
Muddasir Basheer
Department of Zoology, Guru Ghasidas Vishwavidyalaya (Central University), Koni, Bilaspur, 495009 Chhattisgarh, India
LiveDNA: 91.16824
Seema Rai
Department of Zoology, Guru Ghasidas Vishwavidyalaya (Central University), Koni, Bilaspur, 495009 Chhattisgarh, India
LiveDNA: 91.16626
Hindole Ghosh
Department of Zoology, Guru Ghasidas Vishwavidyalaya (Central University), Koni, Bilaspur, 495009 Chhattisgarh, India
Younis Ahmad Hajam
Department of Zoology, Guru Ghasidas Vishwavidyalaya (Central University), Koni, Bilaspur, 495009 Chhattisgarh, India
LiveDNA: 91.25047
Pakistan Journal of Biological Sciences
Year: 2018 | Volume: 21 | Issue: 7 | Page No.: 340-347







ABSTRACT
Background and Objectives: Polycystic ovary syndrome (PCOS) is a common heterogeneous endocrinological and metabolic disorder in women of reproductive age which leads to infertility/subfertility. The present study was commenced to elucidate the therapeutic efficacy of melatonin in the pathogenesis of letrozole induced polycystic ovary syndrome (PCOS) in Wistar rat. Materials and Methods: Letrozole was administered orally (1 mg kg–1) to induce PCOS condition in Wistar female rats for a period of 2-3 weeks followed by a dose of melatonin (200 µg/100 g b.wt.) to PCOS induced rats. On the completion of experimental period the level of cytokines and expression level of different receptors was assessed. Results: The PCOs rats showed down regulation in melatonin (MT1 and MT2), estrogen (ER-α) and cytokine (IL-2R and IL-6R) receptors expression and high circulatory level of IL-6 and TNF-α during PCO condition. These anomalies in expression pattern and circulatory level of cytokines were restored following the treatment. Conclusion: Finding of present study showed the role of melatonin supplementation at receptor level and also suggesting a crosstalk between MT1R / MT2R via cytokine IL-2R and IL-6R resulting in modulation of ER-α receptors.
PDF Abstract XML References Citation
Received: June 15, 2018;
Accepted: September 12, 2018;
Published: November 10, 2018
Copyright: © 2018. This is an open access article distributed under the terms of the creative commons attribution License, which permits unrestricted use, distribution and reproduction in any medium, provided the original author and source are credited.
How to cite this article
Muddasir Basheer, Seema Rai, Hindole Ghosh and Younis Ahmad Hajam, 2018. Therapeutic Efficacy of Melatonin Against Polycystic Ovary Syndrome (PCOS) Induced by Letrozole in Wistar Rat. Pakistan Journal of Biological Sciences, 21: 340-347.
DOI: 10.3923/pjbs.2018.340.347
URL: https://scialert.net/abstract/?doi=pjbs.2018.340.347
DOI: 10.3923/pjbs.2018.340.347
URL: https://scialert.net/abstract/?doi=pjbs.2018.340.347
INTRODUCTION
Polycystic ovary syndrome (PCOS) is one of the most common reproductive and endocrine disorders affecting ovarian follicular development and consequently ovulation which occurs due to imbalance in steroidal hormones1,2. The diversity in its phenotypes and involvement of multiple organs contributes to complexity and heterogeneity of this syndrome of unknown etiology affecting 5-10% of women of reproductive age3. The main features of PCOS are chronic persistent anovulation, hyperandrogenism, bilateral cystic enlargement of ovaries4.
A direct action of neurohormone melatonin has already been reported on ovarian functions, steroidogenesis modulation in rodents and women5,6. Further, there is evidence of presence of high concentration of melatonin in preovulatory follicular fluid7 and this high concentration of melatonin was noted to be associated with MT1 and MT2 receptor on ovarian tissue of rat and human8.
Reproductive failure during PCO condition indicates the inflammatory ovarian response and hence be immunologically determined. Earlier findings published provides a clear cut proof regarding melatonin’s potent free radical scavenger with strong antioxidative action as well as immune enhancer in mammals including human9-12 and hence, actively involved in immune regulation/immunoproliferation13,14. Melatonin receptors also play a very important role in the physiology of an animal including inflammatory responses. Inflammatory events are particularly initiated and/or orchestrated by the sequential release of different anti/proinflammatory circulatory cytokine (s)/cytokine like molecules from local resident and infiltrating cells. Cytokines are critical to the success of reproductive process. They may act on germ cells, the embryo, non-hematopoietic cells in the gonads and uterus and hence can directly act on reproductive processes/functions. Further they also act indirectly by promoting immunologically receptive environment for the production of gametes as well as implantation and further gestational physiology till parturition. Therefore, it becomes important to understand the involvement of cytokines in the reproductive function which may help the researcher to design some therapeutic molecule aiming to reverse the reproductive pathologies like PCOS which may be a consequence of pathologies of ovarian immune origin. Therefore, the aim of present study was to examine the effect of melatonin in pathogenesis of PCOS induced by letrozole.
MATERIALS AND METHODS
Animals and maintenance: Female albino rats of Wistar strain (170±10 g b.wt.) of same age were used in this study. Animals were housed under standard husbandry conditions (25±2°C temp, 60-70% relative humidity and 12 h photoperiod) and had access to standard rat feed and drinking water ad libitum. All the animals were treated and cared in accordance with the guidelines recommended by Committee for the Purpose of Control and Supervision of Experiments on Animals (CPCSEA), Ministry of Environment and Forests, Government of India. The study was approved by Institutional Animal Ethics Committee of Guru Ghasidas Vishwavidyalaya, Bilaspur (994/GO/ERe/S/06/CPCSEA).
Preparation of letrozole and melatonin doses: Letrozole was dissolved in 1% CMC (Carboxymethyl cellulose) as a vehicle solvent and then the desired volume was added to obtain the desired concentration of 1 mg kg–1. Melatonin solution was prepared by dissolving it in a few drops of ethanol and then diluted with normal saline upto the desired concentration.
Experimental design: After an acclimatization period of 2-3 weeks, rats were randomly divided into following experimental groups:
| Group I | : | Normal control (1% CMC, p.o.) |
| Group II | : | PCOS control (letrozole -1 mg kg–1 p.o.) |
| Group III | : | Mel (per se) (200 μg/100 g i.p) |
| Group IV | : | PCOS (letrozole-1 mg kg–1 p.o.)+Mel (200 μg/100 g i.p) |
Vaginal smear observation: The PCOS condition was confirmed by preparation of vaginal smear to observe reproductive cycle irregularities. Vaginal secretions were collected daily with a plastic pipette by inserting the tip in the vagina of female rats, filled with 15 μL of normal saline. One drop of collected vaginal fluid was placed on glass slides. Vaginal fluid was fixed by placing the slides on a slide warming table and stained with methylene blue staining solution. After staining, slides were washed to remove the excess stain, dried and observed under light microscope to assess the changes in the estrus cycle. Three types of cells were recognized: Round and nucleated cells were epithelial cells; irregular cells without a nucleus were the cornified cells and the small round cells were the leucocytes, their mutual proportion was used for the determination of different phases of the estrus cycle15.
Sample collection: After completion of experimental period, the rats of all the groups were sacrificed by euthanized in desiccator by using diethyl ether as an anesthesia. The blood samples were collected directly through cardiac puncture in heparinized tubes and centrifuged at 3000 g for 15 min to collect the plasma. The blood plasma was stored at -20°C for serum cytokine assay. The ovaries were collected and lysed for the assessment of receptor expression of various receptors.
Serum cytokine analysis
Circulatory level of IL-6: Sandwich ELISA was performed to quantify the level of IL-6 in serum collected from the rats according to the manufacturer’s instruction (Koma Biotech, Seoul, Korea; Cat. No. K0331230). Lower and upper limits of analytic sensitivities were 16 and 1000 pg mL–1. All the assays were carried out in triplicate.
Circulatory level of TNF-α: Sandwich ELISA was performed to quantify the level of TNF-α in serum collected from the rats according to the manufacturer’s instruction (Koma Biotech; Cat. No. K0331186). Lower and upper limits of analytic sensitivities were 16 and 2000 pg mL–1. All the assays were carried out in triplicate.
Receptor assay
qRT-PCR: The qRT-PCR was done for the expression assay of cytokine (IL-2R and IL-6R) by first extracting the total mRNA, followed by cDNA synthesis. The primers were purchased from Imperial life sciences (P) limited (Table 1).
RNA Isolation: Total RNA was isolated from the frozen tissue samples. Tissues were slowly crushed with the help of mortar and pestle. About 1 mL of RIBOZOL (Trizole) was added into it and homogenized for 5 min. The tissue samples homogenized in RIBOZOL was transferred to a fresh microtube and left for 5 min at RT. After incubation, 200 μL of chloroform was added, vertexed and left it for 10 min on RT, then centrifuged at 13000 rpm for 10 min at 4°C. Supernatant was collected in a fresh microtube and 0.5 mL isopropanol was added, vortexed and kept it for 10 min on RT. The sample was centrifuged at 13000 RPM on 4°C. The supernatant was discarded and the pellet was washed with 80% ethanol by again centrifuging it on 7500 rpm for 5 min at 4°C. Washed pellet was air dried and dissolved in 50 μL of DEPC water and aliquoted it into 5 tubes and kept in -80°C for further processing.
cDNA synthesis: The cDNA synthesis was done by using Thermo kit K1632. About 1 μL of total RNA isolated from the tissues were used as template where 1 μL of primer oligo (dT) 18 and 10 μL of nuclease-free water was added. It was mixed gently, centrifuged briefly and incubated at 65°C for 5 min. Then the sample was transferred on ice, spins down and places the vial back on ice.
Then, 4 μL of 5X Reaction Buffer, 1 μL of RiboLock RNase Inhibitor (20 U μL–1), 2 μL of 10 mM dNTP Mix and 1 μL of Revert Aid H Minus M-MuLV Reverse Transcriptase (200 U μL–1) were added. Total volume of the reaction mixture was 20 μL. It was mixed gently and spin down. Finally for cDNA synthesis, the reaction mixture was incubated at 42°C for 60 min.
Western blot analysis: The western blot analysis was performed according to the method of Treek et al.16. Ovarian tissues were dissected in chilled PBS, homogenized and lysed in lysate buffer. The protein content of the lysates was quantified using the Bradford method17. The membranes were immunostained with β-actin antibodies in 1:1000 dilutions (A-2228; Sigma-Aldrich) as internal loading control. Immune detection of β-actin was performed with donkey anti-mouse IgG-HRP (1:1000). Bands were quantified by the measurement of optical density using Scion Image Analysis Software (Scion Corporation, MD, USA). Values were expressed as the ratio of the density of the specific signal to the β-actin signal. The ratio of density was calculated with respect to β-actin (house-keeping gene) and expressed as percentage relative integrated density values of MT1, MT2 and ER-α.
Statistical analysis: The experimental data were expressed as Mean±SE. Statistical analysis was performed using one-way ANOVA followed by Student’s t-test with the SPSS 16.0 statistical software (SPSS, Chicago, IL, USA). A probability value p<0.05 (*) or p<0.01 (**) was considered to be statistically significant.
| Table 1: | List of primer |
 | |
RESULTS
Expression of melatonin receptors MT1 and MT2: The MT1 receptor expression was noted significantly down regulated during PCO condition. Whereas, administration of exogenous melatonin resulted in the upregulation of expression level of MT1 significantly. A significantly decreased expression was observed in MT2 receptor, but the administration of melatonin showed recovery pattern for the expression towards the normal significantly (Fig. 1, 2).
Expression of estrogen receptor–ER-α: A significant down regulation in expression of ER-α receptors on ovarian tissue of PCO rats was noted. Administration of exogenous melatonin resulted in restoration of the disruption of expression level. Treatment of melatonin to PCO rats resulted with significant reversal of receptor of ER-α (Fig. 3).
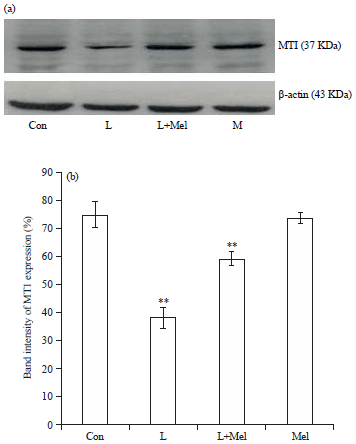 | |
| Fig. 1(a-b): | Western blot analysis for variation in the expression of melatonin receptor MT1 in ovary of female rats. The data are expressed as percentage band intensity of receptor expression in ovary. β-actin expression was used as control. Data are expressed as Mean±SE, n = 6 females per group |
| Con: Control, L: Letrozole, Mel: Melatonin,**p<0.01, Con vs. L, L vs. L+Mel | |
Expression of cytokines-IL-2R and IL-6R: The IL-2R expression pattern was noted down regulated in letrozole induced PCO rats. Treatment of melatonin showed a significant increase (Fig. 4). Expression of IL-6R was found significantly reduced in PCO induced rats which was restored towards the control group on administration of exogenous melatonin (Fig. 5).
Circulatory level of cytokines: Circulatory level of cytokine IL-6 increased significantly in PCO rats when compared to the control group of rats (Fig. 6). The circulatory level of TNF-α was noted significantly increased in PCOS rats compared to the control group of rats. However, after the administration of melatonin restoration in the level was found significant towards the controlled group (Fig. 7). Treatment of melatonin resulted in the restoration of the circulatory cytokines (TNF-α and IL-6) level significantly and comparable to the controlled group.
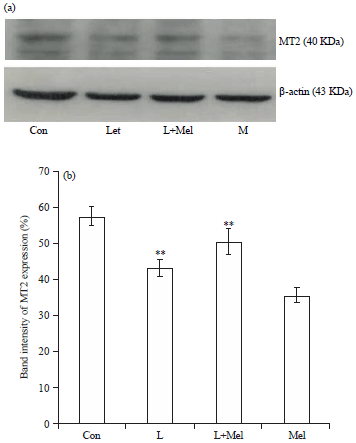 | |
| Fig. 2(a-b): | Western blot analysis for variation in the expression of melatonin receptor MT2 in ovary of female rats. The data are expressed as percentage band intensity of receptor expression in ovary. β-actin expression was used as control. Data are expressed as Mean±SE, n = 6 females per group |
| Con: Control, L: Letrozole, Mel: Melatonin, **p<0.01, Con vs. L, L vs. L+Mel | |
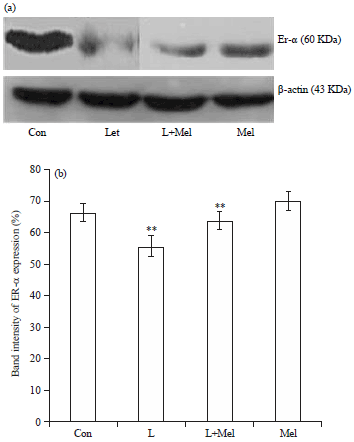 | |
| Fig. 3(a-b): | Western blot analysis for variation in the expression of estrogen receptor ER-a in ovary of female rats. The data are expressed as percent band intensity of receptor expression in ovary. ß-actin expression was used as control. Data are expressed as mean ±SE, N= 6 females per group |
| Con: Control, L: Letrozole, Mel: Melatonin, **p<0.01; Con vs. L, L vs. L+Mel | |
DISCUSSION
Present finding clearly showed the presence of MT1 and MT2 receptors in ovarian tissue of rat through which melatonin can exert its influence the reproductive system. Further ovarian tissue of PCO rats demonstrates down regulation of MT1 and MT2 receptors and melatonin treatment resulted in the restoration of the MT1 and MT2 receptors. Our observation confirms influence of exogenous melatonin and its receptor (MT1 and MT2) during such heterogeneous endocrine disorder of PCO which indicates the sensitivity of HPO axis for the treatment of melatonin. The expression of ER-α was significantly decreased in polycystic ovarian tissues.
Expression of ER-α receptor following administration of melatonin alone appeared static when compared to each other.
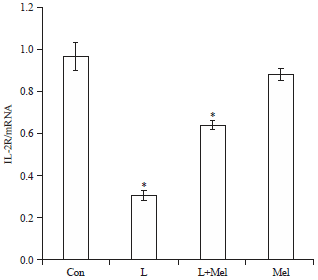 | |
| Fig. 4: | Receptor expression analysis for variation in the expression of cytokine receptor IL-2R in ovary of female rats. The data are expressed as mRNA expression in ovary. Data are expressed as Mean±SE, n = 6 females per group |
| Con: Control, L: Letrozole, Mel: Melatonin, *p<0.05, Con vs. L, L vs. L+Mel | |
 | |
| Fig. 5: | Receptor expression analysis for variation in the expression of cytokine receptor IL-6R in ovary of female rats. The data are expressed as mRNA expression in ovary. Data are expressed as Mean±SE, m = 6 females per group |
| Con: Control, L: Letrozole, Mel: Melatonin, *p<0.05, **p<0.01, Con vs. L, L vs. L+Mel | |
The present finding can be correlated with the fact that during PCO condition there is a failure to upregulation of aromatase enzyme in granulose cell therefore, estradiol concentration of follicles fails to increase adequately and hence, led to the abnormal expression of ER-α18.
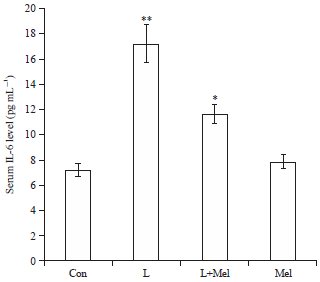 | |
| Fig. 6: | Variations in the level of serum cytokine IL-6 in letrozole induced PCO rats and in recovery by melatonin. Data are expressed as Mean±SE, N = 6 females per group |
| Con: Control, L: Letrozole, Mel: Melatonin, *p<0.05, **p<0.01, Con vs. L, L vs. L+Mel | |
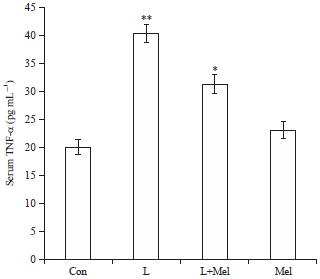 | |
| Fig. 7: | Variations in the level of serum cytokine TNF-α in letrozole induced PCO rats and its recovery by melatonin. Data are expressed as Mean±SE, N = 6 females per group |
| Con: Control, L: Letrozole, Mel: Melatonin, *p<0.05, **p<0.01, Con vs. L, L vs. L+Mel | |
Further, present findings can be justified on the basis of the previous reports suggesting inhibitory action of melatonin during the binding of E2 ER complex to the estrogen responsive element on DNA19,20.
Further, a differential expression of MT1 receptor noted in all experimental groups of rats where letrozole induced PCOs female showed down regulation in MT1 R expression in ovary. Upregulation of MT1 R in ovarian tissue following melatonin injection to the control group provide a clear cut evidence of direct involvement of MT1 receptor in ovarian physiology specifically during the pathogenicity of PCOS.
Letrozole induced PCO female rats showed a significant increase in serum cytokine level of IL-6 as well as TNF-α in letrozole induced PCO rats indicting the internal inflammation which could be due to the tissue specific pathogenicity of local micro environment and stress induced. These finding is in concurrence with the previous report suggesting that free radical generation act via a variety of proinflammatory cytokines during various pathological condition of female reproduction including polycystic ovarian disease (PCOD)19. Since melatonin is reported a strong immunomodulatory which could be the main factor for the restoration in immune status of ovarian tissue. The present finding provides a clear cut evidence of involvement of exogenous melatonin resulted in the restoration of cytokine receptors as that of the control. Action of melatonin can be explained as this neurohormone can activate or deactivate the signaling pathway through its receptor MT1 and MT2 and also by upregulating few cytokines like IL-2 and others (IL-6, TNF-α) in seasonal breeder21,22.
Previous studies provide the evidence that estrogen receptor are localized in the small antrum of granulosa cell and dominant follicles but not in the theca cells23,24. Down regulation in ER-α receptor expression in ovarian tissue of PCO rats appears as one of the responsible component resulted into the arthymicity in the pulsatility of LH and FSH and could be because of the disturbances in the LH surge. Further, reports on ER knockout animals revealed that ER-α is a prerequisite for the proper functioning of hypothalamic pituitary ovarian (HPO) axis and therefore regulates the successful ovulation25 and hence support partially the present finding. Knockout ER-α mice showed a similar phenotype to that of PCO with high circulating LH concentration which coincides our present finding. Such kind of hormonal balancing during the PCO may be considered as one of the major cause leading in the disarrangement of feedback mechanism and ultimately characterized by impaired ovulation such as PCOS with alteration in the expression of ER-α26.
The ER-α receptors are present in the ovary differently in ovarian-theca and granulosa cells27. ER-α, mRNA and protein which are expressed minimally in the ovary are located predominantly in the theca layer27. The ER-β mRNA and protein which are highly expressed in the granulosa cells of the developing follicles. The disruption in the expression of ER-α in present findings can be explained because of elevated circulatory LH serum levels during PCO due to failure of the negative feedback action of estradiol on the hypothalamic-pituitary axis.
Nonetheless, since cytokine usually participate in ovarian physiology as paracrine/autocrine regulators of steroidogenesis, production of prostaglandins and LH, FSH receptor induction28-30. Literature suggested regarding origin of cytokines either leucocytes produced during ovulation or from resident ovarian cells31. Since cytokines are signaling molecule therefore may be involved in upregulating/ downregulating particular cytokines like IL-2 and IL-6 in serum and additionally by associating its own receptor with cytokine receptor. Presence and participation of IL-6 is reported in normal ovary where it may participate in follicular development by reducing the hormone binding follicle stimulating hormone capacity of granulose cell32. Therefore, exogenous melatonin given to PCO rats showed synchronization in the cytokine circulatory serum (IL-6, TNF-α) and receptor expression (IL-2R, IL- 6R) via LH/FSH and hence modulating the ER-α expression with a crosstalk among MT1 and MT2 receptor. TNF α again is a pro inflammatory cytokine associated with the regulation of the tumor growth by increasing production of IL-6. TNF-α along with IL-1 inhibits the secretion of progesterone. The TNF α may also be involved in regression of corpora lutea32. Atresia is a hormonally controlled form of apoptosis of ovarian follicles and cytokines play a role in its regulation.
CONCLUSION
The present observation clearly explained that melatonin may have modulatory effect on steroidogenesis via its receptors (MT1/MT2) in association with ER-α in modulation of ovarian function during PCO condition. Present study further showed that during PCO disorder the molecular alterations might have brought the changes in steroidogenesis associated gene expression which following the treatment of melatonin was reversed via the crosstalk of the MT1/MT2, ER-α and the cytokines IL-2R and IL-6R.
SIGNIFICANCE STATEMENT
The findings of this study evidenced a novel therapeutic approach using melatonin exogenously. Alterations at molecular level which are observed during the PCOS pathogenicity might have changed the steroidgenic pathway related expression of genes. Howover, these alterations were reversed following melatonin supplementation. This study will help the researchers to uncover certain areas that many researchers have not been able to explore.
ACKNOWLEDGMENTS
Authors greatly acknowledge DBT BUILDER program, Department of Biotechnology, Ministry of Science and Technology, Government of India, New Delhi (Sanction No. BT/PR7020/INF/22/172/2012) for financial assistance and Department of Zoology, Guru Ghasidas Vishwavidyalaya for providing the laboratory facilities.
REFERENCES
- Skrtic, A., L. Sokolic, A. Borovecki, J. Rosa and V. Fenzl, 2011. Immunohistochemical localization of CD31, NOTCH1 and JAGGED1 proteins in experimentally induced polycystic ovaries of immature rats. Acta Histochem., 113: 262-269.
CrossRefDirect Link - Baravalle, C., N.R. Salvetti, G.A. Mira, N. Pezzone and H.H. Ortega, 2006. Microscopic characterization of follicular structures in letrozole-induced polycystic ovarian syndrome in the rat. Arch. Med. Res., 37: 830-839.
CrossRefDirect Link - Dasgupta, S. and B.M. Reddy, 2008. Present status of understanding on the genetic etiology of polycystic ovary syndrome. J. Postgraduate Med., 54: 115-125.
CrossRefDirect Link - Guttmann-Bauman, I., 2005. Approach to adolescent polycystic ovary syndrome (PCOS) in the pediatric endocrine community in the USA. J. Pediatr. Endocrinol. Metab., 18: 499-506.
CrossRefDirect Link - Masana, M.I., Jr.J.M. Soares and M.L. Dubocovich, 2005. 17β-estradiol modulates hMT1 melatonin receptor function. Neuroendocrinology, 81: 87-95.
CrossRefDirect Link - Reiter, R.J., D.X. Tan, L.C. Manchester, S.D. Paredes, J.C. Mayo and R.M. Sainz, 2009. Melatonin and reproduction revisited. Biol. Reprod., 81: 445-456.
CrossRefDirect Link - Ronnberg, L., A. Kauppila, J. Leppaluoto, H. Martikainen and O. Vakkuri, 1990. Circadian and seasonal variation in human preovulatory follicular fluid melatonin concentration. J. Clin. Endocrinol. Metab., 71: 492-496.
PubMedDirect Link - Dubocovich, M.L. and M. Markowska, 2005. Functional MT1 and MT2 melatonin receptors in mammals. Endocrine, 27: 101-110.
CrossRefDirect Link - Reiter, R.J., J.C. Mayo, D.X. Tan, R.M. Sainz, M. Alatorre-Jimenez and L. Qin, 2016. Melatonin as an antioxidant: Under promises but over delivers. J. Pineal Res., 61: 253-278.
CrossRefDirect Link - Rai, S., M. Basheer, H. Ghosh, D. Acharya and Y.A. Hajam, 2015. Melatonin attenuates free radical load and reverses histologic architect and hormone profile alteration in female rat: An in vivo study of pathogenesis of letrozole induced poly cystic ovary. J. Clin. Cell. Immunol., Vol. 6.
CrossRefDirect Link - Rai, S., Y.A. Hajam, M. Basheer and H. Ghosh, 2016. Biochemical and histopathological inflections in hepato-renal tissues of streptozotocin (STZ) induced diabetic male rats: Impact of exogenous melatonin administration. J. Clin. Res. Bioeth., Vol. 7.
CrossRefDirect Link - Carrillo-Vico, A., P.J. Lardone, N. Alvarez-Sanchez, A. Rodriguez-Rodriguez and J.M. Guerrero, 2013. Melatonin: Buffering the immune system. Int. J. Mol. Sci., 14: 8638-8683.
CrossRefDirect Link - Seema, R. and C. Haldar, 2003. Pineal control of immune status and hematological changes in blood and bone marrow of male squirrels (Funambulus pennanti) during their reproductively active phase. Comp. Biochem. Physiol. Part C: Toxicol. Pharmacol., 136: 319-328.
CrossRefPubMedDirect Link - Haldar, C., S. Rai and R. Singh, 2004. Melatonin blocks dexamethasone-induced immunosuppression in a seasonally breeding rodent Indian palm squirrel, Funambulus pennanti. Steroids, 69: 367-377.
CrossRefPubMedDirect Link - Bilal, A., N. Jahan, A. Ahmed, S.N. Bilal and S. Habib, 2013. Antifertility activity of hydroalcoholic extract of Ocimum basilicum Linn. leaves on female wistar rats. J. Reprod. Contraception, 24: 45-54.
CrossRefDirect Link - Treeck, O., C. Haldar and O. Ortmann, 2006. Antiestrogens modulate MT1 melatonin receptor expression in breast and ovarian cancer cell lines. Oncol. Rep., 15: 231-235.
CrossRefDirect Link - Bradford, M.M., 1976. A rapid and sensitive method for the quantitation of microgram quantities of protein utilizing the principle of protein-dye binding. Anal. Biochem., 72: 248-254.
CrossRefPubMedDirect Link - Stocco, C., 2008. Aromatase expression in the ovary: Hormonal and molecular regulation. Steroids, 73: 473-487.
CrossRefDirect Link - Rato, A.G., J.G. Pedrero, M.A. Martinez, B. Del Rio, P.S. Lazo and S. Ramos, 1999. Melatonin blocks the activation of estrogen receptor for DNA binding. FASEB J., 13: 857-868.
CrossRefDirect Link - Lopes, J., D. Arnosti, J.E. Trosko, M.H. Tai and D. Zuccari, 2016. Melatonin decreases estrogen receptor binding to estrogen response elements sites on the OCT4 gene in human breast cancer stem cells. Genes Cancer, 7: 209-217.
CrossRefPubMedDirect Link - Ahmad, R. and C. Haldar, 2010. Photoperiodic regulation of MT1 and MT2 melatonin receptor expression in spleen and thymus of a tropical rodent Funambulus pennant during reproductively active and inactive phases. Chronobiol. Int., 27: 446-462.
CrossRefDirect Link - Carrillo-Vico, A., J.M. Guerrero, P.J. Lardone and R.J. Reiter, 2005. A review of the multiple actions of melatonin on the immune system. Endocrine, 27: 189-200.
CrossRefDirect Link - Iwai, T., Y. Nanbu, M. Iwai, S. Taii, S. Fujii and T. Mori, 1990. Immunohistochemical localization of oestrogen receptors and progesterone receptors in the human ovary throughout the menstrual cycle. Virchows Arch. A, 417: 369-375.
CrossRefDirect Link - Horie, K., K. Takakura, H. Fujiwara, H. Suginami, S. Liao and T. Mori, 1992. Immunohistochemical localization of androgen receptor in the human ovary throughout the menstrual cycle in relation to oestrogen and progesterone receptor expression. Hum. Reprod., 7: 184-190.
CrossRefDirect Link - Couse, J.F. and K.S. Korach, 1999. Reproductive phenotypes in the estrogen receptor-alpha knockout mouse. Ann. D'endocrinol., 60: 143-148.
Direct Link - Jayes, F.L., K.A. Burns, K.F. Rodriguez, G.E. Kissling and K.S. Korach, 2014. The naturally occurring luteinizing hormone surge is diminished in mice lacking estrogen receptor beta in the ovary. Biol. Reprod., 90: 1-9.
CrossRefDirect Link - Jakimiuk, A.J., S.R. Weitsman, H.W. Yen, M. Bogusiewicz and D.A. Magoffin, 2002. Estrogen receptor α and β expression in theca and granulosa cells from women with polycystic ovary syndrome. J. Clin. Endocrinol. Metab., 87: 5532-5538.
CrossRefDirect Link - Adashi, E.Y., 1990. The potential relevance of cytokines to ovarian physiology: The emerging role of resident ovarian cells of the white blood cell series. Endocrine Rev., 11: 454-464.
CrossRefDirect Link - Wang, L.J. and R.J. Norman, 1992. Concentrations of immunoreactive interleukin-1 and interleukin-2 in human preovulatory follicular fluid. Hum. Reprod., 7: 147-150.
CrossRefDirect Link - Wang, L.J., M. Brannstrom, S.A. Robertson and R.J. Norman, 1992. Tumor necrosis factor α in the human ovary: Presence in follicular fluid and effects on cell proliferation and prostaglandin production. Fertil. Steril., 58: 934-940.
CrossRefDirect Link - Brannstrom, M., G. Mayrhofer and S.A. Robertson, 1993. Localization of leukocyte subsets in the rat ovary during the periovulatory period. Biol. Reprod., 48: 277-286.
CrossRefDirect Link - Machelon, V., D. Emilie, A. Lefevre, F. Nome, I. Durand-Gasselin and J. Testart, 1994. Interleukin-6 biosynthesis in human preovulatory follicles: Some of its potential roles at ovulation. J. Clin. Endocrinol. Metab., 79: 633-642.
CrossRefDirect Link