crab Portunus pelagicus larvae. Indian J. Geo-Mar. Sci., 45: 1512-1521.
Direct Link - Abol-Munafi, A.B., N. Pilus, R.M. Amin, M.N. Azra and M. Ikhwanuddin, 2017. Digestive enzyme profiles from foregut contents of blue swimming crab, Portunus pelagicus from Straits of Johor, Malaysia. J. Assoc. Arab Univ. Basic Applied Sci., 24: 120-125.
CrossRefDirect Link - Ikhwanuddin, M., A.N. Liyana, M.N. Azra, Z. Bachok and A.B. Abol-Munafi, 2014. Natural diet of blue swimming crab, Portunus pelagicus at strait of Tebrau, Johor, Malaysia. Sains Malaysiana, 43: 37-44.
Direct Link - Chai, C.J., Y.B. Esa, M.F.S. Ismail and M.S. Kamarudin, 2017. Population structure of the blue swimmer crab Portunus pelagicus in coastal areas of Malaysia inferred from microsatellites. Zool. Stud., Vol. 56.
CrossRefDirect Link - Romano, N., M.A. Safee, M. Ebrahimi and A. Arshad, 2016. Fatty acid compositional changes during the embryonic development of the swimming crab, Portunus pelagicus (Portunidae: Decapoda). Invertebr. Reprod. Dev., 60: 112-117.
CrossRefDirect Link - Tureli, C. and I. Yesilyurt, 2018. Reproductive biology of blue swimming crab, Portunus segnis (Forskal, 1775) in Yumurtalık Cove, Northeastern Mediterranean, Turkey. Mediterr. Mar. Sci., 18: 424-432.
CrossRefDirect Link - Ahyat, N.M., F. Mohamad, A. Ahmad and A.A. Azmi, 2017. Chitin and chitosan extraction from Portunus pelagicus. Malays. J. Anal. Sci., 21: 770-777.
CrossRefDirect Link - Amin-Safwan, A., H. Muhd-Farouk, M. Nadirah and M. Ikhwanuddin, 2016. Effect of water salinity on the external morphology of ovarian maturation stages of orange mud crab, Scylla olivacea (Herbst, 1796) in captivity. Pak. J. Biol. Sci., 19: 219-226.
CrossRefDirect Link - Ikhwanuddin, M.H.D., M.N. Azra, N.F. Noorulhudha, A. Siti-Aishah and A.B. Abol-Munafi, 2016. Embryonic development and hatching rate of blue swimming crab, Portunus pelagicus (Linnaeus, 1758) under different water salinities. Turk. J. Fish. Aquat. Sci., 16: 669-677.
CrossRefDirect Link - Shentu, J., J. Xu and Z. Ding, 2015. Effects of salinity on survival, feeding behavior and growth of the juvenile swimming crab, Portunus trituberculatus (Miers, 1876). Chin. J. Oceanol. Limnol., 33: 679-684.
CrossRefDirect Link - Puckett, B.J., D.H. Secor and S.J. Ju, 2008. Validation and application of lipofuscin-based age determination for Chesapeake Bay blue crabs Callinectes sapidus. Trans. Am. Fish. Soc., 137: 1637-1649.
CrossRefDirect Link - Ikhwanuddin, M., M.L. Shabdin and A.B. Abol-Munafi, 2009. Size at maturity of blue swimming crab (Portunus pelagicus) found in Sarawak coastal water. J. Sustainability Sci. Manage., 4: 56-65.
Direct Link - Allain, R.N., M. Moriyasu, B.D. Crawford and S.C. Courtenay, 2011. Lipofuscin quantification as a potential tool for age estimation in snow crabs, Chionoecetes opilio (O. Fabricius, 1788) (Decapoda, Oregoniidae). Crustaceana, 84: 1441-1463.
CrossRefDirect Link - Romano, N. and C. Zeng, 2006. The effects of salinity on the survival, growth and haemolymph osmolality of early juvenile blue swimmer crabs, Portunus pelagicus. Aquaculture, 260: 151-162.
CrossRefDirect Link - Falconer, T.R.L., I.D. Marsden, J.V. Hill and C.N. Glover, 2019. Does physiological tolerance to acute hypoxia and salinity change explain ecological niche in two intertidal crab species? Conserv. Physiol., Vol. 7, No. 1.
CrossRefDirect Link - Anand, T. and P. Soundarapandian, 2011. Sea ranching of commercially important blue swimming crab Portunus pelagicus (Linnaeus, 1758) in Parangipettai coast. Int. J. Sci. Nat., 2: 215-219.
Direct Link - Suratman, S., A.N.A.R. Hussein, M.T. Latif and K. Weston, 2014. Reassessment of physico-chemical water quality in Setiu Wetland, Malaysia. Sains Malays., 43: 1127-1131.
Direct Link - Suratman, S., A.N.A.R. Hussein, N.M. Tahir, M.T. Latif, R. Mostapa and K. Weston, 2016. Seasonal and spatial variability of selected surface water quality parameters in Setiu wetland, Terengganu, Malaysia. Sains Malays., 45: 551-558.
Direct Link - Kinne, O., 1964. The effects of temperature and salinity on marine and brackish water animals: 2. Salinity and temperature-salinity combinations. Oceanogr. Mar. Biol.: Annu. Rev., 2: 281-339.
Direct Link - Grzesiuk, M. and A. Mikulski, 2006. The effect of salinity on freshwater crustaceans. Pol. J. Ecol., 54: 669-674.
Direct Link - Kumulu, M. and D.A. Jones, 1995. Salinity tolerance of hatchery-reared postlarvae of Penaeus indicus H. Milne Edwards originating from India. Aquaculture, 130: 287-296.
CrossRefDirect Link - Lemos, D., V.N. Phan and G. Alvarez, 2001. Growth, oxygen consumption, ammonia-N excretion, biochemical composition and energy content of Farfantepenaeus paulensis Perez-Farfante (Crustacea, Decapoda, Penaeidae) early postlarvae in different salinities. J. Exp. Mar. Biol. Ecol., 261: 55-74.
CrossRefDirect Link - Sang, H.M. and R. Fotedar, 2004. Growth, survival, haemolymph osmolality and organosomatic indices of the western king prawn (Penaeus latisulcatus Kishinouye, 1896) reared at different salinities. Aquaculture, 234: 601-614.
CrossRefDirect Link - Maggi, V. and P.N. Riddle, 1965. Histochemisty of tissue culture cells: A study of the effects of freezing and of some fixatives. J. Histochem. Cytochem., 13: 310-317.
Direct Link - Prento, P., 1997. The effects of freezing, storage, and thawing on cell compartment integrity and ultrastructure. Histochem. Cell. Biol., 108: 543-547.
CrossRefDirect Link