
Almoataz Bellah Ali El-Mouhamady
Division of Genetic Engineering and Biotechnology, Department of Genetics and Cytology, National Research Centre, 33 El Buhouth Street, 12622 Dokki, Giza, Egypt
LiveDNA: 20.25983
Mohamed Ahmed El-Metwally
Departmant of Mycology Research, Plant Pathology Research Institute, Agricultural Research Center, Giza, Egypt
LiveDNA: 20.359
Pakistan Journal of Biological Sciences
Year: 2020 | Volume: 23 | Issue: 4 | Page No.: 425-438







ABSTRACT
Background and Objective: Soybean crop occupies a high position among cereal crops because it is a highly nutritious plant. This investigation aimed to take advantage of the imported soybean lines in improving the local varieties. Materials and Methods: Three local varieties and 4 imported lines of soybean plant were planted during two seasons for estimating some genetic parameters including (Heritability in broad sense, PCV (%), GCV (%), DZ, expected genetic advance and genetic advance as percentage of mean) in some agro-morphological traits besides molecular characterization for the seven soybean materials using 6 SCoT primers. Results: The four imported soybean genotypes besides the local cultivar Giza 111 exhibited highly trend for all studied traits in both seasons. Heritability and genetic advance were exhibited significance and high values in most studied traits under both seasons. Conclusion: The imported soybean genotypes confirmed significant excellence in all studied traits and recorded highly genetic stability for both seasons depending on results obtained of all genetic parameters. Therefore, the decision to use it as a new and good genetic source to improve, develop and raise the productivity of Egyptian soybean varieties as well as its resistance to biotic and abiotic stresses will be right and fruitful.
PDF Abstract XML References Citation
Received: November 08, 2019;
Accepted: December 24, 2019;
Published: March 15, 2020
Copyright: © 2020. This is an open access article distributed under the terms of the creative commons attribution License, which permits unrestricted use, distribution and reproduction in any medium, provided the original author and source are credited.
How to cite this article
Almoataz Bellah Ali El-Mouhamady and Mohamed Ahmed El-Metwally, 2020. Appreciation of Genetic Parameters and Molecular Characterization in Some Promising Accessions of Soybean (Glycine max L.). Pakistan Journal of Biological Sciences, 23: 425-438.
DOI: 10.3923/pjbs.2020.425.438
URL: https://scialert.net/abstract/?doi=pjbs.2020.425.438
DOI: 10.3923/pjbs.2020.425.438
URL: https://scialert.net/abstract/?doi=pjbs.2020.425.438
INTRODUCTION
Soybean is one of the most important agricultural, food and industrial crops in the world. Its importance is summarized in the containment seeds on a protein of 40% and oil without cholesterol with 20%. Soybean farming has been successful in on a large scale in Egypt. The Egyptian Ministry of Agriculture has registered an agricultural area up to 22.2 thousand feddan with a production capacity reach to 25000 t in order to take advantage of this crop as animal feed and food important and vital to humans. As well as a large sector of important food industries such as not limited white cheese from soybean milk, yogurt and ice cream. The purpose of this study was to describe the genetic and environmental differences among promising soybeans accessions imported from different regions and planted under Egyptian conditions during two consecutive years. As well as identification of similarities and differences at the molecular genetics level using a set of primers such as RAPD or ISSR or SCoT PCR-analysis. Thus, determination the extent of benefit from these new accessions for improving Egyptian soybean cultivars to face the biotic and a biotic stresses would be very fruitful. The following is a quick review for the most important results of some papers carried out in this regard. The genetic variability on 16 traits associated with yield and its components in 25 soybean accessions was showed by Malik et al.1. It was observed that highly percentages of heritability were observed in some traits such as, 100-grain weight, days to maturity, days to flowering completion and grain yield/plant. Sundry genetic breeding methods participated to the improvement of soybean accessions with high output and acclimatized to various Brazilian edaphoclimatic conditions were revealed by Hamawaki et al.2. It could be concluded that the persistent advance of genetic improvement through traditional breeding for some soybean genotypes abut the genetic variability and implementation of genetic parameters input which assist a more effective selection operation. In the same context3 studied the genetic variability including genetic and phenotypic variation besides heritability in broad sense in some Egyptian cultivars of soybean namely, Crawford, Giza 21, Giza 22, Giza 35, Giza 82, Giza 83 and Giza 111 for some agro-morphological traits and showed that heritability in broad sense was high in plant height, number of pods/plant and grain yield/plant traits which means that the environmental variation on these characters was very little with highly rank for the values of genetic stability in these traits conversely the other characters. On the other hand, the study dealt with the molecular description between these genotypes found that the primers (OPC10, OPF-4, OPA-17 and OPG-5) generated 12 fragments where (8 of them were polymorphic with 66.67% polymorphism besides 4 bands were monomorphic). In the same regard4 calculated some genetic parameters in 41 soybean accessions in addition, 5 checks for some yield and its components traits and showed that highly results of GCV and PCV were observed in some studied traits such as, number of pods/plant (35.45, 38.62), plant height (23.49, 25.84) and yield/plant (21.35, 24.86) which confirmed appear of highly impact of genetic variation with significantly form. While, plant height, number of pods/plant, grain yield/plant and 1000-grain weight recorded highly values of heritability besides high genetic advance. The genetic divergence and inhabitance framework of 205 terrestrial Glycine max nucleus combinations in Northeast China from 9 latitude clans and 9 longitude hordes were appreciated through using SSR markers5. A total of 973 alleles were revealed by 43 SSR loci and the median number of alleles per locus was 22.628 in addition, molecular depiction using 54 SSRs primers pairs and genetic diversity were done in 8 soybean accessions6. The final results confirmed that the 8 soybean genotypes were generated 216 alleles using SSRs primers and cluster analysis consisted all soybean materials into two main cluster having 06 and 02 accessions. Li et al.7 discussed cross identification and salinity tolerance in glycine max and glycine soja through using SSR markers and detected that the collection among molecular SSR markers and simple physiological parameters was the ideal and optimum technique used for testing and screening the veritable crosses of glycine max and glycine soja for salinity tolerance besides fabricating anew soybean cultivars tolerance for this stress. The aim of this investigation is determine the limit of genetic stability for a group of new soybean imports compared to local varieties to consider the possibility of cultivating these new materials in the future under Egyptian conditions to benefit from its positive advantages in the soybean breeding program.
MATERIALS AND METHODS
Genetic materials: The present study was carried out in the farm of National Research Centre in (Nubaria, Beheira Governorate, Egypt) and Genetics and Cytology Department, Genetic Engineering and Biotechnology Division, National Research Centre, Dokki, Giza, Egypt in the period from the first of May to the first of September in seasons 2017 and 2018, respectively. This investigation aimed to know the genetic behavior of 7 soybean accessions included 4 imported lines from different regions and tolerance for abiotic stresses in addition, resistance for many diseases where the lines (PR-162-11 and Ocepara 4) were imported from USA (FB1-7636 and SR-4-1) were performed from brazil beside the three Egyptian cultivars (Giza 21, Giza 82 and Giza 111) (Table 1), respectively.
| Table 1: | Pedigree and origin of seven soybean genotypes |
 | |
| All imported varieties were planted after performing quarantine agricultural procedures | |
Methods
Studied traits: Fifty plants for each genotype for each season were using to calculate and estimate some agro-morphological characters as follows:
| • | Plant height (cm): Measured at maturity as the length in centimeter from ground level to the tip of the main stem |
| • | First pod height (cm): Measured at maturity the length in centimeter from the ground level to up to the insertion node of the first pod on the main stem |
| • | Number of pods/plant: Obtained by counting the total number of filled pods/plant at maturity |
| • | Number of nodes/plant: Obtained by counting the total number of nodes/plant at maturity |
| • | Number of grains/plant: Obtained by counting threshed, air-dried seeds/plant |
| • | 1000-grain weight (g): Recorded in g/1000 random whole seeds after air-drying to uniform moisture |
| • | Grain yield/plant (g): Measured from mass of threshed, air-dried grains/plant |
Sowing: The seven soybean genotypes were planting in the period from the 1st May-1st September in seasons 2017 and 2018 in a randomized complete block design with three replicates for each experiment in each season. Each cultivar was grown in 3 rows for each replicate where each replicate included 21 rows, length of row was 10 m, 25 cm between each 2 rows and 20 cm space between each 2 plants in row. All recommended agriculture practices were applied.
Experimental replicate area: The 52.5 m2 (Length was 10 m, 5.25 m was width of each replicate) and irrigation method was sprinkler irrigation.
Statistical analysis: Data obtained from all studied traits of the seven soybean genotypes was analysis using the SPSS ver.17 and analysis of variance was detected as recorded by Gomez and Gomez8.
Estimates of genetic parameters: Variance components, heritability in broad sense, genetic coefficient of variability (GCV (%)), phenotypic coefficient of variability (PCV (%)), Dz or the difference between the phenotypic coefficient of variation (PCV (%)) and genotypic coefficient of variation (GCV (%)), expected genetic advance in addition, genetic advance as percentage of mean were the most important measurements calculated through the two seasons in this study as follows.
The genetic coefficient of variability (GCV (%)) and phenotypic coefficient of variability (PCV (%)) were estimates according to the method suggested by Burton and Devane9 as follows:
Environmental variance (σ2e) = MSe
Genotypic variance (Gv) or (σ2g) = Msg-MSe/r
Phenotypic variance (Phv) or (σ2ph) = (σ2e)+(σ2g) or Mse+MSg
Where:
| MSe | = | Mean square of error |
| MSg | = | Mean square of genotypes |
| r | = | Number of replicates |
Genetic coefficient of variability ![]()
Phenotypic coefficient of variability ![]()
Estimation of heritability in broad sense: Broad sense heritability (H2) expressed as the percentage of the ratio of the genotypic variance (gv) to the phenotypic variance (phv) and was estimated on genotype mean basis as described by Burton and Devane9 and Johnson et al.10 as:
The Dz is the difference between the phenotypic coefficient of variation PCV (%) and genotypic coefficient of variation GCV (%) or PCV (%)-GCV (%).
Estimation of genetic advance: The expected genetic advance (GA) and percentage of the mean (GAM) assuming selection of superior 5% of the genotypes was estimated in accordance with the methods illustrated by Johnson et al.10 as:
where, K is the standardized selection differential at 5% selection intensity (K = 2.068).
The genetic advance as percentage of mean (GAM) was computed as:
GAM (%) = (GA)/×100
Estimation of simple phenotypic correlation coefficients: Simple phenotypic correlation coefficients among all studied traits in two seasons (2017 and 2018) were estimated using the formula suggested by Miller et al.11.
Molecular markers: Molecular markers technique aimed to estimate the phylogenetic tree, relationships and stand on the fragments responsible for similarities and differences among various soybean genotypes. So, this investigation studied the molecular variation generated between the seven soybean cultivars. Specifically, they have descended from various genetic origins and highly different environmental zones. Therefore, knowing the similarities and differences between them will enhance the nominal objective of this study, it is the use of imported American and Brazilian soybean varieties in the breeding programs of improvement and development Egyptian soybean crop such as for increasing yielding and quality of seeds in addition, resistance for biotic and abiotic stresses.
DNA isolation procedure: The bulked DNA extraction was performed using DNeasy plant Mini Kit (QIAGEN). Isolation protocol of DNA was as follows:
| • | Plant culture were ground using liquid nitrogen to a fine powder, then, the powder was transferred to an appropriately sized tube |
| • | Then, 400 μL of buffer AP1 and 4 μL of RNase a stock solution (100 mg mL–1) were added to a maximum of 100 mg of ground plant culture then vortexed vigorously |
| • | Mixture was incubated for 10 min at 65°C and mixed 2-3 times during incubation by inverting tube |
| • | Then, 130 μL of buffer AP2 was added to the lysate, mixed and incubated for 5 min on ice |
| • | Lysate was applied to the QIA shredder spin column sitting in a 2 mL collection tube and centrifuged for 2 min at maximum speed (10.000 rpm) |
| • | Supernatant from step 5 was transferred to a new tube without disturbing the cell-debris pellet. Typically, 450 μL of lysate was recovered |
| • | Then, 0.5 volume of buffer AP3 and 1 volume of ethanol (96-100%) were added to the cleared lysate and mixed by pipetting |
| • | Then, 650 μL of the mixture from step 7 was applied through DNeasy Mini spin column setting in a 2 mL collection tube. Then, centrifuged for 1 min at 8000 rpm and flow-through was then discarded |
| • | DNeasy column was then placed in a new 2 mL collection tube. Then, 500 μL buffer AW was added onto the DNeasy column and centrifuged for 1 min at 8000 rpm |
| • | Then, 500 μL buffer AW was added to DNeasy column and centrifuged for 2 min at maximum speed (10.000 rpm) to dry the column membrane |
| • | DNeasy column was then transferred to a 1.5 mL microfuge tube and 100 μL of preheated (65°C) buffer AE was pipetted directly onto the DNeasy column membrane. Then, incubated for 5 min at room temperature and centrifuged for 1 min at 8000 rpm to elute |
| • | Elution was repeated once as described. A new microfuge can be used for first elute. Alternatively, the microfuge tube can be reused for the second elution step to combine the elutes |
Polymerase chain reaction (PCR) condition stock solutions:
 |
Ethidium bromide:
| • | The stock solution was prepared by dissolving 1 g of ethidium bromide in 100 mL distilled water and mixed well with magnetic stirrer |
| • | Transferred to a dark bottle and stored at room temperature |
Sample loading dye (5x):
 |
The PCR was performed in 30 μL volume tubes according to Williams et al.12 that contained the following:
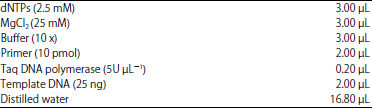 |
Polymerase chain reaction (PCR) condition for SCoT Technique: The DNA amplifications were performed in an automated thermal cycle (model Techno 512) programmed for one cycle at 94°C for 4 min followed by 45 cycles of 1 min at 94°C, 1 min at 57°C and 2 min at 72°C. the reaction was finally stored at 72°C for 10 min (Table 2).
Gel preparation procedure:
| • | Agarose (1.50 g) was mixed with (100 mL) L x TBE buffer and boiled in microwave |
| • | Ethidium bromide (5 μL) was added to the melted gel after the temperature became 55°C |
| • | The melted gel were poured in the tray of mini-gel apparatus and comb was inserted immediately, then comb was removed when the gel become hardened |
| • | The gel was covered by the electrophoretic buffer (1xTBE) |
| • | DNA amplified product (15 μL) was loaded in each well |
| • | DNA ladder (100 bp) mix was used as standard DNA with molecular weights of 3000, 1500,1000, 900, 800, 700, 600, 500, 400, 300, 200 and 100 bp. The run was performed for about 30 min at 80 V in mini submarine gel BioRad |
| Table 2: | List of the primer names and their nucleotide sequences used in the study for SCoT procedure |
 | |
Statistical analysis: The similarity matrices were done using Gel works ID advanced software UVP-England Program. The relationships among genotypes as revealed by dendrograms were done using SPSS windows (Version 10) program. Jaccard13 computer package was used to calculate the pairwise difference matrix and plot the phonogram among cultivars14.
RESULTS
Variation and interaction: Data presented in Table 3 and related with analysis of variance test revealed highly significant differences among all soybean accessions for all studied traits during the two seasons (2017 and 2018). Coefficient of variance percentage was very low and ranged from (0.48%) for 1000-grain weight trait to (16.15%) for first pod height trait in season 2018, respectively.
Mean performance: Mean values for all studied traits of the 7 soybean genotypes during the two seasons were presented in Table 4. Results obtained from the mean values of all studied traits for all soybean accessions proved that the differences between them were not significant enough to promote the existence for genetic variation between the two seasons. While, they were mostly differences due to the environmental factor. On the other hand, the soybean materials were different from each other in many traits and this point will explain in this recent context. For example, the highest mean values of plant height trait during two seasons were observed in the cultivars; (Giza 111, Giza 21 and Giza 82) and mean values were (102.25, 103.17 and 106.89 cm). While that, the highest mean values for first pod height trait in the average of two seasons were obtained in cultivars (Ocepara 4, PR-162-11 and SR-4-1) and the values were (13.29, 13.71 and 27.89 cm). In the same context, the highest average of soybean genotypes obtained in two seasons for grain yield/plant were Giza 111 (22.76 g), SR-4-1 (23.44 g), Ocepara 4 (24.81 g), PR-162-11 (25.09 g) and FB1-7636 (26.14 g), respectively and so on.
| Table 3: | ANOVA analysis of all studied traits for seven soybean genotypes in the two seasons (2017 and 2018) |
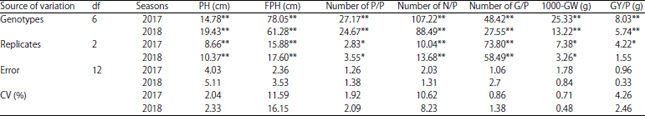 | |
| *Significant at 5% probability level, **Significant at 1% probability level, PH: Plant height, FPH: First pod height, P/P: Pods/plant, N/P: Nodes/plant, G/P: Grains/plant, GW: 1000-grain weight, GY/P: Grain yield/plant (g) | |
Genetic parameters: Data viewed in Table 5 revealed that heritability in broad sense was appeared high in all studied traits under the conditions of two seasons except plant height trait where it was recorded medium values and lowest percentages in the same time for 2 years (47.04 and 48.27%). While that, the values for the rest traits were ranged from (70.99%) in grain yield/plant to (94.52%) in number of nodes/plant for the first seasons and from (75.40%) in number of grains/plant to (95.68%) in number of nodes/plant trait for the 2nd year.
Results in Table 5 showed that the values estimated of GCV% in the 1st season (2017) were high in grain yield/plant (6.66%), 1st pod height trait (37.90%) and number of nodes/plant trait (44.15%) and so on. The same results were detected in season 2018 where grain yield/plant was recorded (5.74%), 1st pod height exhibited (37.72%) and number of nodes/plant was (38.78%), respectively. The highest values of GCV% in both seasons (2017 and 2018) were observed in number of nodes/plant trait followed by 1st pod height trait and the values were (44.15 and 38.78%) for the 1st trait and (37.90 and 37.72%) for the other one, respectively. The PCV (%) assessment was highest in the traits, grain yield/plant (7.91%), first pod height (39.64%) and number of nodes/plant (45.41%) in season 2017. While that, results of PCV% in season 2018 were shown high in grain yield/plant (6.25%), number of nodes/plant trait (39.64%) and first pod height traits (41.03%), respectively. The lowest percentages for PCV (%) in 2 years were observed in 1000-grain weight trait (1.65 and 1.19%). While that, maximum PCV (%) in two seasons was shown in the traits, 1st pod height (39.64 and 41.03%) and number of nodes/plant traits (45.41 and 39.64%). The differences between phenotypic and genotypic coefficient of variation (DZ) were low for all studied traits in both seasons (2017 and 2018) where it ranged from (0.11) in number of grains/plant trait to (1.74) in 1st pod height trait for the 1st year and ranged from (0.11) in 1000-grain weight trait to (3.31) in 1st pod height for the 2nd season. The lowest values of (DZ) were showed in the traits, number of grains/panicle (0.11 and 0.37) and 1000-grain weight (0.16 and 0.11), but the maximum level of (DZ) in both seasons was observed in 1st pod height trait (1.74 and 3.31), respectively. Data assessment of expected genetic advance (GA) based on 5% selection confirmed that the lowest level of genetic advance through both years was observed in grain yield/plant trait (2.66 and 2.54). While, the highest values during the 2 seasons were shown in 1st pod height trait (9.89 and 8.30) and number of nodes/plant trait (11.85 and 10.86) and the rest traits were detected medium to low results in this regard, respectively. Appreciates of genetic advance as percentage of mean (GAM (%)) varied from (2.71%) in plant height trait to (88.36%) in number of nodes/plant trait for the 1st season and ranged from (2.03%) in 1000-grain weight trait to (78.12%) in number of nodes/plant for the second season. The highest rank of (GAM%) in both seasons was shown in grain yield/plant trait (11.57 and 10.87%), first pod height trait (74.64 and 71.36%) and number of nodes/plant trait (88.36 and 78.12%), respectively.
Simple phenotypic correlation coefficient: Results obtained in Table 6 and 7 showed that highly significant positively of phenotypic correlation coefficients was observed among grain yield/plant and each of the following traits, plant height, 1st pod height, number of pods/plant, number of nodes/plant, number of grains/plant and 1000-grain weight during the 2 seasons (2017 and 2018), respectively.
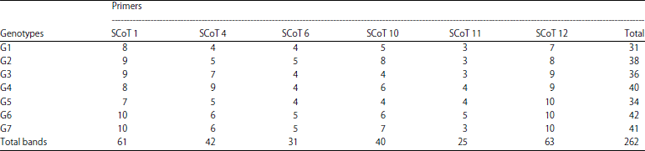
Molecular characterization: The profile of SCoT analysis was viewed in Table 8 and Fig. 1. Data detected that 55 amplicons were produced from the 6 SCoT primers where 27 of them were monomorphic and the rest (28) were polymorphic with 50.91% polymorphism including 13 unique bands with range size 115-831 bp. The first primer SCoT 1 detected 11 bands (6 of them were monomorphic and 5 polymorphic) with 45.45% polymorphism with sizes ranged from 193-816 bp. While, SCoT 4 primer produced 12 fragments (4 monomorphic and 8 polymorphic with 66.66% polymorphism) including 6 unique bands and the range size was from 253-831 bp. The third primer SCoT 6 showed 6 bands (4 of them were monomorphic and 2 polymorphic with 33.33% polymorphism) including one unique band and the range size was 275-477 bp. For SCoT 10 primer, there were 10 fragments (3 of them were monomorphic and 7 polymorphic included 4 unique bands), polymorphism percentage was 70.0% and the range size was 256-683 bp. While, primer SCoT 11 generated 5 fragments where (3 of them were monomorphic and 2 polymorphic with 40.0% polymorphism) besides one unique band and range sizes were ranged from 395-619 bp. Primer SCoT 12 produced 11 amplicons (7 monomorphic and 4 polymorphic including one unique band) with 36.36% polymorphism and range size was 115-138 bp, respectively. Results in Table 9 confirmed that genotype one (PR-162-11) recorded the lowest number of amplified fragments (31). While, the genotype number 6 (Giza 82) exhibited the highest number of amplified fragments (42) and the rest accessions revealed various number of amplified fragments. Primer SCoT 11 displayed the lowest number of fragments (25) for all entries. While that, Primer SCoT 12 produced the highest number of amplicons (63) for the same genotypes under studying.
| Table 4: | Mean performance and combined analysis for all studied traits of the seven soybean genotypes in the two seasons (2017 and 2018) |
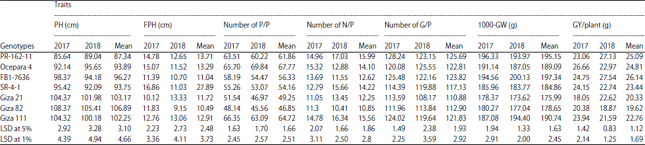 | |
| PH: Plant height, FPH: First pod height, P/P: Pods/plant, N/P: Nodes/plant, G/P: Grains/plant, GW: 1000-grain weight, GY/P: Grain yield/plant (g) | |
| Table 5: | Estimates of some genetic parameters for all studied traits in soybean genotypes under the two seasons (2017 and 2018) |
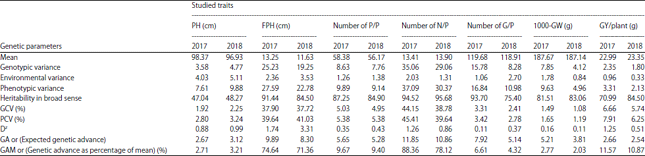 | |
GCV (%): Genotypic coefficient of variance percentage, PCV (%): Phenotypic coefficient of variance percentage, Dz: Difference between the phenotypic coefficient of variation (PCV (%)) and genotypic coefficient of variation (GCV (%)), PH: Plant height, FPH: First pod height, P/P: Pods/plant, N/P: Nodes/plant, G/P: Grains/plant, GW: 1000-grain weight, GY/P: Grain yield/plant (g) | |
| Table 6: | Simple phenotypic correlation coefficients among all studied traits in soybean genotypes for season (2017) |
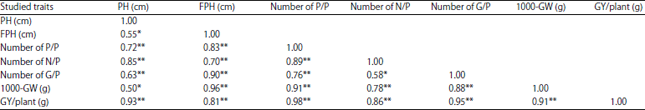 | |
| *Significant at 5% probability level, **Significant at 1% probability level, PH: Plant height, FPH: First pod height, P/P: Pods/plant, N/P: Nodes/plant, G/P: Grains/plant, GW: 1000-grain weight, GY/P: Grain yield/plant (g) | |
| Table 7: | Simple phenotypic correlation coefficients among all studied traits in soybean genotypes for season (2018) |
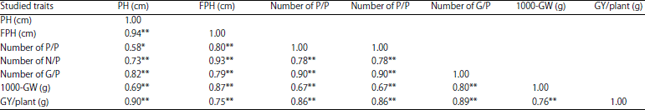 | |
| *Significant at 5% probability level, **Significant at 1% probability level, PH: Plant height, FPH: First pod height, P/P: Pods/plant, N/P: Nodes/plant, G/P: Grains/plant, GW: 1000-grain weight, GY/P: Grain yield/plant (g) | |
| Table 8: | Polymorphic loci amplified by the 6 SCoT primers for the 7 soybean genotypes |
 | |
| Table 9: | Total bands produced from each primer for 7 soybean accessions and all amplified fragments in each genotype |
 | |
Data shown in Table 10 showed that one negative specific marker was observed by SCoT 1 primer in genotype number 5 at size 519 bp. While, primers SCoT 6 and SCoT 11 produced one positive specific marker for each of them in genotype number 6 at molecular weights of 388 and 828 bp in addition, only one positive marker was generated by SCoT 12 primer for genotype 7 with size 333 bp, respectively. For SCoT 4 primer, 7 specific markers were generated by this primer as follows; one negative marker with size 347 bp was showed in genotype one, two positive markers with sizes 312 bp and 831bp were observed in genotype 3, three positive markers with sizes 296, 309 and 659 bp observed in genotype 4 besides one positive marker with size 470 bp was shown in genotype 7, respectively. The SCoT 10 primer displayed four positive and one negative specific markers as follows (two positive markers with sizes 617 and 683 bp in genotype 2, two positive markers with sizes 515 and 603 bp in genotype 7 and one negative marker only in genotype 5 at molecular weight of 322 bp), respectively.
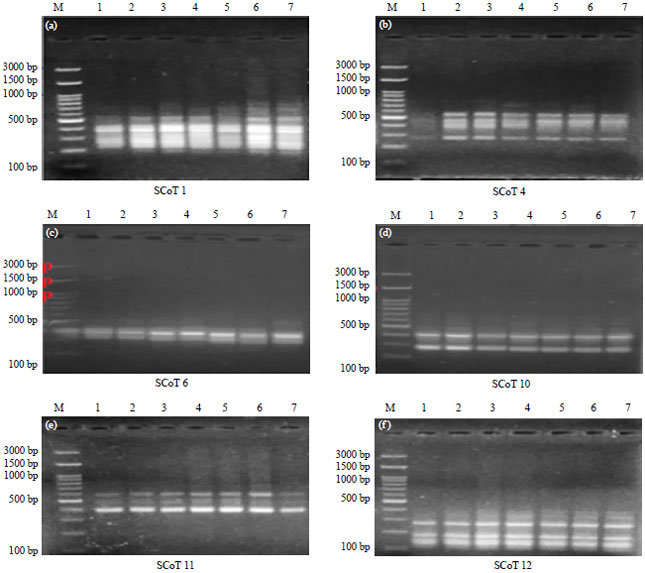 | |
| Fig. 1(a-f): | PCR fragments with 6 SCoT primers, (a) SCoT 1, (b) SCoT 4, (c) SCoT 6, (d) SCoT 10, (e) SCoT 11 and (f) SCoT 12 of 7 soybean genotypes (1:7) |
| 1: PR-162-11, 2: Ocepara 4, 3: FB1-7636, 4: SR-4-1, 5: Giza 21, 6: Giza 82 and 7: Giza 111; M: DNA Ladder as marker and molecular weights as follows: 100, 200, 300, 400, 500, 600, 700, 800, 900, 1000, 1500 and 3000 bp, respectively | |
| Table 10: | Mapping of positive (P) and negative specific markers for the 7 soybean genotypes using 6 SCoT primers |
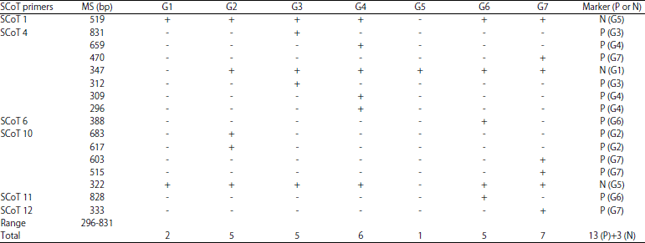 | |
P: Positive, N: Negative, MS: Molecular size, +: Present, -: Absent, G: Genotype | |
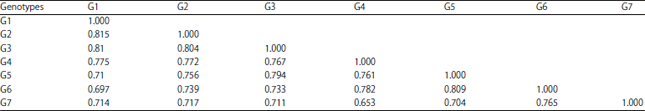
| Table 11: | Genetic similarity percentages of 7 soybean genotypes using 6 SCoT primers |
 | |
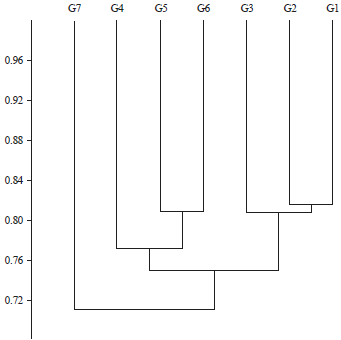 | |
| Fig. 2: | Dendrogram representing the genetic relationship among the 7 soybean accessions using UPGMA cluster analysis of Nei-Li’s similarity coefficient generated from the 6 SCoT markers |
Proximity matrix analysis (genetic similarity): Data presented in Table 11 showed 21 pairwise comparisons to debate the genetic relationships between 7 soybean accessions revealed in terms of similarity. The genetic similarity ranged from (0.653-0.815) with an average of 0.734, where, the lowest value of similarity was (0.653) among (G4 and G7) and the highest limit of genetic similarity was (0.815) within (G1 and G2). Other high genetic similarity values also for example not limited were observed among G2 and G3, G5 and G6, G1 and G3 and their values were (0.804, 0.809 and 0.810), respectively. The other results of the rest genetic similarities were observed also high.
Cluster analysis (phylogenetic tree): Results obtained from cluster analysis and shown in Fig. 2 consisted the 7 soybean genotypes to two main cluster. The cluster I divided into two sub-cluster where the first one included one sub-sub cluster contained one group (G1 and G2) and G3. Whereas, the second sub-sub cluster included G4 and (G5 and G6). Finally, the second cluster contained G7 only.
DISCUSSION
Results obtained in Table 3 detected that weak environmental impact and the extent of genetic stability for all soybean genotypes under studying during the two seasons. Analysis of variance confirmed the potential genetic variability and diversity in the seven soybean cultivars and this investigation was showed preferable scope for genetically enhancing and development by traditional breeding method which were agreement with those reported by authors4,15,16. Superior soybean genotypes briefly described in the item of results Table 4 and which exhibited highly limit for all studied traits have been demonstrated highly genetic stability during the two seasons. As well as their high yielding because it showed high level of plant height and this mechanism made it able to carry a large amount of pods/plant which has a great reflection on increasing of other studied traits such as first pod height, number of nodes/plant, number of grains/plant and 1000-grain weight which was eventually given the highest output. Therefore, the fruitful feasibility of this study is to ensure the efficiency of imported soybean accessions and evaluate them besides the most powerful Egyptian varieties. In this case plant breeders have a strong and new genetic base at the same time to be included in the breeding program to improve the Egyptian varieties through the process of direct hybridization by transferring required traits such as high yielding and resistance for both types of stresses3,4,15,17. The appreciation of components for variability is very fruitful for any influential selection technique to development a population because selection for convenient entries depends on the amount of variation in all genotypes under testing. Heritability in broad sense was appeared high in all studied traits in both seasons except plant height trait where it appeared medium for the 2 years (Table 5). This confirmed that low environment impact in the first case of increasing heritability on these traits in addition, the genetic variation was the greatest part of phenotypic variation. In the same regard, the fruitful and affective role of additive gene action was strongly visible for inheriting and improvement these traits for the previous soybean genotypes through simple selection process. While, in the second case of medium limit of heritability in broad sense for plant height trait during two seasons revealed the medium effect for each of environment and genetic variation which indicated that this trait might be governed by non-additive gene action besides the important role of the interaction among environment and genotype. In the fact, PCV (%) was always higher than GCV (%) in all traits under studying for both seasons indicating that the viewed variation was not only due to genotype but also depend on the significant impact of environment and the selection process for these traits. On the other hand, the gap between PCV (%) and GCV (%) was very little for all studied traits in both years which indicated that environmental impact was very weak on the expression of these traits and this reflects the extent of highly genetic stability for these accessions. Thus, selection processes for the development of the previous studied traits through phenotype would be the biggest goal in the present soybean accessions (Table 5). Similar results were obtained by Jain et al.18, Neelima et al.19, Mesfin20 and Nilahayati et al.21. High trend observed in genetic advance (GA) of both seasons for traits, number of nodes/plant and first pod height traits besides medium data of it for plant height, number of pods/plant, number of grains/plant and 1000-grain weight may be confirmed the fruitful role of additive gene action for controlling the expression of these traits. In the other side, this result indicated that the genetic improvement for these traits could be emotional by simple selection for individual plant which reflected to enhance the recent seven soybean materials. On the other hand, decreasing values of genetic advance (GA) in grain yield/plant trait in both years might be governed by non-additive gene action and the interaction among genotype and environment on the expression of this trait confirmed the weak role of individual plant selection for enhancing this trait in the previous materials. Thus, increasing limits of heritability, GCV (%) and (GA) in some traits may be governed by additive gene action and in the case of decreasing these parameters on the rest traits perhaps controlling by non-additive gene action. Regarding of GAM or (Genetic advance as percentage of mean (%)), high values observed in number of nodes/plant, first pod height and grain yield/plant traits could be confirmed the fruitful role of additive gene action for controlling and inheriting the previous traits. These results were in agreement with those reported by Hamawaki et al.2, El-Mouhamady et al.3, Chandrawat et al.4, Tawfik and El-Mouhamady16, Abou El-Nasr et al.22. Shoaib et al.23. El-Demardash et al.24 and Khatab et al.25. Data of phenotypic correlation coefficient through the 2 years and related to grain yield/plant besides the other traits under investigation confirmed firstly results were very similar during the 2 study years. Secondly, the progress achieved by the integration of the rest other traits and their collective contribution reflected on increasing of the final yielding. This is the positive trend and required in the way of genetic improvement desired for all quantitative traits responsible for increasing yield and its components. Therefore, the actual increasing in the values of the rest studied traits must have a positive role in increasing the final output (Table 6, 7). Similar results were agreement with those reported by El-Demardash et al.24, Malik et al.26, El-Mouhamady27, Sarutayophat28, Shree et al.29 and Vu et al.30. Results of SCoT analysis profile in Table 8 which showed the highest number of fragments and unique bands or positive specific markers in this study can be arranged in ascending order as follows (5 fragments and one unique band generated by SCoT 11 primer. Six bands and one unique band generated by SCoT 6 primer, 10 amplicons and 4 unique bands generated by SCoT 10 primer. The SCoT 1 and 12 primers were exhibited (11) fragments for each of them and number of unique band were (0 and 1) for each of them and 12 bands including 6 unique bands were generated by SCoT 4 primer, respectively. On the same regard, the lowest polymorphism (%) was shown in primer SCoT 6 (33.33%). While, the biggest level of polymorphism (%) was observed in primer SCoT 10 (70%), respectively. The highest number of amplified fragments were obtained in primers SCoT 1 (61) and SCoT 12 (63). This result indicated that these two primers have already succeeded in finding taxonomic differences at the genetic molecular level which had the greatest effect in comparing among accessions under studying based on agro-morphological traits and genetic parameters (Table 9). This mechanism will determine the most appropriate of these genotypes using in the Egyptian Soybean improvement program by transmitting important quantitative traits through hybridization. Similar results were in agreement with those obtained by El-Mouhamady et al.3, Al-Kordy et al.17, Heiba et al.31,32, Eldessouky et al.33, El-Mouhamady et al.34, Esmail et al.35, Ramadan et al.36, Khatab et al.37 and El-Mouhamady et al.38. Molecular markers have been very successful in drawing mapping consisting of 13 positive specific markers in addition, 3 negative markers that had the greatest effect in comparing among the seven soybean genotypes (Table 10). Therefore, these specific markers are considering as a genetic taxonomic basis at the molecular level in this regard. It was necessary identify and know these molecular genetic differences between these soybean accessions before using and introducing it in the soybean improvement program. These results were in agreement with those reported by El-Mouhamady et al.3, Al-Kordy et al.17, Khatab et al.37 and El-Mouhamady et al.38. Results presented in Table 11 and related to the genetic similarity confirmed that the relationships among (G1 and G2), (G1 and G3), (G2 and G3) and (G5 and G6) recorded the best genetic similarity in this regard and revealed that (G1, G2, G3, G5 and G6) considering very genetically closed from each other. Therefore, these materials can be introduced in breeding programs to enhance and improve an important traits of soybean crop under local conditions. These results were in agreement with those reported by Khatab et al.25, Esmail et al.35, Ramadan et al.36, Khatab et al.37, El-Mouhamady et al.38 and El-Mouhamady and Habouh39. Cluster analysis viewing in Fig. 2 showed the degree of genetically convergence or divergence between the seven soybean varieties. This fact gives an important reflection on the actual relationship between the Egyptian and imported varieties of soybean. This may help breeders and researchers to know the extent of environmental and genetic suitability within these materials before introducing it in the soybean breeding and improvement program to convey some important quantitative traits such as high yield, water stress resistance, salinity tolerance and other strategic traits in breeding program. These results were in agreement with those reported by Khatab et al.25, Esmail et al.35, Ramadan et al.36, Khatab et al.37, El-Mouhamady et al.38 and El-Mouhamady and Habouh39. After a quick and interesting review of all contents for this study can be very briefly clarify the fruitful recommendation of this work. It is introducing of imported soybean genotypes into the Egyptian national program for breeding and improving soybean crop traits by transferring the excellent characters necessary to increase yielding and resistance of abiotic and biotic stresses from those imports.
CONCLUSION
The present investigation was conducted to study the genetic variability in some imported soybean accessions besides 3 Egyptian cultivars during two seasons including agro-morphological traits. Molecular characterization for all materials was done using 6 SCoT primers. Analysis of results was given moral and excellent evidence for the extent of genetic stability based on all data obtained from the genetic parameters. Molecular markers analysis profile showed the prospect of distinctiving, recognizing, selection process of genotypes which improvement by using this method.
SIGNIFICANCE STATEMENT
This study succeed for introducing new promising soybean accessions with highly yielding and resistance for abiotic and biotic stresses in Egyptian agriculture. This strategy can be beneficial for enhancing and improving Egyptian soybean crop through utilizing them in a hybridization program with imported soybean accessions to convey important traits mentioned above. This study also will help the researchers to identify specific markers which will be the taxonomic basis at the molecular level for these soybean genotypes. Thus, a new theory for discovering anew soybean genotypes with highly yielding and tolerance for many biotic and abiotic stresses in addition, the great benefit in the national program to improve the Egyptian soybean crop may be arrived at.
REFERENCES
- Malik, M.F.A., A.S. Qureshi, M. Ashraf and A. Ghafoor, 2006. Genetic variability of the main yield related characters in soybean. Int. J. Agric. Biol., 8: 815-819.
Direct Link - Hamawaki, O.T., L.B. de Sousa, F.N. Romanato, A.P.O. Nogueira, C.D.S. Júnior and A.C. Polizel, 2012. Genetic parameters and variability in soybean genotypes. Comunicata Scientiae, 3: 76-83.
Direct Link - El-Mouhamady, A.A., H.M. Abdel-Rahman, T.A. Elewa and K.A. Aboud, 2017. Molecular characterization and stability analysis for yield and its components traits in soybean (Glycine max L.). Int. J. Curr. Microbiol. Applied Sci., 6: 1835-1857.
CrossRefDirect Link - Chandrawat, K.S., K.S. Baig, S. Hashmi, D.H. Sarang, A. Kumar and P.K. Dumai, 2017. Study on genetic variability, heritability and genetic advance in soybean. Int. J. Pure Applied Biosci., 5: 57-63.
CrossRefDirect Link - Zhao, H., Y. Wang, F. Xing, X. Liu and C. Yuan et al., 2018. The genetic diversity and geographic differentiation of the wild soybean in Northeast China based on nuclear microsatellite variation. Int. J. Genom., Vol. 2018.
CrossRefDirect Link - Koutu, G.K., A. Shrivastava, Y. Singh and S. Tiwari, 2019. Molecular characterization and genetic diversity assessment of soybean varieties using SSR markers. Int. J. Curr. Microbiol. Applied Sci., 8: 173-182.
CrossRefDirect Link - Li, F., X. Liu, S. Wu, Q. Luo and B. Yu, 2019. Hybrid identification for Glycine max and Glycine soja with SSR markers and analysis of salt tolerance. PeerJ., Vol. 7.
CrossRefDirect Link - Burton, W.G. and E.H. Devane, 1953. Estimating heritability in tall fescue (Festuca arundinacea) from replicated clonal material. Agron. J., 45: 478-481.
CrossRefDirect Link - Johnson, H.W., H.F. Robinson and R.E. Comstock, 1955. Estimates of genetic and environmental variability in soybeans. Agron. J., 47: 314-318.
CrossRefDirect Link - Miller, P.A., J.C. Williams, H.F. Robinson and R.E. Comstock, 1958. Estimates of genotypic and environmental variances and covariances in upland cotton and their implications in selection. Agron. J., 50: 126-131.
CrossRef - Williams, J.G.K., A.R. Kubelik, K.J. Livak, J.A. Rafalski and S.V. Tingey, 1990. DNA polymorphisms amplified by arbitrary primers are useful as genetic markers. Nucleic Acids Res., 18: 6531-6535.
CrossRefPubMedDirect Link - Jaccard, P., 1908. Nouvelles recherches sur la distribution florale. Bull. Soc. Vaud. Sci. Nat., 44: 223-270.
CrossRefDirect Link - Yang, X. and C. Quiros, 1993. Identification and classification of celery cultivars with RAPD markers. Theor. Applied Genet., 86: 205-212.
CrossRefDirect Link - Baraskar, V.V., V.H. Kachhadia, J.H. Vachhani, H.R. Barad, M.B. Patel and M.S. Darwankar, 2014. Genetic variability, heritability and genetic advance in soybean [Glycine max (L.) Merrill]. Electron. J. Plant Breed., 5: 802-806.
Direct Link - Tawfik, R.S. and A.B.A. El-Mouhamady, 2019. Molecular genetic studies on abiotic stress resistance in sorghum entries through using half diallel analysis and Inter-Simple Sequence Repeat (ISSR) markers. Bull. Natl. Res. Centre, Vol. 43.
CrossRefDirect Link - Al-Kordy, M.A., H.F. Ibrahim, A.B.A. El-Mouhamady and H.M. Abdel-Rahman, 2019. Genetic stability analysis and molecular depiction in elite entries of rice (Oryza sativa L.). Bull. Natl. Res. Centre, Vol. 43.
CrossRefDirect Link - Jain, R.K., A. Joshi, H.R. Chaudhary, A. Dachoura and C.L. Khatik, 2018. Study on genetic variability, heritability and genetic advance in Soybean [Glycine max (L.) Merrill]. Leg. Res. Int. J., 41: 532-536.
CrossRefDirect Link - Neelima, G., S.P. Mehtre and G.W. Narkhede, 2018. Genetic variability, heritability and genetic advance in soybean. Int. J. Pure Applied Biosci., 6: 1011-1017.
CrossRefDirect Link - Mesfin, H.H., 2018. Path analysis, genetic variability and correlation studies for soybean (Glycine max (L.) Merill) for grain yield and secondary traits at Asosa, Western Ethiopia. Greener J. Plant Breed. Crop Sci., 6: 35-46.
CrossRefDirect Link - Nilahayati, Rosmayati, D.S. Hanafiah and F. Harahap, 2018. Genetic variability and heritability on Kipas Putih soybean mutant lines using gamma rays irradiation (M3 generation). IOP Conf. Ser.: Earth Environ. Sci., Vol. 122.
CrossRefDirect Link - Abou El-Nasr, T.H.S., M.M. Ibrahim, K.A. Aboud and M.A. El-Enany, 2013. Assessment of genetic variability for three coriander (Coriandrum sativum L.) cultivars grown in Egypt, using morphological characters, essential oil composition and ISSR markers. World Applied Sci. J., 25: 839-849.
Direct Link - Shoaib, R.M., N.S. Abdel-Samea, W.A. Ramadan, M.M. Ibrahim and K.A. Aboud, 2016. Elucidation of genetic parameters among some selected genotypes of prickly oil lettuce (Lactuca serriolla L.) in Egypt, using morpho-agronomic traits and RAPD markers. Res. J. Pharm. Biol. Chem. Sci., 7: 1255-1264.
Direct Link - El-Demardash, I.S., A.A. El-Mouhamady, H.M. Abdel-Rahman, T.A. Elewa and K.A. Aboud, 2017. Using gamma rays for improving water deficit tolerance in rice. Curr. Sci. Int., 6: 321-327.
Direct Link - Khatab, I.A., A.B.A. El-Mouhamady, S.A. Mariey and T.A. Elewa, 2019. Assessment of water deficiency tolerance indices and their relation with ISSR markers in barley (Hordeum vulgare L.). Curr. Sci. Int., 8: 83-100.
Direct Link - Malik, M.F.A., M. Ashraf, A.S. Qureshi and A. Ghafoor, 2007. Assessment of genetic variability, correlation and path analyses for yield and its components in soybean. Pak. J. Bot., 39: 405-413.
Direct Link - Sarutayophat, T., 2012. Correlation and path coefficient analysis for yield and its components in vegetable soybean. Songklanakarin J. Sci. Technol., 34: 273-277.
Direct Link - Shree, Y., S. Ram, S. Bhushan, N. Verma, E. Ahmad and S. Kumar, 2018. Correlation between yield and yield attributing traits in soybean (Glycine max (L.) Merrill). J. Pharmacogn. Phytochem., 7: 298-301.
Direct Link - Vu, T.T.H., T.T.C. Le, D.H. Vu, T.T. Nguyen and T. Ngoc, 2019. Correlations and path coefficients for yield related traits in soybean progenies. Asian J. Crop Sci., 11: 32-39.
CrossRefDirect Link - Heiba, S.A.A., A.A. El-Mouhamady, S.E. Eldessouky, H.B. Ali and T.A. Elewa, 2016. Study the genetic variations related to the resistance of heavy metals toxicity in some rice genotypes using RAPD markers. Int. J. Curr. Microbiol. Applied Sci., 5: 174-189.
CrossRefDirect Link - Heiba, S.A.A., S.E.I. Eldessouky, A.A. El-Mouhamady, I.S. El-Demardash and A.A. Abdel-Raheem, 2016. Use of RAPD and ISSR assays for the detection of mutation changes in wheat (Triticum aestivum L.) DNA induced by Ethyl-Methane Sulphonate (EMS). Int. J. Chem. Tech. Res., 9: 42-49.
Direct Link - Eldessouky, E.I., S.A.A. Heiba, A.A. El-Mouhamady and Y.M. Abdel-Tawab, 2016. DNA fingerprinting and half diallel analysis of some rice genotypes under water deficit conditions. Res. J. Pharmaceut. Biol. Chem. Sci., 7: 985-997.
Direct Link - El-Mouhamady, A.A., Z.M. El-Ashary, F.I. Mohamed, T.A. Elewa and K.A. Aboud, 2016. Study the effect of water stress conditions on some genotypes of bread wheat (Triticum aestivum L.) based on morphological, physiological traits and DNA fingerprinting. Res. J. Pharmaceut. Biol. Chem. Sci., 7: 2065-2077.
Direct Link - Esmail, R.M., A.A. Abdel Sattar, S. Abdel-Samea, A.A. El-Mouhamady, E.M. Abdelgany and F.B. Fathallaha, 2016. Assessment of genetic parameters and drought tolerance indices in maize diallel crosses. Res. J. Pharmaceut. Biol. Chem. Sci., 7: 2409-2428.
Direct Link - Ramadan, W.A., H.M. Abdel-Rahman, A.A. El-Mouhamady, M.A.F. Habouh and K.A. Aboud, 2016. Molecular genetic studies on some barley entries for drought tolerance. Int. J. PharmTech Res., 9: 265-285.
Direct Link - Khatab, I.A., A.A. El-Mouhamady, H.M. Abdel-Rahman, M.A. Farid and I.S. El-Demardash, 2017. Agro-morphological and molecular characterization of sorghum (Sorghum vulgare L.) for water stress tolerance. Int. J. Curr. Res. Biosci. Plant Biol., 4: 37-55.
CrossRefDirect Link - El-Mouhamady, A.B.A., H.M. Abdel-Rahman, A.A. Rizkalla and M.A. El-Metwally, 2019. Assessment of water stress tolerance in wheat genotypes based on half diallel analysis and DNA fingerprinting. Pak. J. Biol. Sci., 22: 103-116.
CrossRefDirect Link - El-Mouhamady, A.AB.A. and M.A.F. Habouh, 2019. Genetic improvement of some rice genotypes for salinity tolerance using generation mean analysis. Curr. Sci. Int., 8: 321-348.
Direct Link