
J.J. Jolly
Horseshoe Crab Research Group, Institute of Marine Biotechnology, Universiti Malaysia Terengganu, 21030 Kuala Nerus, Terengganu, Malaysia
S.K. Dzulkiply
Horseshoe Crab Research Group, Institute of Marine Biotechnology, Universiti Malaysia Terengganu, 21030 Kuala Nerus, Terengganu, Malaysia
M.A. Yusof
Horseshoe Crab Research Group, Institute of Marine Biotechnology, Universiti Malaysia Terengganu, 21030 Kuala Nerus, Terengganu, Malaysia
N.A. Kamaruding
Horseshoe Crab Research Group, Institute of Marine Biotechnology, Universiti Malaysia Terengganu, 21030 Kuala Nerus, Terengganu, Malaysia
N. Ismail
Horseshoe Crab Research Group, Institute of Marine Biotechnology, Universiti Malaysia Terengganu, 21030 Kuala Nerus, Terengganu, Malaysia
LiveDNA: 60.26983
Pakistan Journal of Biological Sciences
Year: 2019 | Volume: 22 | Issue: 9 | Page No.: 427-434







ABSTRACT
Background and Objectives: Hemocyanin Subunit IIIA is believed to possess antimicrobial properties, but its efficacy against microbial pathogens is still unclarified. Thus, this study aimed to determine antimicrobial activities of hemocyanin subunit IIIA and to identify the best activator of this protein. Materials and Methods: The hemocyanin was partially purified using spin column affinity, its fraction was applied to Hi-Prep Sephacryl Exclusion 26/60 2-200 HR column, followed by Hi-Prep 26/10 Desalting Column on fast protein liquid chromatography. The purity of hemocyanin was validated by Matrix Assisted Laser Desorption Ionization-Time of Flight/Mass Spectrometry. The antimicrobial activity was performed by Disc Diffusion Test. Results: Purified hemocyanin subunit IIIA was identified to have a molecular weight of 72.9 kDa. SDS was found to be the best activator of hemocyanin, as indicated by elevated level of phenoloxidase. As for antimicrobial activity, hemocyanin was minimally inhibited by all bacteria strains tested (Escherichia coli, Staphylococcus aureus and Klebsiella pneumoniae), with relatively lower Minimum Inhibitory Concentration (MIC) at 0.005 g mL–1, than recorded MIC for fungal test strains. Two fungal strains (Penicillium sp. and A. niger) show susceptible response to phenoloxidase using MgSO4 as inducer. Whereas, lysate-treated CaCl2 induced susceptibility only to A. niger. Conclusion: Hemocyanin shows better antimicrobial activity than phenoloxidase because of its broad-spectrum activity against bacterial and fungal strains tested. Hence, the hemocyanin may potentially become a new antimicrobial candidate to be discovered for a future use in treatment of resistant bacteria.
PDF Abstract XML References Citation
Copyright: © 2019. This is an open access article distributed under the terms of the creative commons attribution License, which permits unrestricted use, distribution and reproduction in any medium, provided the original author and source are credited.
How to cite this article
J.J. Jolly, S.K. Dzulkiply, M.A. Yusof, N.A. Kamaruding and N. Ismail, 2019. Broad-spectrum Antimicrobial Activity of Purified Hemocyanin Subunit IIIA Isolated from Asian Horseshoe Crab, Tachypleus gigas. Pakistan Journal of Biological Sciences, 22: 427-434.
DOI: 10.3923/pjbs.2019.427.434
URL: https://scialert.net/abstract/?doi=pjbs.2019.427.434
DOI: 10.3923/pjbs.2019.427.434
URL: https://scialert.net/abstract/?doi=pjbs.2019.427.434
INTRODUCTION
In the 21st century of modern-era, antibiotic resistance has become a global issue of concern. A concern arises over the resistance towards a commonly reported bacteria such as Escherichia coli, Klebsiella pneumoniae, Staphylococcus aureus and Streptococcus pneumoniae and also bacteria associated with life-threatening diseases such as tuberculosis, malaria, HIV and influenza1-3, because they can easily transform to virulent strains. The scenario is more alarming when the development and approval of new antibiotic has declined steadily over the past few decades, leaving a few option for treating resistant bacteria4,5.
Hemocyanin, a respiratory protein found in horseshoe crabs’ hemolymph plasma plays several roles such as oxygen carrier, storage protein, buffers and osmolytes as well as carrier of molting hormones6. In view of its structural-classification, the hemocyanin and phenoloxidase are known to possess a highly preserved bi-copper binding-site; wherein they are classified as type-3 copper of oxygen-binding protein7,8. Both of these proteins share similar physicochemical properties and pattern of bi-copper active site9-11, but their biological-functions are physiologically different. Activation of hemocyanin to phenoloxidase can be stimulated by in vitro effectors, which are usually denaturing agents such as detergents, salts and alcohols12,13. SDS is a common detergent used to unfold and expose the active site to phenolic substrates to bind with hemocyanin14,15. The hemocyanin from arthropod and molluscs has been found to give a good inhibition against microbial pathogens16,17. To date, it is still remained unclear on the mechanism of hemocyanin acts as antimicrobial agent in both inactivated and activated forms in Asian horseshoe crab, T. gigas. Therefore, this study aimed to determine antimicrobial activities of new isolated purified hemocyanin subunit IIIA, which are believed to be the first report on T. gigas and to identify the best activator of this protein to its derivative (phenoloxidase).
MATERIALS AND METHODS
Blood collection of Tachypleus gigas: Fifty live horseshoe crabs, T. gigas were collected at two respective locations: Chendor (4°9'37"N 103°24'3"E) and Cherating (4°6'0"N, 103°23'0"E), between April-September 2015, whereby both are located at State of Pahang, Malaysia. Approximately, 15-25 mL blood was extracted at 30% of total body weight of the animals. Protocol of horseshoe crab blood extraction was conducted based on the method described by Noraznawati et al.18.
Partial purification of hemocyanin: Hemocyanin was purified using Spin Column Nanosep by separating proteins using two types of molecular weight cut-off (MWCO) filters, each at 30 and 100 K (Pall Life Sciences, Mexico) following method described by Wang et al.19 and Hangel et al.20, with some modifications. Briefly, 50 μL hemolymph sample at concentration of 15 g mL–1 was pipetted into the sample reservoir using 30K MWCO filter, followed by centrifugation at 14,000×g for 10 min. The solution that remained in the reservoir was taken for the next purification procedure using 100K MWCO filter. Then, the solution at the bottom-phase was kept for further use in the next purification using fast protein liquid chromatography ÄKTA Purifier (FPLC) (GE Healthcare, UK). Next, the hemocyanin filtrate was suspended into suspension buffer (50 mM HEPES, 1 mM NaCl, 159 mM EDTA and 1% Triton X-100, adjusted at pH 7.5). Subsequently, gel filtration was performed using Hi-prep Sephacryl Size Exclusion 26/60 2-200 HR Column Chromatography (GE-Healthcare, Sweden) and pre-equilibrated with the same buffer. After filtration, the sample was subjected to desalting process using Hi-PrepTM 26/10 Desalting Column (GE-Healthcare, Sweden), with bed dimension of 26 mm (width)×100 mm (length). In the desalting process, the column was first equilibrated by injecting 50 mM sodium phosphate buffer at pH 7, containing 0.15 M NaCl at a flow-rate of 15 mL min–1. Finally, 10 mL of hemocyanin filtrate was injected into FPLC. The eluant containing hemocyanin was kept at -20°C until further use.
Protein profiling using sodium dodecyl sulfate polyacrylamide gel electrophoresis (SDS-PAGE): SDS-PAGE was performed using the method described by Laemmli21. The molecular weight of purified hemocyanin was determined using 12% resolving gel. The gel was then run at 100 volts for 1 h. After electrophoresis, protein bands were stained using 0.05% Coomassie Brilliant Blue R-250 (Bio-Rad, US).
Identification of protein using matrix assisted laser desorption/ionization time of flight mass spectrometry (MALDI-TOF/MS): The purified sample was subjected to MALDI-TOF-MS, with the purpose to validate purity of the sample. After a distinct peak of the purified sample obtained, the peptide matching sequence was searched and identified through MASCOT database.
Quantification of phenoloxidase activity: Bioassay of phenoloxidase was performed following method described by Fan et al.22, with some modifications. Prior to the assay, hemocyanin was activated using 20 mM Tris-acetic acid (pH 7.5), containing 0.15 M NaCl and pre-incubated for 10 min at 20°C. As for reaction with metal ions, reaction buffer comprised of 20 mM Tris-HCl buffer at pH 7.1 was incubated for 20 min at 40°C. The phenoloxidase activity was determined by measuring the formation of ortho-quinone from reduction of catechol. The gradient of color changes produced from this reaction were measured by reading an absorbance at 560 nm using Glomax Multi Detection System (Omega, US). An induction of phenoloxidase was performed by using the same mixture, substituting the hemolymph lysate supernatant with 50 mM of respective protein cleavage inducers namely EDTA, citric acid, MgSO4, CaCl2, SDS and thiourea.
Determination of antimicrobial activity of hemocyanin and phenoloxidase using disc-diffusion test (DDT): DDT assay of hemocyanin and phenoloxidase were performed following method described by Reller et al.23, with some modifications. For anti-bacterial activity, 4 bacteria strains [Escherichia coli (PC64), Staphylococcus aureus (PC152) and Klebsiella pneumoniae (PC135)] were inoculated on Muller-Hinton agar. Then, ten drops of respective samples (hemocyanin and phenoloxidase) at concentration of 0.02 g mL–1 were pipetted on 6 mm paper-disc (GE Healthcare, UK), dried and placed on the agar plate. In subsequent step, the agar plates were incubated overnight at 37°C. The diameter of inhibition zone was measured and recorded in triplicates for each test strain. For negative control, metal ion and hemocyanin buffers were used. In addition, tetracycline (30 μg) acts as positive control. Three fungi strains were tested in the anti-fungal activities namely Saccharomyces cerevisiae, Penicillium sp. and Aspergillus niger. The fungi test strains were grown on potato dextrose agar. Similar protocol as used in the anti-bacterial activity was performed for anti-fungal activity. In the anti-fungal activity, metal ion and hemocyanin buffers were assigned as negative controls. Whereas, nystatin (100 μg) was used as positive control.
Statistical analysis: All of experimental data was calculated using simple formula performed in Microsoft Excel®, which was expressed as Mean±standard deviation.
RESULTS
Determination of elution peak and molecular weight of purified hemocyanin subunit IIIA: Purification of hemocyanin obtained in this study showing three major peaks consisted of peak A was identified as hemocyanin, peaks B and C were α2-macroglobulin and C-reactive protein, respectively (Fig. 1).
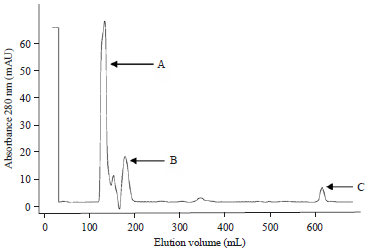 | |
| Fig. 1: | Elution diagram showing a purified hemocyanin from T. gigas |
Three major peaks on the diagram indicate A: Hemocyanin, B: α2-macroglobulin, C: C-reactive protein | |
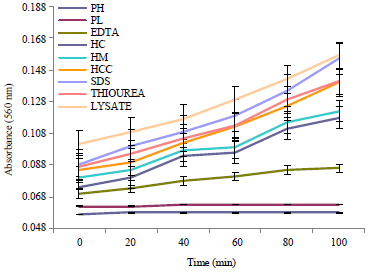 | |
| Fig. 2: | Conversion of hemocyanin into phenoloxidase by protease inducers |
PH: Pure hemocyanin (negative control), PL: Pure lysate (negative control), EDTA: HC+EDTA, HC: HC+citric acid, HM: HC+MgSO4, HCC: HC+CaCl2, SDS: HC+SDS, THIOUREA: HC+Thiourea, LYSATE: HC+ lysate (positive control) | |
The protein sequence was identified as hemocyanin subunit IIIA, having a molecular weight of 72.9 kDa and similarity of protein sequence with Carscinoscorpius rotundicauda and Limulus polyphemus.
Effect of protease inducers on enzymatic activity of phenoloxidase: Both pure hemocyanin and pure lysate that served as negative control in phenoloxidase activity showed a basal level throughout the incubation indicating no formation of phenoloxidase (Fig. 2).
| Table 1: | Anti-bacterial and anti-fungal susceptibility tests of hemocyanin subunit IIIA treated with various protease inducers |
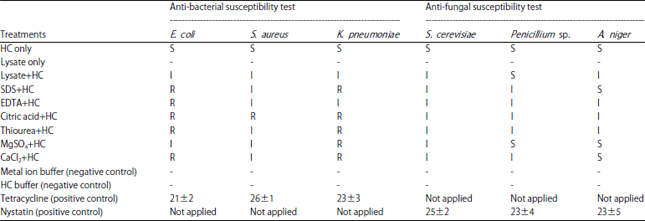 | |
Sources: CLSI24, CLSI25, Tokarzewski et al.26, HC: Hemocyanin, S: Susceptible, I: Intermediate, R: Resistant, -: No inhibition, anti-bacterial and anti-fungal susceptibility disc diffusion tests indicate E. coli and K. pneumoniae (R<11, <12, I<14, S>15), S. aureus (R<14, <15, I<18, S>19), Penicillium sp. and A. niger (R<14, I<18, S>18), S. cerevisiae (R<16, <17, I<25, S>26) | |
The activity of phenoloxidase was seen the highest using SDS as protease inducer, as marked by an increment pattern of absorbance value reaching maximal at 0.110, followed by thiourea, CaCl2, MgSO4, citric acid and EDTA, with respective absorbance values (0.108, 0.098, 0.096, 0.093 and 0.073) (Fig. 2).
Antimicrobial activity of inactivated and activated form of purified hemocyanin IIIA: Anti-bacterial susceptibility test showed that all bacteria strains tested are susceptible towards hemocyanin (Table 1). Activation of hemocyanin by lysate shows that all bacteria strains have intermediate response indicating broad spectrum activities. Both E. coli and K. pneumoniae displayed resistance response towards phenoloxidase through activation by SDS, thiourea and CaCl2. Meanwhile, E. coli was also resistant to phenoloxidase when induced by EDTA. EDTA-treated hemocyanin showed an intermediate response towards K. pneumoniae and S. aureus, suggesting broad spectrum activity. Only S. aureus showed intermediate response towards phenoloxidase using treatment of SDS, thiourea and CaCl2, indicating a specific spectrum activity only to Gram-positive bacteria. Moreover, all bacteria strains were resistant towards phenoloxidase, which initially treated by citric acid, indicating no spectrum activity. Additionally, phenoloxidase activated by MgSO4 showed intermediate response to both E. coli and S. aureus suggesting broad spectrum activities against Gram-positive and negative bacteria. Only K. pneumoniae was observed to give resistant inhibition towards activation hemocyanin by MgSO4.
Anti-fungal susceptibility test showed that all fungi test strains were susceptible towards hemocyanin (Table 1). On the other hand, all fungi strains tested show intermediate response towards phenoloxidase as activated by EDTA, citric acid and thiourea. Meanwhile, both Penicillium sp. and S. cerevisiae displayed intermediate response towards phenoloxidase by SDS and CaCl2. Aspergillus niger showed a susceptible response towards hemocyanin-treated by SDS and CaCl2. Both S. cerevisiae and A. niger showed intermediate response towards phenoloxidase by lysate as inducer. In contrast, Penicillium sp. shows susceptibility response towards phenoloxidase as activated by lysate. Meanwhile, hemocyanin-treated by MgSO4 shows susceptibility response towards Penicillium sp. and A. niger; whereas, S. cerevisiae shows only intermediate response.
It was observed that the MIC value of hemocyanin for the three bacteria strains tested was recorded at 0.005 g mL–1 (Table 2). A clear visible inhibition zones were observed for concentrations of 0.01, 0.02 and 0.04 g mL–1. In addition, no visible inhibition zone was observed for hemocyanin concentration at 0.0025 g mL–1. As for fungi, the MIC value of hemocyanin that able to kill S. cerevisiae was recorded at 0.01 g mL–1; whereas, for both Penicillium sp. and A. niger were inhibited at 0.02 g mL–1. At both hemocyanin concentrations (0.02 and 0.04 g mL–1), a clear visible inhibition zone was observed. It was further noticeable that hemocyanin did not inhibit fungal strains at concentration of 0.0025 g mL–1.
| Table 2: | MIC test of purified inactivated hemocyanin subunit IIIA against bacteria and fungi strains |
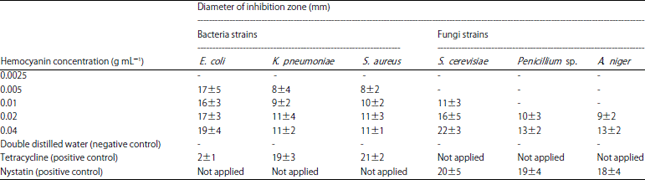 | |
DISCUSSION
Based on the DDT assay conducted in this study, inactivated hemocyanin shows broad spectrum activities by killing both gram-negative bacteria (E. coli and K. pneumoniae) and gram-positive bacteria (S. aureus). As for anti-fungal test, all fungi strains tested (S. cerevisiae, Penicillium sp. and A. niger) displayed susceptibility response towards inactivated hemocyanin. The broad-range spectrum of responses has been found by all fungi strains towards phenoloxidase. This findings in fact was supported with Coates and Talbot27, who demonstrated microbicidal effect of hemocyanin derived phenoloxidase in Limulus polyphemus towards Bacillus megaterium, B. subtilis, Saccharomyces cerevisiae, E. coli, Micrococcus luteus and Pantoea agglomerans. The reason behind the diminished number of colonies forming unit were associated with the toxicity of hemocyanin derived phenoloxidase reaction product upon activation using phenolic substrates (e.g., L-DOPA)27.
All the three tested bacteria strains have shown similar MIC value of 0.005 g mL–1 with observed diameter of inhibition zone for E. coli, K. pneumonia and S. aureus were at 17±5, 8±4 and 8±2 mm, respectively. In reference to Knowledge Base the Antimicrobial Index the tetracycline MIC values for both of E. coli and K. pneumoniae is within a range of 0.5-8 μg mL–1 and S. aureus is ranging from 2-8 μg mL–1. Therefore, MIC value of inactivated hemocyanin falls within the range of tetracycline activity, suggesting it is a potent anti-bacterial agent. The MIC value of S. cerevisiae was recorded at 10 μg mL–1, with diameter of inhibition zone was recorded at 11 mm. By comparison, both Penicillium sp. and A. niger demonstrated the same MIC value of 0.02 g mL–1, with diameter of inhibition zone at 10 and 9 mm, respectively. This finding was supported by Carrillo-Munoz et al.28, who found the general MIC value for anti-fungal activity for nystatin is ranging from 0.019-10 mg mL–1; therefore, the MIC activity of hemocyanin towards S. cerevisiae, Penicillium sp. and A. niger is less potent than nystatin as an anti-fungal agent.
Various in vitro and in vivo inducers have been proposed in the activation of hemocyanin to phenoloxidase. In chelicerates, such as tarantula, Schenk et al.29 found that SDS induces a strong phenoloxidase activity by inducing an optimal conformational transition in the protein which may result in opening a channel to the bi-copper center allowing bulky phenolic substrates access to catalytic-site. Moreover, Laino et al.30 reported that phenoloxidase activity is inducible in both phenoloxidase and hemocyanin by incubation with SDS. It is interesting to note that the SDS imitates the effects of natural activators such as fatty acids, phospholipids and small antimicrobial peptides and interacts with phenoloxidase and hemocyanin without inducing denaturation31. Other than SDS, Coates et al.32 suggested incubation of hemocyanin with anionic phospholipids phosphatidylserine and phosphatidylinositol showed a better activator of phenoloxidase, by inducing 80% of the activity than the neutral lyso-phosphatidylcholine and SDS-micelles.
pH also influences the oxygen-binding affinity of hemocyanin in arthropods33. This phenomenon has been explained by Sterner and Decker34 as “Bohr-effect” which is defined as capability of these proteins to respond to a pH variation in the medium with a different affinity of oxygen molecules in meeting physiological demands. In an acidic environment, the total charge of hemocyanin is affected, thus influencing the bonding μ: η2-η2 bridge between the two copper atoms35. According to Brouwer et al.36, at pH 7, calcium does not have any noticeable effect on oxygen binding by Limulus hemocyanin. The oxygen affinity of the Limulus hemocyanin decreases at high calcium concentration and this might explain the preference of functional to take-up of catechol. This explanation could also be applied to other divalent metal ions such as magnesium. The addition of thiourea to urea as an additional chaotropic agent has shown interesting features for protein solubilization but also to control protease activity37. Due to limited proteolytic activity by thiourea, conformational changes occurred resulting in moderate conversion of phenoloxidase.
The benefit of hemocyanin as anti-fungal agent is that it may not impose any detrimental effect on the environment and it is easily to metabolize38. Inactivated hemocyanin produces the highest antimicrobial activities compared to its activated form, the phenoloxidase. However, both activated and inactivated hemocyanin has shown better results for anti-fungal test as compared to anti-bacterial test. Moreover, hemocyanin displayed more inhibition activities on fungi surpassing with the inhibition on phenoloxidase. The mechanism of action hemocyanin as an antimicrobial agent is apparently influenced by the molecular structure because inducers used in this study disrupt the native conformation of the hemocyanin39. This could also suggest the phenoloxidase is not the only form of antimicrobial agent in vivo, but the native hemocyanin also plays a role in combating pathogenic invasions in situ in the horseshoe crab40. This however, needs further investigation in order to uncover the mechanism underlying this dichotomy.
CONCLUSION
SDS was found to have strong inducer in activating the phenoloxidase activity. Hemocyanin gave broad-spectrum antimicrobial activity than phenoloxidase as indicated by susceptibility responses to all bacteria (E. coli, S. aureus and K. pneumoniae) and fungal tested (S. cerevisiae, Penicillium sp. and A. niger). Hence, hemocyanin has a high potential to be used as antimicrobial candidate in combating resistant bacteria.
SIGNIFICANCE STATEMENT
This study discovered the broad-spectrum antimicrobial activity of purified hemocyanin subunit IIIA isolated from Asian horseshoe crab, T. gigas which has not yet been clarified and scarcely investigated, in comparison to antimicrobial properties claimed in American horseshoe crab (Limulus polyphemus) and other mollusc that can be beneficial for offering candidates of antibiotic drugs in resolving the emergence issue of antibiotics resistance occurring worldwide. This study will help the researchers to uncover the critical areas of revealing susceptibility of Hemocyanin Subunit IIIA to virulent strains of bacteria that gave better responses than its derivative (phenoloxidase), which found to have narrow antimicrobial responses that many researchers were not able to explore. Thus, a new theory on distinct antimicrobial properties between hemocyanin subunit IIIA and phenoloxidase may be arrived at.
ACKNOWLEDGMENTS
We thank Malaysia Ministry of Science, Technology and Innovations for funding this project through the Institute of Pharmaceutical and Nutraceutical Malaysia with grant number Vot 63912.
REFERENCES
- Rossolini, G.M., F. Arena, P. Pecile and S. Pollini, 2014. Update on the antibiotic resistance crisis. Curr. Opin. Pharmacol., 18: 56-60.
CrossRefDirect Link - Roca, I., M. Akova, F. Baquero, J. Carlet and M. Cavaleri et al., 2015. The global threat of antimicrobial resistance: Science for intervention. New Microbes New Infect., 6: 22-29.
CrossRefDirect Link - Chaudhary, A.S., 2016. A review of global initiatives to fight antibiotic resistance and recent antibiotics׳ discovery. Acta Pharm. Sin. B, 6: 552-556.
CrossRefPubMedDirect Link - Ventola, C.L., 2015. The antibiotic resistance crisis: Part 1: Causes and threats. Pharm. Ther., 40: 277-283.
PubMedDirect Link - Nagarajan, K., S.K. Marimuthu, S. Palanisamy and L. Subbiah, 2017. Peptide therapeutics versus superbugs: Highlight on current research and advancements. Int. J. Pept. Res. Ther., 24: 19-23.
CrossRefDirect Link - Coates, C.J. and J. Nairn, 2014. Diverse immune functions of hemocyanins. Dev. Comp. Immunol., 45: 43-55.
CrossRefDirect Link - Decker, H. and F. Tuczek, 2000. Tyrosinase/catecholoxidase activity of hemocyanins: Structural basis and molecular mechanism. Trends Biochem. Sci., 25: 392-397.
CrossRefDirect Link - Solomon, E.I., U.M. Sundaram and T.E. Machonkin, 1996. Multicopper oxidases and oxygenases. Chem. Rev., 96: 2563-2606.
CrossRefPubMedDirect Link - Decker, H., M. Ryan, E. Jaenicke and N. Terwilliger, 2001. SDS-induced phenoloxidase activity of hemocyanins from Limulus polyphemus, Eurypelma californicum and Cancer magister. J. Biol. Chem., 276: 17796-17799.
CrossRefDirect Link - Nagai, T., T. Osaki and S.I. Kawabata, 2001. Functional conversion of hemocyanin to phenoloxidase by horseshoe crab antimicrobial peptides. J. Biol. Chem., 276: 27166-27170.
CrossRefDirect Link - Decker, H. and E. Jaenicke, 2004. Recent findings on phenoloxidase activity and antimicrobial activity of hemocyanins. Dev. Comp. Immunol., 28: 673-687.
CrossRefPubMedDirect Link - Decker, H., N. Hellmann, E. Jaenicke, B. Lieb, U. Meissner and J. Markl, 2007. Minireview: Recent progress in hemocyanin research. Integr. Comp. Biol., 47: 631-644.
CrossRefDirect Link - Nillius, D., E. Jaenicke and H. Decker, 2008. Switch between tyrosinase and catecholoxidase activity of scorpion hemocyanin by allosteric effectors. FEBS Lett., 582: 749-754.
CrossRefDirect Link - Lei, K., F. Li, M. Zhang, H. Yang, T. Luo and X. Xu, 2008. Difference between hemocyanin subunits from shrimp Penaeus japonicus in anti-WSSV defense. Dev. Comp. Immunol., 32: 808-813.
CrossRefDirect Link - Coates, C.J. and H. Decker, 2014. Immunological properties of oxygen-transport proteins: Hemoglobin, hemocyanin and hemerythrin. Cell. Mol. Life Sci., 74: 293-317.
CrossRefDirect Link - Wang, Y.Y., P.C.P. Cheng and D.W. Chan, 2003. A simple affinity spin tube filter method for removing high‐abundant common proteins or enriching low‐abundant biomarkers for serum proteomic analysis. Proteomics, 3: 243-248.
CrossRefDirect Link - Hengel, S.M., E. Floyd, E.S. Baker, R. Zhao, S. Wu and L. Pasa-Tolic, 2012. Evaluation of SDS depletion using an affinity spin column and IMS-MS detection. Proteomics, 12: 3138-3142.
CrossRefDirect Link - Laemmli, U.K., 1970. Cleavage of structural proteins during the assembly of the head of bacteriophage T4. Nature, 227: 680-685.
CrossRefDirect Link - Fan, T., Y. Zhang, L. Yang, X. Yang, G. Jiang, M. Yu and R. Cong, 2009. Identification and characterization of a hemocyanin-derived phenoloxidase from the crab Charybdis japonica. Comp. Biochem. Physiol. Part B: Biochem. Mol. Biol., 152: 144-149.
CrossRefDirect Link - Reller, L.B., M. Weinstein, J.H. Jorgensen and M.J. Ferraro, 2009. Antimicrobial susceptibility testing: A review of general principles and contemporary practices. Clin. Infect. Dis., 49: 1749-1755.
CrossRefDirect Link - Tokarzewski, S., G. Ziolkowska and A. Nowakiewicz, 2012. Susceptibility testing of Aspergillus niger strains isolated from poultry to antifungal drugs-a comparative study of the disk diffusion, broth microdilution (M 38-A) and Etest® methods. Polish J. Vet. Sci., 15: 125-133.
Direct Link - Coates, C.J. and J. Talbot, 2018. Hemocyanin-derived phenoloxidase reaction products display anti-infective properties. Dev. Comp. Immunol., 86: 47-51.
CrossRefDirect Link - Carrillo-Munoz, A.J., G. Quindos, C. Tur, M.T. Ruesga and Y. Miranda et al., 1999. In-vitro antifungal activity of liposomal nystatin in comparison with nystatin, amphotericin B cholesteryl sulphate, liposomal amphotericin B, amphotericin B lipid complex, amphotericin B desoxycholate, fluconazole and itraconazole. J. Antimicrob. Chemother., 44: 397-401.
CrossRefPubMedDirect Link - Schenk, S., J. Schmidt, U. Hoeger and H. Decker, 2015. Lipoprotein-induced phenoloxidase-activity in tarantula hemocyanin. Biochim. Biophys. Acta (BBA)-Proteins Proteomics, 1854: 939-949.
CrossRefDirect Link - Laino, A., S. Lavaris, G. Suarez, A. Lino and M. Cunnigham, 2015. Characterization of phenoloxidase activity from spider Polybetes pythagoricus hemocyanin. J. Exp. Zool.-A Ecol. Genet. Physiol., 323: 547-555.
CrossRefDirect Link - Yoon, B.K., J.A. Jackman, M.C. Kim and N.J. Cho, 2015. Spectrum of membrane morphological responses to antibacterial fatty acids and related surfactants. Langmuir, 31: 10223-10232.
CrossRefDirect Link - Coates, C.J., S.M. Kelly and J. Nairn, 2011. Possible role of phosphatidylserine-hemocyanin interaction in the innate immune response of Limulus polyphemus. Dev. Comp. Immunol., 35: 155-163.
CrossRefDirect Link - Gonzalez, A., E. Nova, M. del Campo, A. Manubens, A. de Ioannes, J. Ferreira and M.I. Becker, 2017. The oxygen-binding properties of hemocyanin from the mollusk Concholepas concholepas. Biochim. Biophys. Acta (BBA)-Proteins Proteomics, 1865: 1746-1757.
CrossRefDirect Link - Sterner, R. and H. Decker, 1994. Inversion of the Bohr effect upon oxygen binding to 24-meric tarantula hemocyanin. Proc. Natl. Acad. Sci. USA., 91: 4835-4839.
CrossRefDirect Link - Hirota, S., N. Tanaka, I. Micetic, P. di Muro, S. Nagao, H. Kitagishi and L. Bubacco, 2010. Structural basis of the lactate-dependent allosteric regulation of oxygen binding in arthropod hemocyanin. J. Biol. Chem., 285: 19338-19345.
CrossRefDirect Link - Brouwer, M., C. Bonaventura and J. Bonaventura, 1983. Metal ion interactions with Limulus polyphemus and Callinectes sapidus hemocyanin: Stoichiometry and structural and functional consequences of calcium(II), cadmium(II), zinc(II) and mercury(II) binding. Biochemistry, 22: 4713-4723.
CrossRefDirect Link - Castellanos-Serra, L. and D. Paz-Lago, 2002. Inhibition of unwanted proteolysis during sample preparation: Evaluation of its efficiency in challenge experiments. Electrophoresis, 23: 1745-1753.
CrossRefDirect Link - Besser, K., G.P. Malyon, W.S. Eborall, G.P. da Cunha and J.G. Filgueiras et al., 2018. Hemocyanin facilitates lignocellulose digestion by wood-boring marine crustaceans. Nat. Commun., Vol. 9.
CrossRefDirect Link - Dolashka, P., A. Dolashki, J.V. Beeumen, M. Floetenmeyerd, L. Velkova, S. Stevanovic and W. Voelter, 2016. Antimicrobial activity of molluscan hemocyanins from Helix and Rapana snails. Curr. Pharm. Biotechnol., 17: 263-270.
Direct Link