
O.S. Folorunso
Department of Microbial, Biochemical and Food Biotechnology, Faculty of Natural and Agricultural Sciences, University of the Free State, 9301 Bloemfontein, South Africa
LiveDNA: 27.30539
S. Adeola
Department of Biochemistry, Faculty of Science, Lagos State University, P.O. Box 0001, Lasu Post Office, Ojo, Lagos, Nigeria
A.O. Giwa Ajeniya
Department of Chemistry, Faculty of Science, Lagos State University, P.O. Box 0001, Lasu Post Office, Ojo, Lagos, Nigeria
LiveDNA: 234.23067
Pakistan Journal of Biological Sciences
Year: 2020 | Volume: 23 | Issue: 5 | Page No.: 628-637







ABSTRACT
Background and Objective: Medicinal plants are fast becoming essential pharmaceuticals for disease and infection management. The vast antimicrobial properties of these plants reside in the inhibitory properties of their endogenous secondary metabolites. Therefore, this study aimed to assess if the volatile oil of Syzygium samarangense inhibits enteric bacteria growth and its effect against the caseinolytic activity of the extracellular protease of Salmonella typhimurium. Materials and Methods: The volatile oil was extracted by hydrodistillation, while the antimicrobial assay was assessed with the microdilution method. The extracellular protease was partially purified by salting out, followed by size-exclusion chromatography. The mode of inhibition of this enzyme was deduced from the enzyme-substrate kinetics using a line-weaver burke plot. Results: The antimicrobial properties of the oil were reported against ten enteric bacteria. Proteus vulgaris has the highest IC50 value of 0.75±0.004% v/v, while S. typhimurium, the most sensitive bacterium, showed the lowest IC50 value of 0.17±0.005% v/v. The extracellular protease of S. typhimurium was partially purified to achieve 3.73 purification fold and 314.2 μmol min–1 mg–1 protein. The optimal caseinolytic activity of this enzyme was found at pH 7.5 and 40 °C. The protease showed significantly higher activity in the presence of Zn2+ (9.3±0.33 U min–1) as compared to the control (7.0±0.58 U min–1) (p<0.05), however, K+, Ca2+, Co2+, Ba2+, Pb2+ and Hg2+ considerably reduced the enzyme activity. The activity of this enzyme was competitively inhibited by the volatile oil as an inhibitor. Conclusion: The volatile oil of S. samarangense inhibited a wide range of enteric bacteria and, therefore proposed as a potential antimicrobial agent. Inhibiting the extracellular protease of S. typhimurium may be one of its modes of action against these pathogens.
PDF Abstract XML References Citation
Copyright: © 2020. This is an open access article distributed under the terms of the creative commons attribution License, which permits unrestricted use, distribution and reproduction in any medium, provided the original author and source are credited.
How to cite this article
O.S. Folorunso, S. Adeola and A.O. Giwa Ajeniya, 2020. Syzygium samarangense Volatile Oil Inhibited Bacteria Growth and Extracellular Protease of Salmonella typhimurium. Pakistan Journal of Biological Sciences, 23: 628-637.
DOI: 10.3923/pjbs.2020.628.637
URL: https://scialert.net/abstract/?doi=pjbs.2020.628.637
DOI: 10.3923/pjbs.2020.628.637
URL: https://scialert.net/abstract/?doi=pjbs.2020.628.637
INTRODUCTION
The antimicrobial potentials of the volatile oil (VO) from medicinal plants are of great interest in ayurvedic and alternative medicines. The VO is naturally produced as secondary metabolites and has been explored for a wide range of applications in pharmaceutical, preservation and aromatherapy1,2. The choice of the VO as a complementary antimicrobial agent is a reminiscence of the significant health advantages resided in the natural medicinal plants3.
S. samarangense (Myrtaceae) has been observed as a medicinal plant of momentous value. It has edible fruits rich in phenols, flavonols and antioxidants suitable to cure hypertension and several inflammatory conditions when consumed4-6. The ethnopharmaceutical advantages of the secondary metabolites extracted from different species of Syzygium have been described and recently reviewed by Chua et al.7. The VO of the Syzygium fruits has been analyzed to contain hexanal, (Z)-hex-3-enal, hexan-1-ol and trans-caryophyllene8 while the oil from the leaves contained γ-terpinene, α-pinene and p-cymene as significant components9,10. Though a lot had been reported about the physiological evaluation and the ethnobotanical importance of this plant, however, no efforts have been directed towards the antimicrobial effect of its volatile oil (VO)8,11.
S. typhimurium (Enterobacteriaceae) is a causative agent of salmonellosis and typhoid infections with a current mortality rate of 144, 500 in 15.5 million cases12-14. This Gram-negative peritrichous flagellate bacillus contaminates foods and water and exclusively causes systemic infection in human15,16. Together with other enteric pathogens, S. typhimurium possesses a type III secretion effector proteins, which allow for the release of self-defence proteases17-19. The strategic release of extracellular peptidases and proteases to digest the host extracellular matrix proteins facilitates the pathogenicity of these bacteria20,21. Therefore, targeting the activities of these proteases will be of great benefit in antimicrobial chemotherapy, especially against the emerging MDR pathogens22,23.
In this study, the potential of S. samarangense VO to inhibit microbial growth from a range of pathogenic enteric and nosocomial bacteria was evaluated. Furthermore, enzyme-substrate kinetics of partially purified extracellular protease of S. typhimurium was monitored in the presence of the VO as a potential inhibitor. The model adopted in this study enables a prediction of the mode of inhibition the VO imposed on the extracellular protease activity and may give insight into the general antimicrobial effect of this oil against the pathogens.
MATERIALS AND METHODS
This study spanned through a period of nine months starting from January, 2019 and the analysis was jointly carried out at the Lagos State University, Ojo, Lagos, Nigeria and the University of the Free State, Bloemfontein, South Africa.
Collection of S. samarangense plant: The S. samarangense foliage was located and collected at Ojo Local Government Area of Lagos State (Coordinates N 6° 28' 6" and E 3° 10' 59"). The sample was authenticated at the departmental herbarium, Department of Botany, Lagos State University Ojo Lagos (Voucher No: LASU_BOT/2019/02/13/01_010). The leaves were air-dried for seven days, chopped and weighed.
Collection and culturing of bacteria: Ten different enteric and nosocomial pathogenic Gram-negative bacteria were obtained from the Department of Microbiology of the Lagos State University Teaching Hospital (LASUTH) and the Nigerian Institute of Medical Research (NIMR) Lagos Nigeria. Each of the bacterial isolates was revived on Nutrient agar (Merck) but used on Mueller-Hinton Agar (MHA) (Merck) for optimum growth of the bacteria during susceptibility tests.
Extraction of the VO of S. samarangense by hydrodistillation: The VO was extracted under the optimum operating conditions24. Briefly, 500 g of dried leaves was packed into the 34/35 Quick-Fit 5 L flask containing 2.0 L of distilled water with fitted Clevenger condenser. The VO was collected over 2 mL n-hexane (Merck) for 3 h at 80°C, dried over anhydrous copper II tetraoxosulphate VI (Merck) and stored at 4°C until it was used.
Microbial growth inhibition by the VO of S. samarangense: The minimum inhibitory concentration (MIC) and minimum bactericidal concentration (MBC) of the VO were analysed using the micro-broth dilution method with little modification25. Briefly, Mueller-Hinton broth (MHB) was prepared with 0.5% v/v Tween-80 (Sigma) as a dissolving medium for the VO. Each of the microwell plates received 200 μL broth. Also, 200 μL of the VO was added to the first microwell in each row of the microplate (21.5×17 cm) to prepare two-fold serial dilution of the oil in each row. At 4°C, each of the microwells was quickly inoculated with 5.0×105 CFU mL–1 bacterial suspension26.
The initial OD620nm was taken with a microplate reader (EZ Read 800, Biochrom) as A0. The plate was covered and incubated at 37°C for 18 h in a shaker at 350 rpm and another OD620nm was read as A1. About 3 μL of each of the cultures in the microwell was streaked on the freshly prepared MHA and incubated for 24 h at 37°C. The growth pattern was observed in the microplate and the agar plate accordingly. Each experiment was repeated twice. The MIC was considered as the VO dilution factor that prevents visible growth of the bacteria on the agar plate, while the MBC was regarded as the VO dilution factor that ultimately kills the bacteria and prevents any observable growth. The percentage growth inhibition of the bacteria was estimated as:
This fraction represents the efficiency of growth inhibition by the VO27.
Preparation of the crude extracellular protease from S. typhimurium: A-25.0 mL MHB culture of S. typhimurium incubated for 24 h on a shaker at 37°C was centrifuged (Heal Force® Neofuge 13) at 13, 000×g for 10 min at 4°C. The bacterial-free clarified supernatant was decanted, stored at 4°C and used as crude enzyme extract (CEE).
Determination of the total protein and caseinolytic activity of the enzyme: The total protein was quantified using 2.0 mg mL–1 of Bovine Serum Albumin (BSA) (Sigma) as a standard purified protein. Furthermore, 100 μL of CEE was assayed with casein (Sigma) (2.0-10.0 mg mL–1) in 0.05 mM Tris-HCl buffer pH 7.2 using 0.2 mg mL–1 of L. tyrosine (Sigma) as the standard for enzyme activity. Both were measured at 750 nm (EZ Read 800, Biochrom) using Lowry protein assay containing Folin-Ciocalteu’s phenolic reagent28,29. The caseinolytic activity was defined as the amount of enzyme required to liberate 1.0 μmol of L. tyrosine in 60 sec at 37°C, while the specific activity was expressed in a unit of enzyme activity/mg protein. The 1.0 mL enzyme reaction containing substrate and CEE in ratio 5:1 by volume was monitored for 5 min at 37°C and measured spectrophotometrically at 750 nm.
Purification by dialysis of the CEE: The CEE was dialysed (Sigma Cat# D9402, >12 kDa) in 0.05mM Tris-HCl buffer (pH 7.2) for 48 h with a 55% saturated solution of ammonium tetraoxosulphate (VI) (Sigma). The dialysed sample was centrifuged at 5 000 x g for 5 min at 4°C. The sediment was reconstituted in 1.0 mL of 0.05 mM Tris-HCl pH 8.0 buffer. The total protein was estimated from the reconstituted sample, while the caseinolytic assay was carried out with a 0.6% w/v casein solution prepared with 0.05 mM Tris-HCl buffer, pH 7.2.
Purification by gel filtration of the dialysed sample: A chromatographic column (28 cm×1.5 cm) containing 72 h pre-soaked Sephadex G-100 (Sigma) at 30 mg mL–1 was set up with a flow-rate of 100 μL min–1. After the equilibration of the column with 0.05 mM Tris-HCl buffer pH 7.2, 1.0 mL of dialysed reconstituted sample was loaded on the column and 36 eluents were collected. In each eluent, total protein and caseinolytic assay were carried out.
Determination of the optimal pH and temperature for the enzyme activity: The partially purified extracellular protease of S. typhimurium was assayed with a casein solution prepared with 0.05 mM Tris-HCl, pH 6.0-8.5, to determine the optimum pH at 37°C. Similarly, the assay was repeated with the observed optimum pH at varying temperatures (25-60°C).
Caseinolytic activity of the enzyme in the presence of 1 mM of chloride solutions: This assay was conducted to determine the metallic chlorides that may probably increase the caseinolytic activity. The assay was carried out in the presence of 1 mM chloride solution of the following metallic salts Ba2+, Ca2+, Cu2+, Co2+, Fe3+, Hg2+, K+, Mn2+, Pb2+ and Zn2+.
Enzyme inhibition by the VO of S. samarangense: With 3.5% v/v of the VO as a potential inhibitor, the partially purified extracellular protease was assayed with a casein solution at the optimal pH and temperature. The VO was prepared with 0.5% v/v Tween-80. The assay contained an equal volume of the substrate and the inhibitor.
Statistical analysis: Data obtained in this study were analysed with GraphPad Prism version 8.0.2.263 for Windows, GraphPad Software, San Diego, California USA (www.graphpad.com) and MS Window Excel Spreadsheet 2019. The percentage growth inhibition by the VO against each of the susceptible pathogenic bacteria was analysed using the sigmoidal nonlinear regression curve. The IC50 and inhibition gradients were estimated from each of the curves. The magnitude of the hillslope/variable slope was directly related to the efficiency of growth inhibition by the VO and this was used to estimate the rate of inhibition per h of incubation. The data means were compared using the nonparametric unpaired t-test for the rate of inhibition and IC50. Dunnett multiple comparisons of the mean activity of the enzyme, when inhibited by 1 mM chloride salts, was done with one-way ANOVA analysis. All statistical data are presented as either Mean or Mean±SEM. The test of significant difference was at p<0.05.
RESULTS
Extraction of the VO by hydrodistillation: As shown in Table 1, the VO was extracted at a constant temperature of 80°C for 3 h with 0.80 v/w yield/dried weight sample. The hydrophobic VO extracted from S. samarangense appeared as a colourless volatile liquid at room temperature with a strong fragrance odour when perceived.
Antimicrobial activities of the VO against ten selected pathogenic bacteria: Each of the nonlinear curves shown in Fig. 1 represented the % inhibition by the VO against the bacteria growth. The IC50 of the VO was estimated to determine the inhibitory concentration that could kill 50% of the bacterial population (Fig. 2). Furthermore, the rate of growth inhibition per h was estimated from the Hillslope of each of the curves as shown in Fig. 2.
Each of the bacteria responded differently to the inhibitory effect of the VO. P. vulgaris has the highest IC50 value of 0.75±0.004% v/v, while S. typhimurium showed the lowest IC50 value of 0.17±0.005% v/v (Fig. 2). From Fig. 1, P. vulgaris has the steepest sigmoidal curve-a demonstration of the relative resistance to the VO as compared to other bacteria. E. coli, K. oxytoca, K. pneumoniae, P. aeruginosa, S. enteritidis and S. paratyphi, showed IC50 within 0.2-0.4% v/v while S. dysentariae and S. sonnei showed IC50> 0.4% v/v but <0.6% v/v (Fig. 2). The growth inhibition rate for K. oxytoca, K. pneumoniae, P. vulgaris, P. aeruginosa, S. paratyphi, S. typhimurium and S. dysentariae are within 0.1-0.2 h–1 while E. coli and S. enteritidis showed inhibition rate within 0.2-0.3 h–1. From this result, P. vulgaris showed a relatively high resistance in terms of IC50, while the most susceptible bacterium is the S. typhimurium. Likewise, S. sonnei showed the highest inhibition rate as more bacteria colonies were killed per h than any other bacterium tested, however, S. paratyphi depicted the lowest inhibition rate as fewer bacteria colonies were killed per h as compared to other bacteria tested.
Regarding the diarrhoea-causing bacteria, the average IC50 and growth inhibition rate of the VO against the Salmonella species vs. Shigella species are 0.26±0.06% v/v vs. 0.53±0.04% v/v and 0.18±0.03 vs. 0.23±0.08 h–1,` respectively. It could be stated that the VO was significantly more effective against the Salmonella species than the Shigella species (p<0.05) because a two-fold increase in the VO concentration will be needed to exert the same inhibitory effect on the latter as compared to the former.
Partial purification and characterisation of the extracellular protease of S. typhimurium: The extracellular protease of S. typhimurium was partially purified to achieve 45.6% yield, 3.73 purification fold and 5, 341.4 μmol min–1 total activity as compared to the CEE (Table 2). The highest specific caseinolytic activity was 314.2 μmol min–1 mg–1 protein, which was more than a 3.5-fold increase as compared to the CEE (Table 2).
| Table 1: | Extraction of S. samarangense VO by hydrodistillation |
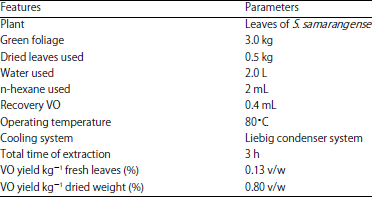 | |
| Table 2: | Partial purification profile of the extracellular protease of S. typhimurium |
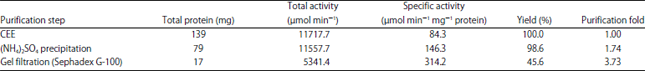 | |
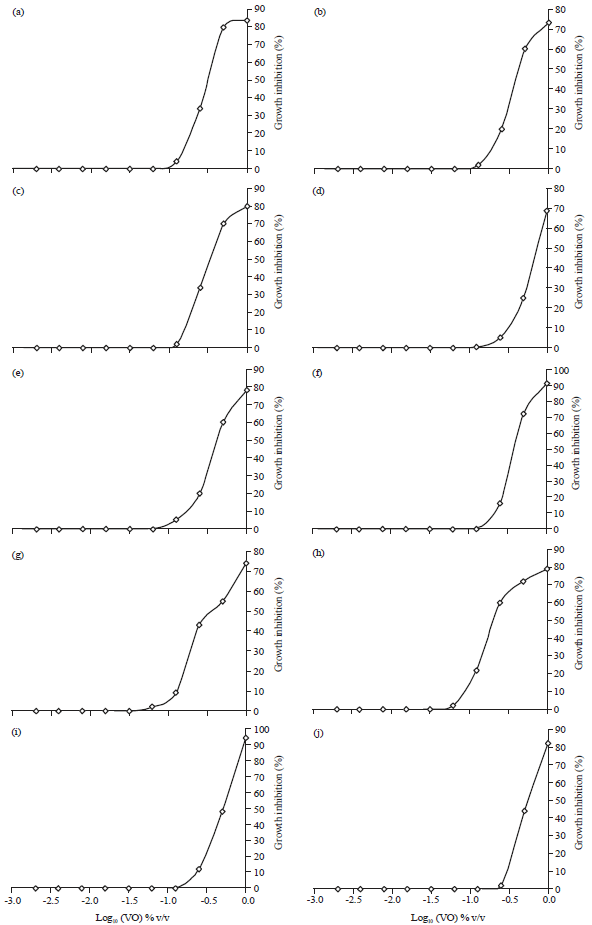 | |
| Fig. 1(a-j): | Growth inhibition curve for each bacterium susceptible to the VO of S. samarangense, (a) Escherichia coli, (b) Klebsiella oxytoca, (c) Klebsiella pneumoniae, (d) Proteus vulgaris, (e) Pseudomonas aeruginosa, (f) Salmonella enteritidis, (g) Salmonella paratyphi, (h) Salmonella typhimurium, (i) Shigella dysenteriae and (j) Shigella sonnei |
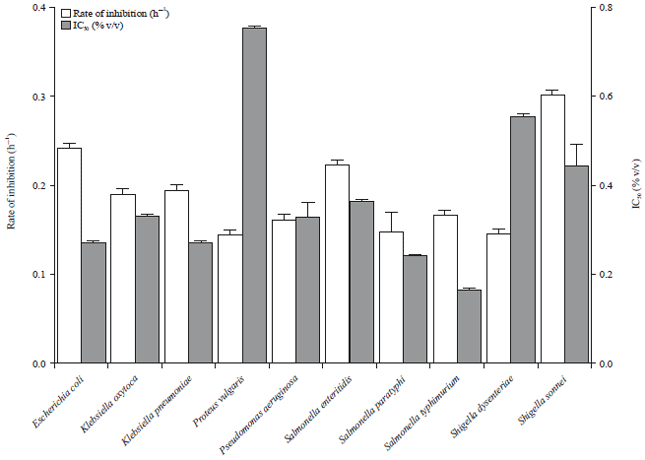 | |
| Fig. 2: | Growth inhibition and IC50 of the VO against the pathogenic bacteria (n = 2) |
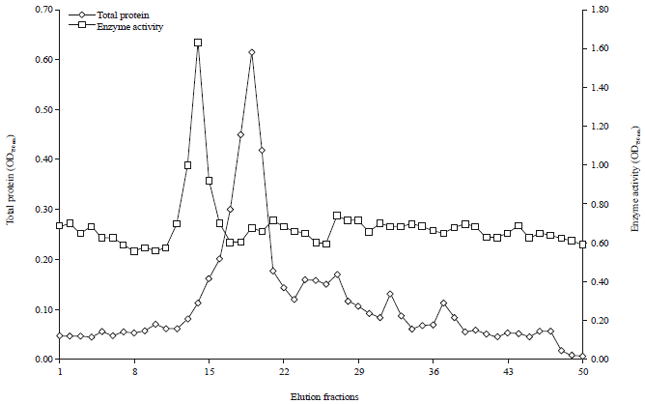 | |
| Fig. 3: | Elution profile for the partial purification of the extracellular protease of S. typhimurium |
Fractions 12-16 and 15-21 are the most probable eluents with high enzyme activity and protein content, respectively | |
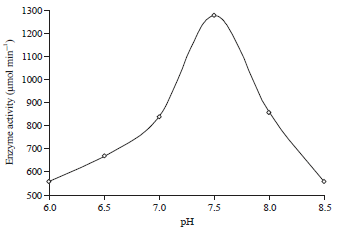 | |
| Fig. 4: | Different pH affected the activity of the partially purified extracellular protease of S. typhimurium |
| Optimum enzyme activity was found at pH 7.5, a slight alkali medium | |
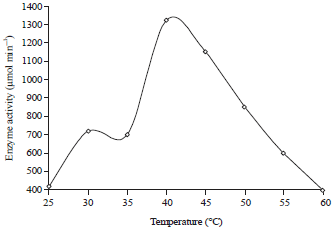 | |
| Fig. 5: | Different temperatures affected the activity of the partially purified extracellular protease of S. typhimurium |
| Optimum enzyme activity was found at 40°C | |
The elution fractions 12-16 (Fig. 3) were pooled to characterise the properties of the enzyme. The optimum caseinolytic activity was found at pH 7.5 (a slight alkali medium) (Fig. 4). The enzyme activity was monitored over a range of temperatures (25-60°C) at pH 7.5 and found to show optimum activity at 40°C after when the enzyme activity decreased steadily (Fig. 5).
Inhibition kinetics of the partially purified extracellular protease of S. typhimurium: Competitive inhibition of the enzyme was observed with casein as a substrate with the VO of S. samarangense as a co-substrate (Fig. 6). About a two-fold increase in the substrate affinity was estimated to normalise the enzyme catalysis in the presence of the inhibiting VO. Lastly, the caseinolytic assay was carried out in the presence of 1.0 mM of different metallic chlorides (Fig. 7).
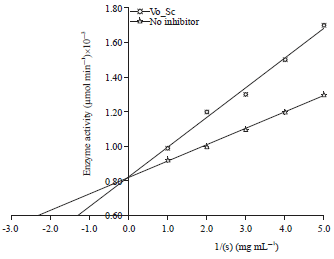 | |
| Fig. 6: | Line-weaver burke plot showing inhibition of the partially purified extracellular protease of S. typhimurium |
Line with the triangle is the enzyme activity without inhibitor, while a line with the asterisk is the enzyme activity in the presence of the VO of S. samarangense (VO_Sc). The Vmax = 1.2×103 μmol min–1, the substrate-affinity constant (Km) in the absence of inhibitor = 0.45 mg mL–1 and in the presence of inhibitor, K’m = 0.81 mg mL–1 | |
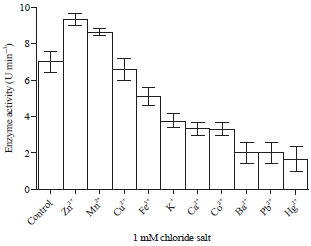 | |
| Fig. 7: | 1 mM metallic chlorides affected the activity of the partially purified extracellular protease of S. typhimurium (n = 3) |
There was a general stepwise reduction in the activity of the enzyme from transitional metallic chloride salts to heavy metal salts | |
There was no significant difference in the activities of the enzyme when the control, without any chloride salt (7.0±0.58 U min–1) was compared with the chlorides of Mn2+ (8.6±0.19 U min–1), Cu2+ (6.6±0.58 U min–1) and Fe3+ (5.1±0.51 U min–1). However, the activity of the enzyme was statistically higher in the Zn2+ (9.3±0.33 U min–1) as compared to the control (p<0.05). Chloride salts of Ba2+, Pb2+ and Hg2+drastically reduced the enzyme activity (Fig. 7).
DISCUSSION
This study demonstrated a wide range of microbial growth inhibition by the VO of S. samarangense. More importantly, there was a significant growth inhibition in all the diarrhoea-associated bacteria, which confirmed the use of this plant against diarrhoea-related infections. Competitive mode of inhibition as revealed in the enzyme-substrate-inhibitor kinetics (Fig. 6), maybe one of the modes by which the VO inhibited the growth of these bacteria.
Few studies have shown the antimicrobial activities of the oils and organic solvent extracts from different species of Syzygium plants30-33, however, there was speculation of more active antimicrobial effects in the Gram-negative than the Gram-positive bacteria perhaps as a result of a more intricate mesh of peptidoglycan layers in the latter32. In this study, all the Gram-negative bacteria tested were considered susceptible to the VO of S. samarangense and this finding supported the outcome of Raj et al.30 and De Oliveira et al.33 who worked respectively on the VO of S. gardneri and the organic extract of S. cumini and observed almost equal antimicrobial effects of the two plants against the Gram-negative and Gram-positive bacteria. Terpenes, chalcones, quercetins, flavonoids, tannins and their derivatives are the main secondary phytoconstituents of Syzygium plants30,31,34-36. Djoukeng et al.31 identified 10 different triterpenes from the leaf extract of S. guineense and discovered the most significant antibacterial activity against S. sonnei but in this study, though there was no attempt to identify the active components of the VO used, however, it is very significant that S. sonnei has the highest growth inhibition per h as compared to other bacteria (Fig. 2). This study confirmed a significant microbial growth inhibition against the diarrhoea-causing Salmonella and Shigella species. This result complements the use of this plant in diarrhoea and this therapeutic effect had been traced to the presence of calcium channel blocking flavonoid in this plant37.
This study also confirmed casein as one of the preferred substrates for the S. typhimurium extracellular protease because more than a 3.5-fold increase in specific enzyme activity was obtained during the partial purification as compared to the CEE (Table 2). The optimal conditions of enzyme activity obtained in this study further re-iterate the survival, colon colonisation, extracellular matrix protein degradation, invasion and pathogenesis of this bacterium in their hosts15,38-40.
Competitive inhibition of the extracellular protease of S. typhimurium for casein was observed with the VO of S. samarangense, which indicates a possible potential drug target. About a 2-fold increase in the substrate concentration will be needed to normalise the caseinolytic activity of the enzyme, which suggests a stronger affinity binding of such component of the VO (Fig. 6). From the extensive work of Amor et al.41, S. samarangense possessed intractable secondary metabolites that selectively inhibited serine protease. Likewise, Tezuka et al.42 screened several medicinal plant extracts and identified catechol and quinone as potential inhibitors of prolyl endopeptidase from Flavobacterium. By considering the phytoconstituents enrichment of S. samarangense VO, it is, therefore, evident that this plant contains microbial protease inhibitor(s). Regarding this study, identification and characterisation of these specific extracellular proteases will enhance further investigation into the type of protease and the exact mechanism of inhibition43-49.
The present result also indicates a metallic requirement for the activity of extracellular protease released by S. typhimurium. Chlorides of Zn2+, Mn2+, Cu2+ and to some extent, Fe3+ may be needed for the elevated caseinolytic activity of this protease. However, Ba2+, Pb2+ and Hg2+ are inhibitors because they are toxic heavy metals that can quickly alter the biological function of proteins50,51.
In this study, the specific extracellular protease of S. typhimurium was not identified and this calls for further studies to identify and classify this specific extracellular protease and their preference for casein. Furthermore, there is a need to identify the specific active component of the oil, which exhibited this antimicrobial function and this could be done by wide-range isolation and screening of these phytoconstituents.
CONCLUSION
The reports from this study show the promising benefits of the VO extracted from the dried leaves of S. samarangense as an antimicrobial agent against a wide range of enteric and nosocomial pathogenic bacteria. This VO inhibited the growth of susceptible bacteria and competitively inhibited the extracellular protease of S. typhimurium. Further identification and isolation of these bactericidal components of this VO will be of immense benefit in antimicrobial chemotherapy.
SIGNIFICANCE STATEMENT
This study discovered that the VO extracted from the leaves of S. samarangense possessed antimicrobial effect by preventing the growth of a wide-range enteric and nosocomial Gram-negative bacteria and competitively hindered the caseinolytic activity of the extracellular protease of S. typhimurium. This can be of benefit for antimicrobial development in alternative and Ayurvedic medicine. This study will help the researcher to uncover the critical areas of enzyme-substrate-inhibitor kinetics that many researchers were not able to explore. Thus, a new theory on the mode and mechanisms of drug interaction with enzyme catalysis may be arrived at.
REFERENCES
- Compean, K.L. and R.A. Ynalvez, 2014. Antimicrobial activity of plant secondary metabolites: A review. Res. J. Med. Plant, 8: 204-213.
CrossRefDirect Link - Ali, B., N.A. Al-Wabel, S. Shams, A. Ahamad, S.A. Khan and F. Anwar, 2015. Essential oils used in aromatherapy: A systemic review. Asian Pac. J. Trop. Biomed., 5: 601-611.
CrossRefDirect Link - Akhtar, M.S., B. Degaga and T. Azam, 2014. Antimicrobial activity of essential oils extracted from medicinal plants against the pathogenic microorganisms: A review. Issues Biol. Sci. Pharm. Res., 2: 1-7.
Direct Link - Izzah, K.N., Y. Awang, P. Ding, Y. Hafiza and M.G.M. Satar, 2015. Antioxidant, polygalacturonase, pectin methylesterase and polyphenol oxidase activities of fresh-cut wax apple (Syzygium samarangense) treated with organic acids. Asian J. Plant Sci., 14: 72-77.
CrossRefDirect Link - Venkatachalam, K., C. Techakanon and S. Thitithanakul, 2018. Impact of the ripening stage of wax apples on chemical profiles of juice and cider. ACS Omega, 3: 6710-6718.
CrossRefDirect Link - Chua, L.K., C.L. Lim, A.P.K. Ling, S.M. Chye and R.Y. Koh, 2019. Anticancer potential of Syzygium species: A review. Plant Foods Hum. Nutr., 74: 18-27.
CrossRefDirect Link - Sobeh, M., M.S. Braun, S. Krstin, F.S. Youssef, M.L. Ashour and M. Wink, 2016. Chemical profiling of the essential oils of Syzygium aqueum, Syzygium samarangense and Eugenia uniflora and their discrimination using chemometric analysis. Chem. Biodivers., 13: 1537-1550.
CrossRefDirect Link - Rastogi, R.P. and M.N. Mehrotra, 1995. Compendium of Indian Medicinal Plants. 1st Ed. CSIR, New Delhi, pp: 1985-1989.
Direct Link - Khandaker, M.M., A.N. Boyce, N. Osman and A.S. Hossain, 2012. Physiochemical and phytochemical properties of wax apple (Syzygium samarangense [Blume] Merrill & L. M. Perry var. Jambu Madu) as affected by growth regulator application. Sci. World J.
CrossRefDirect Link - Vos, T., A.A. Abajobir, K.H. Abate, C. Abbafati and K.M. Abbas et al., 2017. Global, regional and national age-sex specific mortality for 264 causes of death, 1980-2016: A systematic analysis for the Global Burden of disease study 2016. Lancet, 390: 1211-1259.
CrossRefPubMedDirect Link - Naghavi, M., A.A. Abajobir, C. Abbafati, K.M. Abbas and F. Abd-Allah et al., 2017. Global, regional and national age-sex specific mortality for 264 causes of death, 1980-2016: A systematic analysis for the global burden of disease study 2016. Lancet, 390: 1151-1210.
CrossRefDirect Link - Wain, J., R.S. Hendriksen, M.L. Mikoleit, K.H. Keddy and R.L. Ochiai, 2015. Typhoid fever. Lancet, 385: 1136-1145.
CrossRefDirect Link - Spano, S., X. Gao, S. Hannemann, M. Lara-Tejero and J.E. Galán, 2016. A bacterial pathogen targets a host rab-family GTPase defense pathway with a GAP. Cell Host Microbe, 19: 216-226.
CrossRefDirect Link - Spano, S. and J.E. Galan, 2012. A Rab32-dependent pathway contributes to Salmonella typhi host restriction. Science, 338: 960-963.
CrossRefDirect Link - Baldassarre, M., V. Solano-Collado, A. Balci, H.M. Wilson, S. Mukhopadhyay, G. Dougan and S. Spanò, 2019. Salmonella typhi survives in human macrophages by neutralizing the RAB32/BLOC-3 host-defence pathway. bioRxiv., 2019.
CrossRefDirect Link - Wilson, J.W., M.J. Schurr, C.L. LeBlanc, R. Ramamurthy, K.L. Buchanan and C.A. Nickerson, 2002. Mechanisms of bacterial pathogenicity. Postgraduate Med. J., 78: 216-224.
CrossRefDirect Link - Ingmer, H. and L. Brøndsted, 2009. Proteases in bacterial pathogenesis. Res. Microbiol., 160: 704-710.
CrossRefDirect Link - Davin-Regli, A., J.P. Lavigne and J.M. Pagès, 2019. Enterobacter spp.: Update on taxonomy, clinical aspects and emerging antimicrobial resistance. Clin. Microbiol. Rev., Vol. 32, No. 4.
CrossRefDirect Link - Ghosh, C., P. Sarkar, R. Issa and J. Haldar, 2019. Alternatives to conventional antibiotics in the era of antimicrobial resistance. Trends Microbiol., 27: 323-338.
CrossRefDirect Link - Elyemni, M., B. Louaste, I. Nechad, T. Elkamli and A. Bouia et al., 2019. Extraction of essential oils of Rosmarinus officinalis L. by two different methods: Hydrodistillation and microwave assisted hydrodistillation. Scient. World J., Vol. 2019.
CrossRefDirect Link - Janssen, A.M., J.J.C. Scheffer, L. Ntezurubanza and A.B. Svendsen, 1989. Antimicrobial activities of some Ocimum species grown in rwanda. J. Ethnopharmacol., 26: 57-63.
CrossRefDirect Link - Folorunso, O.S., K.O. Amisu and B.F. Ogungbe, 2015. EDTA-Treated cell membrane and multiple subculturing affect the virulence of enteropathogenic bacteria. Res. J. Microbiol., 10: 158-169.
CrossRefDirect Link - Everette, J.D., Q.M. Bryant, A.M. Green, Y.A. Abbey, G.W. Wangila and R.B. Walker, 2010. Thorough study of reactivity of various compound classes toward the Folin−Ciocalteu reagent. J. Agric. Food Chem., 58: 8139-8144.
CrossRefDirect Link - Lowry, O.H., N.J. Rosebrough, A.L. Farr and R.J. Randall, 1951. Protein measurement with the folin phenol reagent. J. Biol. Chem., 193: 265-275.
CrossRefPubMedDirect Link - Raj, G., V. George, N.S. Pradeep and M.G. Sethuraman, 2008. Chemical composition and antimicrobial activity of the leaf oil from Syzygium gardneri Thw. J. Essent. Oil Res., 20: 72-74.
CrossRefDirect Link - Djoukeng, J.D., E. Abou-Mansour, R. Tabacchi, A.L. Tapondjou, H. Bouda and D. Lontsi, 2005. Antibacterial triterpenes from Syzygium guineense (Myrtaceae). J. Ethnopharmacol., 101: 283-286.
CrossRefDirect Link - Ratnam, K.V. and R.R.V. Raju, 2008. In vitro antimicrobial screening of the fruit extracts of two Syzygium species (Myrtaceae). Adv. Biol. Res., 2: 17-20.
Direct Link - De Oliveira, G.F., N.A.J.C. Furtado, A.D. da Silva Filho, C.H.G. Martins, J.K. Bastos, W.R. Cunha and M.L. de Andrade e Silva, 2007. Antimicrobial activity of Syzygium cumini (Myrtaceae) leaves extract. Braz. J. Microbiol., 38: 381-384.
CrossRefDirect Link - Nair, A.G.R., S. Krishnan, C. Ravikrishna and K.P. Madhusudanan, 1999. New and rare flavonol glycosides from leaves of Syzygium samarangense. Fitoterapia, 70: 148-151.
CrossRefDirect Link - Srivastava, R., A.K. Shaw and D.K. Kulshreshtha, 1995. Triterpenoids and chalcone from Syzygium samarangense. Phytochemistry, 38: 687-689.
CrossRefDirect Link - Nonaka, G.I., Y. AiKo, K. Aritake and I. Nishioka, 1992. Tannins and related compounds. CXIX. Samarangenins a and b, novel proanthocyanidins with doubly bonded structures, from Syzygium samarangens and S. aqueum. Chem. Pharm. Bull., 40: 2671-2673.
CrossRefDirect Link - Ghayur, M.N., A.H. Gilani, A. Khan, E.C. Amor, I.M. Villasenor and M.I. Choudhary, 2006. Presence of calcium antagonist activity explains the use of Syzygium samarangense in diarrhoea. Phytother. Res., 20: 49-52.
CrossRefPubMedDirect Link - Chaudhuri, D., A.R. Chowdhury, B. Biswas and D. Chakravortty, 2018. Salmonella typhimurium infection leads to colonization of the mouse brain and is not completely cured with antibiotics. Front. Microbiol., Vol. 9.
CrossRefDirect Link - Drevets, D.A., P.J.M. Leenen and R.A. Greenfield, 2004. Invasion of the central nervous system by intracellular bacteria. Clin. Microbiol. Rev., 17: 323-347.
CrossRefPubMedDirect Link - Graziani, C., C. Losasso, I. Luzzi, A. Ricci, G. Scavia and P. Pasquali, 2017. Salmonella. In: Foodborne Diseases: Third Edition, Dodd, C.E.R., T. Aldsworth, R.A. Stein D.O. Cliver and H.P. Riemann (Eds.)., Academic Press, New York, pp: 133-169.
Direct Link - Amor, E.C., I.M. Villaseñor, A. Yasin and M.I. Choudhary, 2004. Prolyl endopeptidase inhibitors from Syzygium samarangense (Blume) Merr. & LM Perry. Zeitschrift Naturforschung C, 59: 86-92.
CrossRefDirect Link - Tezuka, Y., W. Fan, R. Kasimu and S. Kadota, 1999. Screening of crude drug extracts for prolyl endopeptidase inhibitory activity. Phytomedicine, 6: 197-203.
CrossRefDirect Link - Ramu, P., L.A. Lobo, M. Kukkonen, E. Bjur and M. Suomalainen et al., 2008. Activation of pro-matrix metalloproteinase-9 and degradation of gelatin by the surface protease PgtE of Salmonella enterica serovar Typhimurium. Int. J. Med. Microbiol., 298: 263-278.
CrossRefDirect Link - Lewis, C., H. Skovierova, G. Rowley, B. Rezuchova and D.H.A. Stevenson et al., 2009. Salmonella enterica serovar Typhimurium HtrA: Regulation of expression and role of the chaperone and protease activities during infection. Microbiology, 155: 873-881.
CrossRefDirect Link - Bäumler, A.J., J.G. Kusters, I. Stojiljkovic and F. Heffron, 1994. Salmonella typhimurium loci involved in survival within macrophages. Infect. Immunity, 62: 1623-1630.
Direct Link - Riva, R., T.K. Korhonen and S. Meri, 2015. The outer membrane protease PgtE of Salmonella enterica interferes with the alternative complement pathway by cleaving factors B and H. Front. Microbiol., Vol. 6.
CrossRefDirect Link - Wachtel, R., B. Bräuning, S.L. Mader, F. Ecker, V.R. Kaila, M. Groll and A. Itzen, 2018. The protease GtgE from Salmonella exclusively targets inactive Rab GTPases. Nat. Commun., Vol. 9, No. 1.
CrossRefDirect Link - Tamás, M.J., S.K. Sharma, S. Ibstedt, T. Jacobson and P. Christen, 2014. Heavy metals and metalloids as a cause for protein misfolding and aggregation. Biomolecules, 4: 252-267.
CrossRefDirect Link - Sharma, S.K., P. Goloubinoff and P. Christen, 2008. Heavy metal ions are potent inhibitors of protein folding. Biochem. Biophys. Res. Commun., 372: 341-345.
CrossRefDirect Link