
Mudasar Nabi
Department of Biochemistry, University of Kashmir, Srinagar 190006, India
Aaliya Bhat
Department of Biochemistry, University of Kashmir, Srinagar 190006, India
Shayaq ul Abeer Rasool
Department of Biotechnology, University of Kashmir, Srinagar 190006, India
Sairish Ashraf
Department of Biochemistry, University of Kashmir, Srinagar 190006, India
Rouf Maqbool
Department of Biochemistry, University of Kashmir, Srinagar 190006, India
Showkat Ahmad Ganie
Department of Biochemistry, University of Kashmir, Srinagar 190006, India
Shajrul Amin
Department of Biochemistry, University of Kashmir, Srinagar 190006, India
LiveDNA: 91.24997
Pakistan Journal of Biological Sciences
Year: 2018 | Volume: 21 | Issue: 9 | Page No.: 432-440







ABSTRACT
Background and Objective: Protease inhibitors (PIs) regulate various cellular processes like cell cycle, differentiation, apoptosis and immune responses. Leguminous seeds are rich sources of protease inhibitors and many novel protease inhibitors have been purified from them. To isolate and purify protease inhibitors from seeds of Sophora japonica, characterize and investigate their anti- microbial activity. Materials and Methods: Protease inhibitors (SJ-pi I and SJ-pi II) were purified to homogeneity by ammonium sulfate precipitation, Ion exchange chromatography and column chromatography. The molecular mass was estimated by size exclusion chromatography and by SDS-PAGE and anti- microbial activity was tested by agar disk diffusion method. Results: Two protease inhibitors were isolated and purified from Sophora japonica seeds, SJ-pi I and SJ-pi II, with molecular weight of 15.1 and 31 kDa, respectively. Both purified inhibitors were active over a range of pH (6.0-9.0) and showed maximum activity in the temperature range of 30-40°C. They inhibited the growth of three Gram-positive bacteria. Conclusion: Protease inhibitors were classified as serine protease inhibitors, however further necessary structural investigations need to be carried out so as to group them into specific class of serine protease inhibitors.
PDF Abstract XML References Citation
Received: September 19, 2018;
Accepted: November 11, 2018;
Published: November 29, 2018
Copyright: © 2018. This is an open access article distributed under the terms of the creative commons attribution License, which permits unrestricted use, distribution and reproduction in any medium, provided the original author and source are credited.
How to cite this article
Mudasar Nabi, Aaliya Bhat, Shayaq ul Abeer Rasool, Sairish Ashraf, Rouf Maqbool, Showkat Ahmad Ganie and Shajrul Amin, 2018. Physio-chemical Characterization and Anti-microbial Activity of Serine Protease Inhibitors Purified from the Sophora japonica Seeds. Pakistan Journal of Biological Sciences, 21: 432-440.
DOI: 10.3923/pjbs.2018.432.440
URL: https://scialert.net/abstract/?doi=pjbs.2018.432.440
DOI: 10.3923/pjbs.2018.432.440
URL: https://scialert.net/abstract/?doi=pjbs.2018.432.440
INTRODUCTION
Proteases, despite the fact that they are indispensable to the cells and organisms that host them, may be potentially damaging when over expressed or present in higher concentrations. Their role in the physiology of many human diseases, ranging from cancer and inflammatory disorders to degenerative diseases, now represents an increasingly important feature of these enzymes, besides being involved in carcinogenesis and promoting proliferation, invasion and metastasis1,2. For this reason the activities of these enzymes need to be strictly regulated and controlled which is carried out by number of different mechanisms including regulation of gene expression, recognition of the substrate by the active site, activity regulation by small molecules, changes in cellular location, post-translational modifications, interaction with other proteins and/or through inhibition of proteolysis by protease inhibitors3-5.
Protease inhibitors are of ubiquitous nature and have been isolated from plants, animals and micro-organisms6-8 . In plant kingdom protease inhibitors are widely found in seeds of Graminaceae and Leguminosae, as well as in tubers of Solanaceae. In legumes, PIs accumulate in large amounts during seed maturation and play an important role both in the deposition of storage protein and in plant defense9-12. Their physiological role includes regulation of endogenous proteinases during seed dormancy, reserve protein mobilization and protection against proteolytic enzymes of parasites and insects10. Besides the pivotal function of these PIs in plants, an understanding of the role played by protease and their specific inhibitors in human diseases offers novel and challenging opportunities for preventive and/or therapeutic intervention13. Naturally-occurring plant protease inhibitors are being investigated for their role in the prevention and/or treatment of a diverse set of human pathologies, including cancer, neuro-degenerative and cardiovascular diseases, muscle atrophy and inflammatory disorders. In this context, the United States Food and Drug Administration (FDA) granted a protein extract of soybean (Glycine max) enriched in Bowman-Birk inhibitors (BBI), namely Bowman-Birk inhibitor concentrate (BBIC), as investigational new drug. Up to six clinical trials have been accomplished in patients with benign prostatic hyperplasia, oral leukoplakia and ulcerative colitis14. Although, in most of these cases the intrinsic ability of BBI to inhibit serine proteases has been related to beneficial health properties, the mechanism of action and the identity of their therapeutic targets are largely unknown.
Within this framework, there is a growing interest in naturally-occurring protease inhibitors due to their potential chemo preventive and/or therapeutic properties which can impact on several human diseases, including cancer, neuro-degenerative diseases and inflammatory disorders.
Therefore, the present investigation was planned to screen the Sophora japonica seeds for the presence of protease inhibitor/s and the physio-chemical characterization of the isolated Protease inhibitor (PI). Further, isolated PI/s were analyzed for their anti-tryptic, anti-chymotryptic and anti-microbial activity.
MATERIALS AND METHODS
The work was carried out in Protein Biochemistry Lab, Department of Biochemistry, University of Kashmir, Jammu and Kashmir, India over the period of 2 years from December 2014 to January 2017.
Isolation and purification of protease inhibitors from Sophora Japonica seeds: Mature seeds of locally available legume plants Sophora japonica were selected and procured from the Department of Botany, University of Kashmir. Seeds were shade dried before processing. All extraction processes were carried out at 4°C. Dried seeds (25 g) of the plant were soaked in NaCl (15N) for 24 h. They were then homogenized in ice cold saline-Tris buffer (20 mM Tris, pH 8.0) containing 1 mM sodium metabisulphite at 4°C for 10 min. The filtered homogenate was subjected to centrifugation at 12,00 g for 20 min, the pellet discarded and supernatant retained and stored at 4°C. The proteins were precipitated from the supernatant by 90% saturation of ammonium sulphate and again centrifuged at 12,000 g for 10 min. The pellet formed was dissolved in the minimum volume of double distilled water and dialyzed, against 20 mM Tris buffer, pH 8. Further purification was done on Diethylaminoethyl Cellulose (DEAE-Cellulose) column based ion-exchange chromatography, using Tris-buffer (20 mM, pH 8). The protein was eluted using the linear sodium chloride gradient from 0.05-0.6 M. The eluted fractions were then chromatographed separately on Sephadex G-100 column, using Tris-buffer (0.1 M, pH 8) and NaCl (0.4 M). The eluted fractions were checked for protein concentration by measuring absorbance at 280 nm.
The protein fractions on Sephadex G-100 column resolved into two peak fractions which were named as SJ-pi I (Sophora japonica protease inhibitor I) and SJ-pi II (Sophora japonica protease inhibitor II), respectively.
SDS polyacrylamide gel electrophoresis: Fractions eluted from Sephadex Gel chromatography were checked by SDS PAGE. Molecular weight of purified inhibitors was also determined by SDS-PAGE. For this a protein ladder of known molecular weight was electrophorised along with purified protease inhibitors.
Measurement of trypsin inhibitory activity: The biological activity of inhibitors was measured by the degree of inhibition of tryptic activity using BAPNA as substrate. Different volumes of purified inhibitors were incubated with 10 mg of trypsin (diluted in 1 mM HCl) and with variable volumes of the 0.1 M Tris-HCl buffer, pH 8.0 containing 0.01 M CaCl2. The final volume was made up to 500 μL. The contents were incubated for 10 min at 37°C and 500 μL of N-Benzoyl-DL-arginine-4-nitroanilide hydrochloride (BAPNA) (8 mM in DMSO) was added. The reaction mixture was incubated for 40 min at 47°C and reaction stopped by the addition of 500 μL of acetic acid (30% v/v). The control, lacking inhibitor was prepared under exactly identical conditions. Blank for each volume of the inhibitor tested was obtained by the addition of acetic acid just before the incubation with the substrate BAPNA15. The intensity of the yellow color produced was measured by recording the absorbance at 405 nm. Residual trypsin activity was calculated using the following equation:
Residual trypsin activity (%) = Abs at 405 nm in contro l- Abs at 405 nm in sample×100
Inhibition (%) = 100-Residual trypsin activity (%)
Measurement of chymotrypsin inhibitory activity: Inhibition of chymotrypsin activity was determined on acetyl L-Tyrosine ester (ATEE). Again, different volumes of each inhibitor was pre incubated with 10 μg of enzyme solution of 5 min at room temperature in 1 mL of 46 mM Tris 0.5 mM CaCl2 buffer, pH 8.1 as per the protocol given by Birk et al.16. The reaction was started by the addition of 3 mL of substrate solution to pre incubated enzyme plus inhibitor solution. Blank was prepared for each volume of extract tested by omitting the enzyme from the reaction mixture. The intensity of yellow color produced was measured by recording the absorbance at 410 nm against an appropriate blank. Control, where chymotrypsin was not pre-incubated with inhibitor was also prepared. Percent inhibition of chymotrypsin activity was calculated by using following relationship:
Effect of pH and temperature on inhibition activity of Protease inhibitors: To determine the pH optima, 50 μL purified protease inhibitors were incubated at 30°C in various pH adjusted buffers e.g., 0.1 M Glycine HCl puffer, pH 2.0 and 3.0.0.1 M NaOAc buffer pH 4.2, 4.6, 5.0 and 5.2, 0.1 M sodium carbonate buffer, pH 10. The inhibitory activities were determined separately in each buffer system.
For determining the temperature optima, 20 μL purified PIs were incubated at various temperatures from 10-80°C, for 10 min and then anti-tryptic activities were determined.
Assaying anti-microbial activity of purified protease inhibitors: The antimicrobial activity of purified protease inhibitors was determined against bacteria (Klebsiella pneumonia, Escherichia coli, Staphylococcus aureus, Bacillus subtilis and Streptococcus pneumonia) and against fungi (Candida albicans and Saccharomyces cerevisiae) by agar well diffusion method17. Gentamycin was used as standard anti bacterial drug and Nystatin was used as standard anti-fungal drug to compare zone of inhibition. The diameter of inhibition zone was measured in millimeters.
RESULTS
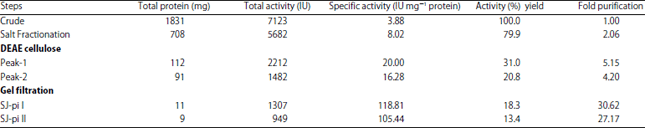
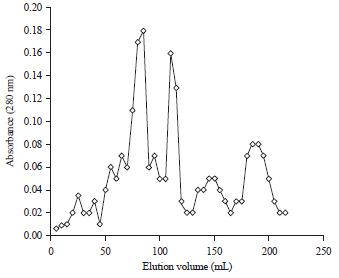
Isolation and purification of protease inhibitors from sophora japonica seeds: Protease inhibitors were purified to homogeneity from seeds of Sophora japonica The crude protein fraction resolved two major peaks and some minor peaks on DEAE cellulose chromatography (Fig. 1). Protein fractions corresponding to the two major peaks showed protease inhibitor activity where as protein fractions corresponding to the minor peaks didn’t show any inhibitory activity against proteases. The pooled major peaks resolved into one major peak subjected to gel filtration chromatography on Sephadex G-100 column (Fig. 2). The fractions under major peaks were found to have anti-protease activity and were named as SJ-pi I (Sophora japonica protease inhibitor I) and SJ-pi II (Sophora japonica protease inhibitor II), respectively. At each purification step specific activity of inhibitors increased over homogenate the results of which were given in Table 1.
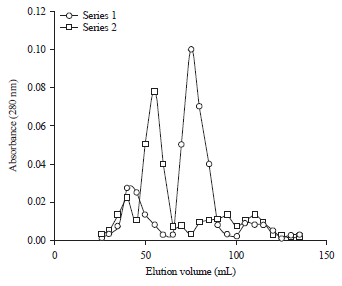
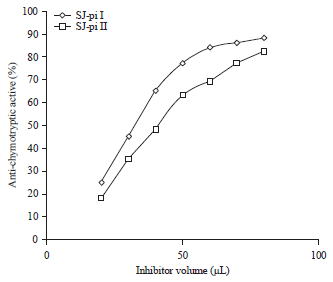
Anti-tryptic and anti-chymotryptic activity: The SJ-pi I and SJ-pi II inhibited activity of Trypsin by 91 and 83%, respectively at the molar ratio of 1:1 (Fig. 3). SJ-pi I and SJ-pi II also inhibited activity of chymotrypsin by 89 and 81%, respectively at the molar ratio of 1:1 (Fig. 4).
| Table 1: | Purification of protease inhibitors from Sophora japonica seeds |
 | |
| BAPNA was used as substrate to measure activity of Trypsin inhibitor. One inhibitor unit (IU) is defined as amount of inhibitor which suppressed liberation of 1μmol substrate per min by active enzyme | |
 | |
| Fig. 1: | Elution profile of crude extract of Sophora japonica on DEAE-cellulose column |
 | |
| Fig. 2: | Elution profiles of crude peak 1 and peak 2 obtained from DEAE cellulose column on Sephadex G-100 column (Series 1 represents absorbance at 280 nm of Peak 1 obtained from DEAE cellulose column and series 2 represents absorbance of Peak 2) |
 | |
| Fig. 3: | Inhibition profile of trypsin activity by purified inhibitors from seeds of Sophora japonica (measured by % inhibition of tryptic activity using BAPNA as substrate) |
 | |
| Fig. 4: | Inhibition profile of chymotrypsin activity by purified inhibitors from seeds of Sophora japonica (measured by inhibition (%) using ATEE as substrate) |
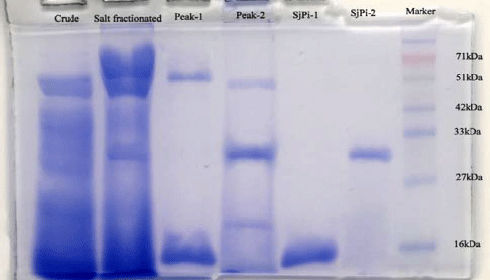 | |
| Fig. 5: | SDS-polyacrylamide gel electrophoretic pattern of crude homogenate and purified inhibitors. Electrophoresis was carried at a current of 50 mA on 4% stacking and 10% resolving gel. About 50 μg of protein was applied in the wells |
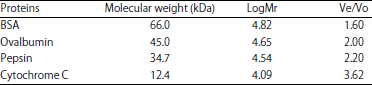
| Table 2: | Molecular weight and Ve/Vo of standard proteins used in calibration of Sephadex G-100 column |
 | |
| Table 3: | Ve/Vo and molecular weight of protease inhibitors SJ-pi I and SJ-pi II |
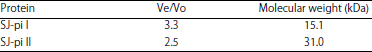 | |
Determination of molecular weight: The molecular weight of the purified inhibitors was determined by gel filtration chromatography on Sephadex G-100 column, pre-equilibrated with standard proteins as shown in Table 2. The molecular weight of purified inhibitors SJ-pi I and SJ-pi II was found to be 15.1 and 31 kDa, respectively by gel filtration chromatography (Table 3), which was further confirmed by SDS-PAGE (Fig. 5).
Effect of pH and temperature on activity of protease inhibitor: Both the inhibitors were active over a range of pH (6.0-9.0). However the maximum activity of SJ-pi I was found to be at pH 8.0 and SJ-pi II had maximum activity at pH 7.0, Fig. 6. Thermo stability of both inhibitors was determined at determined at temperatures ranging from 10-70°C. The purified inhibitors had the maximum activity in the temperature range of 30-40°C. The SJ-pi I was active even at 50°C.
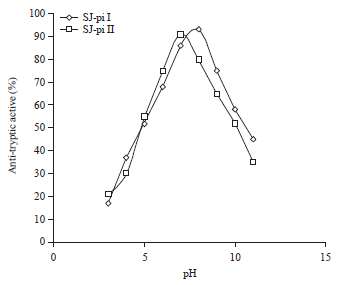 | |
| Fig. 6: | Effect of pH on activity of protease inhibitors purified from seeds of Sophora japonica |
The activity of both the inhibitors decreased markedly with further increase in temperature (Fig. 7).
Assaying anti-microbial activity of extracts: Purified inhibitors didn’t show any inhibitory activity against gram negative Klebsiella pneumoniae, Escherichia coli and against fungal strains Candida albicans and Saccharomyces cerevisiae (Fig. 8). However, both inhibitors exhibited anti-bacterial activity against Staphylococcus aureus, Bacillus subtilis and Streptococcus pneumonia (Fig. 9). The inhibition results are given in Table 4.
DISCUSSION
Protease inhibitors (PIs) are ubiquitous small proteins that interfere and limit the proteolytic activity of proteases.
| Table 4: | Measurement of zone of inhibition of various bacterial strains |
 | |
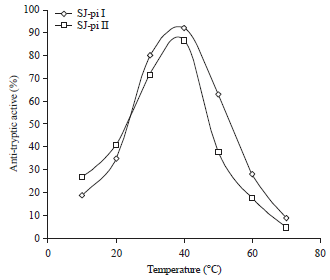 | |
| Fig. 7: | Effect of temperature on activity of protease inhibitors purified from seeds of Sophora japonica |
They have been purified and characterized from a number of sources including plants animals and micro-organisms. Most of the plant protease inhibitors are proteinaceous in nature and are located mainly in seeds, leaves and tubers, which act as specific defense and regulatory proteins. Naturally occurring protease inhibitors are essential for regulating the activity of their corresponding proteases and play key role in biological processes. PIs act as defense agents against insects and micro-organisms18-20. Currently PIs are being investigated for their role in the prevention and/or treatment of a diverse set of human pathologies, including cancer, neuro-degenerative and cardiovascular diseases and inflammatory disorders21. Many reports are available on the isolation, purification and characterization of protease inhibitor from seeds of legume plants19,22-25. Two protease inhibitors were purified to homogeneity from seeds of Sophora japonica in three steps by ammonium sulfate precipitation, DEAE cellulose column chromatography and Sephadex G-100 chromatography. The crude homogenate when subjected to ion exchange chromatography resolved into three major peaks and some minor peaks, however the anti-protease activity was only found in two major peaks. Similar results were reported by Rakashanda et al.7 for lavatera cashmeriana seeds. The two major peaks with anti-protease activity obtained from DEAE cellulose column were pooled and further subjected to purification on gel filtration chromatography using Sephadex G-100 column. Both the peaks individually resolved into a single major peak each with antiprotease activity, suggesting the presence of two protease inhibitors.
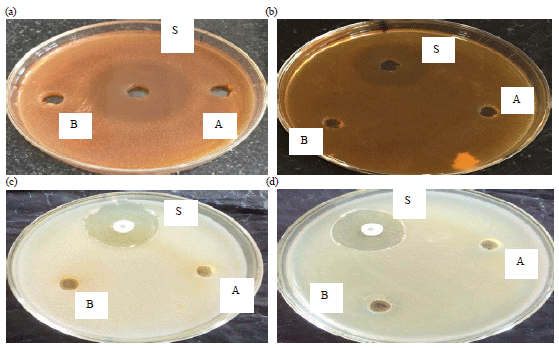 | |
| Fig. 8(a-d): | Anti-microbial activity of purified inhibitors against Gram-negative bacterial and fungal strains. A represents SJ-pi I, B represents SJ-pi II and S represents standard drug, (a) Saccharomyces cerevisiae, (b) Candida albicans, (c) Escherichia coli and (d) Klebsiella pneumonia |
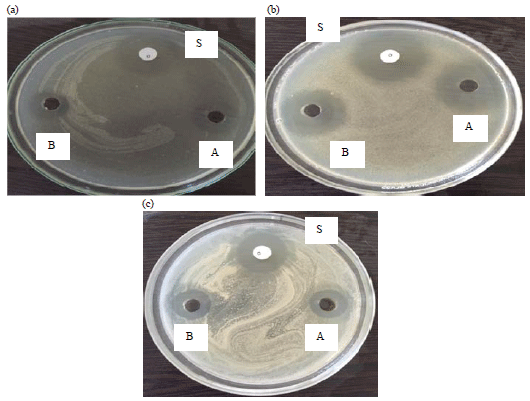 | |
| Fig. 9(a-d): | Anti-microbial activity of purified inhibitors against Gram-positive bacteria, A: SJ-pi I, bB: SJ-pi II and S: Standard drug (Gentamycin), (a) Streptococcus pneumonia, (b) Bacillus subtilis and (c) Streptococcus aureus |
The peak fractions were pooled and named as SJ-pi I and SJ-pi II (Sophora japonica protease inhibitor). The results were in agreement with the study carried out by Sharma et al.26 for Albizia lebbeck seeds. The molecular weight of the two purified inhibitors was determined by SDS PAGE and by gel filtration column chromatography. The molecular weight of SJ-pi I was found to be 15.1 kDa and the molecular weight of SJ-pi II was found to be 31 kDa. Both the purified inhibitors were found to consist of a single polypeptide chain as revealed by SDS-PAGE which is consistent with inhibitors purified from Tamarindus indica seeds27 and with the protease purified inhibitors from Prosopisjuliflora seeds28. On the contrary trypsin inhibitor purified from A. kalkora seeds showed the presence of two disulfide linked polypeptide chains29.
The purified protease inhibitors strongly inhibited activity of trypsin and chymotrypsin. The SJ-pi I inhibited 91% trypsin activity and 83% chymotrypsin activity and SJ-pi II inhibited 89% trypsin activity and 81% chymotrypsin activity at the molar ratio of 1:1. This is comparable to the inhibitors purified from lavatera cashmeriana seeds7 . Both the inhibitors were active over a range of pH (6.0-9.0). However the maximum activity of SJ-pi I was found to be at pH 8.0 and SJ-pi II had maximum activity at pH 7.0 which is consistent with earlier reports30. The trypsin inhibitor purified from mungbean seeds was found to be active between the pH ranges of 4.0-10.0. In contrast, the trypsin inhibitor purified from pigeon pea seeds had maximum activity between pH 7.0 and 10.0, but when exposed to acidic conditions pH 3.0-5.0 the activity was lost31 by 20%. Thermostability of purified inhibitors was determined at different temperatures ranging from 10-70°C. The purified inhibitors had maximum activity in the temperature range of 30-40°C. However, unlike SJ-pi II, SJ-pi I was stable even at 50°C. The activity of both the inhibitors decreased markedly with further increase in temperature. Trypsin inhibitors purified from seeds of Caesalpinia bonduc and Archidendron ellipticum were found to be heat tolerant32 up to 60°C likewise the protease inhibitors purified from Lavatera cashmeriana seeds had the optimum temperature of 30°C however, they were active even upto 60°C. Both SJ-pi I and SJ-pi II had significant antibacterial activity against three tested Gram-positive bacterial strains. However, inhibitors did not exhibit anti-bacterial activity against tested Gram-negative bacterial strains and also they didn’t show any antifungal activity against the tested fungal strains. Earlier reports have also shown proteinase inhibitor purified from Ginkgo biloba seeds did not exhibit any antifungal activity neither it had antibacterial activity against Escherichia coli33.
CONCLUSION
Two protease inhibitors SJ-pi I and SJ-pi II were purified from seeds of Sophora japonica with molecular weight of 15.1 kDa and 31 kDa respectively. Both inhibitors were active over a range of temperature and pH and were found to inhibit the two most important enzymes of serine proteases family viz trypsin and chymotrypsin, hence these two inhibitors can be grouped into the serine protease inhibitors. However further necessary structural investigations needs to be carried out so as to group them into specific class of serine protease inhibitors. SJ-pi I and SJ-pi II possess significant anti bacterial activity against Gram-positive bacterial strains. These PIs can be ideal candidate for exploration in various therapeutic and biological applications.
SIGNIFICANCE STATEMENT
This study discovers two protease inhibitors, SJ-pi I and SJ-pi II, active at optimum pH and temperature and show promising antibacterial activity. This study will be beneficial in pharmaceutical, agricultural and in industrial field. This study provides a platform to the researchers for future studies of these two protease inhibitors to further characterize them and to elucidate their role in various biological pathways.
REFERENCES
- Yang, Y., H. Hong, Y. Zhang and W. Cai, 2009. Molecular imaging of proteases in cancer. Cancer Growth Metastasis, Vol. 2.
CrossRefDirect Link - Hsieh, C.C., B. Hernandez-Ledesma, H.J. Jeong, J.H. Park and O. Ben, 2010. Complementary roles in cancer prevention: Protease inhibitor makes the cancer preventive peptide lunasin bioavailable. Plos One, Vol. 5.
CrossRefDirect Link - Turk, B., 2006. Targeting proteases: Successes, failures and future prospects. Nat. Rev. Drug Discov., 5: 785-799.
CrossRefDirect Link - Drag, M. and G.S. Salvesen, 2010. Emerging principles in protease-based drug discovery. Nature Rev. Drug Discovery, 9: 690-701.
CrossRefDirect Link - Shen, A., 2010. Allosteric regulation of protease activity by small molecules. Mol. Biosyst., 6: 1431-1443.
CrossRefDirect Link - Hossain, M.S., A.K. Azad, S.M. Abu Sayem, G. Mostafa and M.M. Hoq, 2007. Production and partial characterization of feather-degrading keratinolytic serine protease from Bacillus licheniformis MZK-3. J. Boil. Sci., 7: 599-606.
CrossRefDirect Link - Rakashanda, S., S. Mubashir, Y. Qurishi, A. Hamid, A. Masood and S. Amin, 2013. Trypsin inhibitors from Lavatera cashmeriana Camb. seeds: Isolation, characterization and in-vitro cytoxicity activity. Int. J. Pharm. Sci. Invent., 2: 55-65.
Direct Link - Gandreddi, V.S., V.R. Kappala, K. Zaveri and K. Patnala, 2015. Evaluating the role of a trypsin inhibitor from soap nut (Sapindus trifoliatus L. Var. Emarginatus) seeds against larval gut proteases, its purification and characterization. BMC Biochem., Vol. 16.
CrossRefDirect Link - Koiwa, H., R.A. Bressan and P.M. Hasegawa, 1997. Regulation of protease inhibitors and plant defense. Trends Plant Sci., 2: 379-384.
CrossRefDirect Link - Cruz, A.C., F.S. Massena, L. Migliolo, L.L. Macedo and N.K. Monteiro et al., 2013. Bioinsecticidal activity of a novel Kunitz trypsin inhibitor from Catanduva (Piptadenia moniliformis) seeds. Plant Physiol. Biochem., 70: 61-68.
CrossRefDirect Link - Smid, I., A. Rotter, K. Gruden, J. Brzin and M.B. Gasparic et al., 2015. Clitocypin, a fungal cysteine protease inhibitor, exerts its insecticidal effect on Colorado potato beetle larvae by inhibiting their digestive cysteine proteases. Pesticide Biochem. Physiol., 122: 59-66.
CrossRefDirect Link - Sels, J., J. Mathys, B.M.A. de Coninck, B.P.A. Cammue and M.F.C. de Bolle, 2008. Plant Pathogenesis-Related (PR) proteins: A focus on PR peptides. Plant Physiol. Biochem., 46: 941-950.
CrossRefDirect Link - Deu, E., M. Verdoes and M. Bogyo, 2012. New approaches for dissecting protease functions to improve probe development and drug discovery. Nature Struct. Mol. Biol., 19: 9-16.
Direct Link - Clemente, A., F.J. Moreno, M.D.C. Marin‐Manzano, E. Jimenez and C. Domoney, 2010. The cytotoxic effect of Bowman-Birk isoinhibitors, IBB1 and IBBD2, from soybean (Glycine max) on HT29 human colorectal cancer cells is related to their intrinsic ability to inhibit serine proteases. Mol. Nutr. Food Res., 54: 396-405.
CrossRefDirect Link - Lin, J.Y., S.C. Chu, H.C. Wu and Y.S. Hsieh, 1991. Trypsin inhibitor from the seeds of Acacia confusa. J. Biochem., 110: 879-883.
PubMedDirect Link - Birk, Y., A. Gertler and S. Khalef, 1963. A pure trypsin inhibitor from soya beans. J. Biochem., 87: 281-284.
PubMedDirect Link - Bonev, B., J. Hooper and J. Parisot, 2008. Principles of assessing bacterial susceptibility to antibiotics using the agar diffusion method. J. Antimicrob. Chemother., 61: 1295-1301.
CrossRefDirect Link - Guimaraes, L.C., C.F.R. de Oliveira, S. Marangoni, D.G.L. de Oliveira and M.L.R. Macedo, 2015. Purification and characterization of a Kunitz inhibitor from Poincianella pyramidalis with insecticide activity against the Mediterranean flour moth. Pesticide Biochem. Physiol., 118: 1-9.
CrossRefDirect Link - Dantzger, M., I.M. Vasconcelos, V. Scorsato, R. Aparicio, S. Marangoni and M.L.R. Macedo, 2015. Bowman-Birk proteinase inhibitor from Clitoria fairchildiana seeds: Isolation, biochemical properties and insecticidal potential. Phytochemistry, 118: 224-235.
CrossRefDirect Link - Habib, H., K. Fazili, M. Zargar and B. Ganie, 2016. Protease inhibitor associated antimicrobial activity of pea Pisum sativum L. cv. Arkel. Int. J. Pure Applied Biosci., 4: 172-179.
Direct Link - Clemente, A., M. del Carmen Marin-Manzano, M. del Carmen Arques and C. Domoney, 2013. Bowman-Birk Inhibitors from Legumes: Utilisation in Disease Prevention and Therapy. In: Bioactive food Peptides in Health and Disease, Abdou, A.M., A. Clemente, A. Pihlanto, C. Muro and K.C. Hsu et al. (Eds.)., InTech, Croatia.
- Migliolo, L., A.S. de Oliveira, E.A. Santos, O.L. Franco and P. Mauricio, 2010. Structural and mechanistic insights into a novel non-competitive Kunitz trypsin inhibitor from Adenanthera pavonina L. seeds with double activity toward serine-and cysteine-proteinases. J. Mol. Graphics Model., 29: 148-156.
CrossRefDirect Link - De Paula, C.A.A., V.J. Coulson-Thomas, J.G. Ferreira, P.K. Maza and E. Suzuki et al., 2012. Enterolobium contortisiliquum trypsin inhibitor (EcTI), a plant proteinase inhibitor, decreases in vitro cell adhesion and invasion by inhibition of Src protein-Focal Adhesion Kinase (FAK) signaling pathways. J. Biol. Chem., 287: 170-182.
Direct Link - Clemente, A., J.M. Gee, I.T. Johnson, D.A. MacKenzie and C. Domoney, 2005. Pea (Pisum sativum L.) protease inhibitors from the bowman-Birk class influence the growth of human colorectal adenocarcinoma HT29 cells in vitro. J. Agric. Food Chem., 53: 8979-8986.
CrossRefDirect Link - Chevreuil, L.R., J.F.D.C. Goncalves, L.D.A. Calderon, L.A.G.D. Souza and S.C. Pando, 2014. Partial purification of trypsin inhibitors from Parkia seeds (Fabaceae). Hoehnea, 41: 181-186.
CrossRefDirect Link - Sharma, P., A.K. Nath, R. Kumari, S.V. Bhardwaj, 2012. Purification, characterization and evaluation of insecticidal activity of trypsin inhibitor from Albizia lebbeck seeds. J. For. Res., 23: 131-137.
CrossRefDirect Link - Fook, J.M.S.L.L., L.L.P. Macedo, G.E.D.D. Moura, F.M. Teixeira, A.S. Oliveira, A.F.S. Queiroz and M.P. Sales, 2005. A serine proteinase inhibitor isolated from Tamarindus indica seeds and its effects on the release of human neutrophil elastase. Life Sci., 76: 2881-2891.
CrossRefDirect Link - Sivakumar, S., O.L. Franco, P.D. Tagliari, C. Bloch, Jr., M. Mohan and B. Thayumanavan, 2005. Screening and purification of a novel trypsin inhibitor from Prosopis juliflora seeds with activity toward pest digestive enzymes. Protein Peptide Lett., 12: 561-565.
CrossRefDirect Link - Zhou, J.Y., H. Liao, N.H. Zhang, L. Tang, Y. Xu and F. Chen, 2008. Identification of a Kunitz inhibitor from Albizzia kalkora and its inhibitory effect against pest midgut proteases. Biotechnol. Lett., 30: 1495-1499.
CrossRefDirect Link - Kansal, R., R.N. Gupta, K.R. Koundal, K. Kuhar and V.K. Gupta, 2008. Purification, characterization and evaluation of insecticidal potential of trypsin inhibitor from mungbean (Vigna radiate L. Wilczek) seeds. Acta Physiol. Plant., Vol. 30.
CrossRefDirect Link - Godbole, S.A., T.G. Krishna and C.R. Bhatia, 1994. Purification and characterisation of protease inhibitors from pigeon pea (Cajanus cajan (l) millsp) seeds. J. Sci. Food Agric., 64: 87-93.
CrossRefDirect Link - Bhattacharyya, A., S. Rai and C.R. Babu, 2007. A trypsin and chymotrypsin inhibitor from Caesalpinia bonduc seeds: isolation, partial characterization and insecticidal properties. Plant Physiol. Biochem., 45: 169-177.
CrossRefDirect Link - Sawano, Y., K.I. Hatano, T. Miyakawa, H. Komagata, Y. Miyauchi, H. Yamazaki and M. Tanokura, 2008. Proteinase inhibitor from ginkgo seeds is a member of the plant nonspecific lipid transfer protein gene family. Plant Physiol., 146: 1909-1919.
CrossRefDirect Link