
Jayadi
Department of Aquaculture, Faculty of Fishery and Marine Science, Indonesia Muslim University, Makassar-90231, South Sulawesi, Indonesia
LiveDNA: 62.16564
Amrah Husma
Department of Aquaculture, Faculty of Fishery and Marine Science, Indonesia Muslim University, Makassar-90231, South Sulawesi, Indonesia
Ilmia
Department of Aquaculture, Faculty of Fishery and Marine Science, Indonesia Muslim University, Makassar-90231, South Sulawesi, Indonesia
Pakistan Journal of Biological Sciences
Year: 2017 | Volume: 20 | Issue: 7 | Page No.: 336-342







ABSTRACT
Background and Objective: Marosatherina ladigesi, including native fish in South Sulawesi Indonesia, are vulnerable and must be preserved. The objective of the study was to examine known food consumption, survival, growth and the activities of digestive enzymes of M. ladigesi larvae. Material and Methods: The larvae were obtained from natural spawning and the mixed live feed: Infusoria sp., Daphnia sp. and Artemia salina were given twice a day. Feed intake, growth, survival, activities of protease, lipase and amylase enzymes were observed in ages 3, 5, 10, 15, 20, 25, 30, 35 Days after Hatching (DAH). Data were analyzed by following a complete randomized trial design. Results: Activities of protease, amylase and lipase enzymes have been detected since 3 DAH. The protease enzyme activity was obtained at 0.438 U mL min–1 at 35 DAH, while amylase and lipase activities fluctuated. Activities of amylase and lipase reached 0105 and 0.093 U mL–1 min–1 at 15 DAH, respectively. The activity of the enzyme increased at the same age of 10-15 DAH larvae entering a phase of flexion. Conclusion: It was concluded that the use of mixed live feed Infusoria sp., Daphnia sp. and A. salina contribute to enzyme protease, lipase and amylase activities during the ontogeny stages of M. ladigesi. The highest activity of protease suggests that the fish have carnivorous feeding habits. The results of this study can be used to improve the maintenance technique of M. ladigesi larvae in order to accelerate growth and improve survival as an effort to increase the production of M. ladigesi.
PDF Abstract XML References Citation
Received: March 06, 2017;
Accepted: May 22, 2017;
Published: June 15, 2017
Copyright: © 2017. This is an open access article distributed under the terms of the creative commons attribution License, which permits unrestricted use, distribution and reproduction in any medium, provided the original author and source are credited.
How to cite this article
Jayadi, Amrah Husma and Ilmia, 2017. Enzyme Digestive Activities During Ontogeny in Marosatherina ladigesi Larvae. Pakistan Journal of Biological Sciences, 20: 336-342.
DOI: 10.3923/pjbs.2017.336.342
URL: https://scialert.net/abstract/?doi=pjbs.2017.336.342
DOI: 10.3923/pjbs.2017.336.342
URL: https://scialert.net/abstract/?doi=pjbs.2017.336.342
INTRODUCTION
Marosatherina ladigesi’s original habitat is in the freshwater of South Sulawesi, Indonesia1-2. The condition of M. ladigesi in their natural habitat has damaged3 the genetic diversity near by Jayadi et al.4 and the reproductive capacity is declining5. This freshwater ornamental fish was already in the category of at threat of extinction listed by the International Union for Conservation of Nature since 19906. One of the efforts that needs to be made for the preservation of M. ladigesi is domestication. These fish larvae have been successfully cultivated under controlled conditions3,7,8. On the other hand, the survival rate of larvae is still low at the early stage of their life. The M. ladigesi survival rate has been reported as 38.31-39.02% at age 5 DAH8.
The transition from endogenous to exogenous feeding is the critical phase in the early stage of M. ladigesi life. High mortality rates of the fish larvae were found due to the inadequate nutrition after absorption of the yolk-sac and the first feeding of larvae9,10. Furthermore, the food eaten by the larvae does not correspond with the development of digestive organs and digestive enzyme activity11-13. The ability of fish to consume food depends on the completeness of the digestive organs and digestive enzymes14-17.
Digestive physiology of larvae is one of the most important aspects that directly influences the development, survival and the growth of fish16,18. It can be used as information to identify the capabilities of larvae in digestion and absorption of the protein and lipid content of the food for optimizing the survival and growth of the larval rearing19.
The Information on the enzyme activity during the initial phase of larval development can be used as guidance in the artificial feeding management13,15,16. The knowledge of enzyme activity at the early stage of larval development or ontogeny can be a guidance in managing proper feeding during these developments14,15,20,21. The information related to activities of digestive enzymes such as protease, lipase and amylase during the early life history of larvae M. ladigesi has not been reported. Therefore, there is a need to investigate the activity of digestive enzymes M. ladigesi larvae during ontogeny by providing a mix of natural food such as Infusoria sp., Daphnia sp. and Artemia salina under controlled conditions for 35 DAH.
MATERIALS AND METHODS
Larvae rearing: This study was conducted from March-October, 2016 at the freshwater hatchery unit for ornamental fish in the Mangala Municipality Makassar, Indonesia. Larval rearing methods were performed by Jayadi et al.8, by providing a larval rearing container using a fibreglass tank with dimensions of 60 cm length×40 cm width×35 cm height. The volume of water in the rearing tank was approximately 20 L. A water change was regularly conducted every week, with 25% of the water volume per tank. The live feeds in the form of Infusoria sp., Daphnia sp. and nauplii of Artemia salina were given during the experiment. The feeding frequency was twice daily.
Sampling of larvae: The observation of the type and amount of eaten food, growth, survival, activities of protease, lipase and amylase enzymes were conducted at 3, 5, 10, 15, 20, 25, 30 and 35 DAH. Five fish larvae were randomly taken to determine the type and amount of food consumed. The intestines of larvae were dissected under a dissecting microscope (Olympus CX23, Hamburg, Germany). Growth measurements were performed by measuring the total length, from the tip of the snout to the posterior margin of the body of larvae. Linear regression was applied for fish growth analysis.
Analysis of digestive enzymes: For enzyme analysis, the fish samples were taken in the morning before feeding. To obtain the supernatant, 2.0-3.0 mg samples were homogenized (Wiggen Hauser D500 homogenizer, Berlin, Germany) in Tis-HCl buffer 50 mM, 20 mM CaCl2 at pH 7.5 and centrifuged at 12,000 rpm for 15 min. The resulting supernatant was stored in the freezer at -80°C22-24. Protease activities were analysed following the method of Bergmeyer et al.22, 2% casein substrates were mixed with 0.01 M borate buffer (pH 8). The mixture was incubated at 37°C for 10 min, added with 0.1 M TCA and re-incubated at 37°C for 10 min. The mixture was centrifuged at 4000 rpm for 10 min, added with 5.0 mL Na2CO3 and 1 mL reagent Follin (1:2). The absorbance was measured using spectrophotometer (UV-1800, Shimadzu Europa, Duisburg, Germany) at 578 nm. Amylase activities were analyzed according to the method of Bernfield23. The reaction mixture consisted of starch solution 1% and citra buffer solution (pH 5.7). The mixture was incubated at 20°C for 30 min, added with 2 mL DNS and boiled for 5 min. The absorbance was read on a spectrophotometer (UV-1800, Shimadzu Europa, Duisburg, Germany) at 550 nm. Analysis of lipase activity was based on Borlongan24. The analysis was performed by adding 1.5 mL olive oil and 1.5 mL of 0.1 M HCl tri buffer at pH 8.0. The mixture was incubated for 6 h at a temperature of 37°C, followed by the addition of 3 mL ethyl alcohol (95%). The mixture was titrated with 0.01 N NaOH using 0.9% thymolphthalein in ethanol as an indicator.
Statistical analysis: All measurements were carried out in triplicate. Results presented are ±SD. Data were checked for homogeneity of variance using Levene’s test. Data of live food consumed were analysed by Statistical software version 6.1. Survival was compared by Fischer’s chi-square test. Data of length were analysed by using appropriate regression model analysis. Enzymatic activity data were analysed using one-way ANOVA and means were compared with a Duncan’s test. The level of significance was set at p<0.05. Statistical analyses were done using SPSS 16.025.
RESULTS
Diversity of live foods consumed by M. ladigesi larvae during maintenance 35 DAH is detailed in Fig. 1. Larvae started feeding on Infusoria sp., Daphnia sp. and A. salina from 3-35 DAH. Different percentages of Infusoria sp., Daphnia sp. and A. salina were taken by M. ladigesi larvae according to DAH. Based on the proximate analysis, the nutritional values of mixed food (Infusoria sp., Daphnia sp. and A. salina) were 61.47% protein, 15.46% lipid and 7.56% ash. Meanwhile, the analysis of body protein fish at 35 DAH was 54.64%, while the lipid and ash contents were 14.57 and 7.79%, respectively.
Survival rates and growth of M. ladigesi larvae given various types of natural feed during maintenance period 35 DAH are shown in Fig. 2a and b. The survival of the M. ladigesi larvae at the age of 5 DAH decreases then increases with age (Fig. 2a). The total length of newly hatched larvae was observed from 4.45-5.21 mm (Fig. 2b). Slower growth of larvae as indicated by the total length was observed from 5-10 DAH. However, faster growth rates were observed after 10 DAH. The total length reached 10.12-10.57 mm at 35 DAH. Faster growth of larvae occurred from 15-35 DAH. M. ladigesi larval growth pattern positively correlated with the increase of age with the equation of y = 0.169x+3870, R2 = 0.936 (Fig. 2b).
The pattern of digestive enzyme activity of larvae M. ladigesi given different types of natural food is shown in Fig. 3. Protease, amylase and lipase have been detected since the age of 3 DAH then increased rapidly at the age of 10 DAH. Protease enzyme activity in M. ladigesi larvae is higher than amylase and lipase. Enzyme activity patterns of protease tended to increase with age. Whereas lipase and amylase enzyme activity was lower (tended not to increase) and decreased after age 20 DAH. The increased of enzyme activity recorded from 10-15 DAH was associated with the entry of larvae into the flexion phase.
DISCUSSION
The results of this study have found that the maintenance of M. ladigesi larvae using Infusoria sp., Dapnia sp. and A. salina as mixed live food has achieved a 100% survival rate at 35 DAH and also provides a rapid growth rate with the increasing age of the larvae. It shows that the food given is very suitable for the development of M. ladigesi larvae. However, the percentage of food amounts (Infusoria sp., Dapnia sp. and A. salina) eaten by larvae at each age varies depends on the size of the mouth opening. After the larvae of 15 DAH showed increased digestive enzyme activity, the most increased activity is the protease enzyme, which shows that M. ladigesi larvae at that age may have been given artificial food.
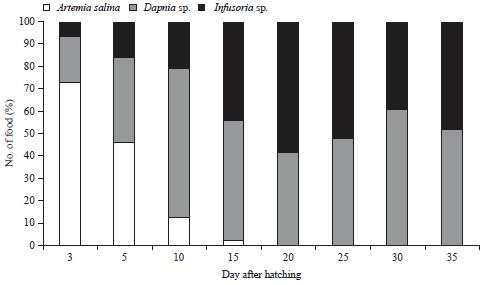 | |
| Fig. 1: | Percentage of the live food consumed by M. ladigesi larvae at various ages |
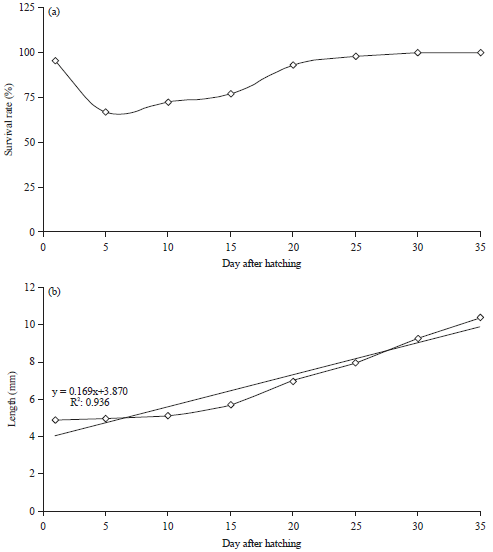 | |
| Fig. 2(a-b): | (a) Survival and (b) Length/growth rate M. ladigesi larvae at various ages |
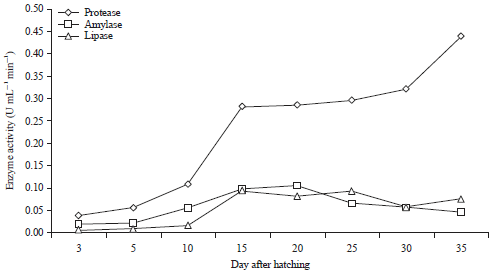 | |
| Fig. 3: | Activities of protease, amylase and lipase enzymes at various ages |
Feeding habits of fish are important to identify during larvae stage. Unsuitable live food in the development of the morphology of fish larvae may lead to high mortality19. The successful fish feeding during the nutritional transition process from the phases endogenous to exogenous on fish larvae increases the survival and growth rate26. Live food is suitable in the transition stage due to its complete nutritional value and can generate digestive enzymes15,20,27. In the present study, larvae began to feed on live food at 3 DAH. The mixed live food was eaten by the larvae. Nonetheless, the amount of food eaten varied in the different age groups. Fish larvae selectively take food based on the size of their mouth opening9. The ability of fish larvae to take live food at the time before or after the egg yolk runs out is different in each fish. M. ladigesi takes live food before the food reserves are depleted8. This indicates that the fish consumed a mixture of endogenous and exogenous at 5 DAH.
The survival rate of M. ladigesi at 5 DAH reached 66.43% (Fig. 2), which was higher than the survival rate of the larvae fed on Daphnia sp. (38.31-39.02%)8. The survival rate of larvae at 10 DAH was 72.19% and increased to 100% until 35 DAH (Fig. 2a). The survival rate of the larvae increased with increasing age. Growth performance of the larvae positively increased with increasing age (y = 0.169x+3870, R2 = 0.936) (Fig. 2b). This showed that the combination of live food may be suitable in terms of quality and quantity. The nutritional values of the mixed live food, such as Infusoria sp., Daphnia sp. and A. salina were 61.47% protein, 46% lipid and 7.56% ash, respectively. The proximate analysis of experimental fish larvae at age 35 DAH was 54.64% protein, 14.57% lipid and 7.79% ash. The nutritional composition of the live food and the proximate analysis of the fish’s body met the nutritional requirement for M. ladigesi larvae for their growth and development. Similarly, it was found that the fish can have faster growth if the nutritional value of the food meets the energy requirement for growth28,29. The mixture of live food such as Infusoria, Daphnia sp. and A. salina may support better growth performance.
The enzyme activity patterns of protease, amylase and lipase in M. ladigesi larvae are shown in Fig. 3. The response of enzyme activity can be affected by the amount and quality of food nutrients consumed30. The enzyme activities are positively correlated with dietary habits31. The live food eaten by larvae contribute to the increased activity of enzymes in the digestive system and it is recommended to feed larvae before and after the yolk disappears9,13,31-33. The larvae at 10-35 DAH tended to consume Daphnia sp. and A. salina (Fig. 1). Furthermore, it increased the response of enzyme activity protease, amylase and lipase (Fig. 3), which led to the increase in growth (Fig. 2b). Moreover, the increased activity of the enzyme also is due to the completeness of enzyme-producing organs13. The activity of protease, lipase and amylase enzymes may indicate that M. ladigesi larvae were capable of digesting protein, carbohydrates and fat from the onset of exogenous feeding.
The activity of protease enzyme in M. ladigesi larvae has been detected at 3 DAH, when the larvae open their mouth. It is the presence of trypsin, the pancreatic enzyme that can activate other digestive enzymes14-16,21,34. The patterns of enzyme protease activity in M. ladigesi larvae increased with increasing age (Fig. 3). Enzyme activity patterns of protease found in the larvae are similar with Anabas testudineus21, Pangasius hypophthalmus34, Sciaenops ocellatus35 and Catla catla36.
Enzyme activity patterns of amylase M. ladigesi larvae are shown in Fig. 3. The activity of amylase enzyme increased from 0.019 U mL–1 min–1 at 3 DAH to 0.105 U mL–1 min–1 at 15-20 DAH and decreased to 0.046 U mL–1 min–1 at 25-35 DAH. Reduced activity of the enzyme amylase protein may be due to the increased body weight of the fish15. The high activity on amylase depends on the eating habits of fish38. The activity of amylase enzyme in M. ladigesi larvae resemble that in Rutilus frisii kutum13, P. hypophthalmus34, Pseudociaena crocea38 and Trichotus blochii39.
The pattern of lipase activity in M. ladigesi larvae shows increased lipase activity from 0.005 U mL–1 min–1 at 5 DAH (the phase of yolk-sac) to 0.093 U mL–1 min–1 at 15 DAH (the flexion phase), then decreased to approximately 0.056 U mL–1 min–1 at 30 DAH. Lipase activity patterns of M. ladigesi are similar to that in R. frisii kutum13, C. carpio larvae16. However, it is different with lipase enzyme activity patterns in fish A. testudineus24 and M. nemurus15.
The understanding of enzyme activity is helpful for designing the food formula for each life stage of fish. High activity of the enzyme can be used as a basic indicator to determine the appropriate time of the artificial feeding of larvae, in which the enzyme capacity is physiologically ready to process food from the outside40. The enzyme activity was found to be high after 30 DAH of larvae. Hence, the artificial feeding should be conducted after that age.
CONCLUSION
The use of mixed live feed Infusoria sp., Daphnia sp. and A. salina contribute to enzyme protease, lipase and amylase activities during the ontogeny stages of M. ladigesi. The highest activity of protease suggests that the fish larvae have carnivorous feeding habits. The development of digestive enzyme activity can be used as a consideration for providing artificial feeding possibilities after the age of 15 DAH.
SIGNIFICANCE STATEMENTS
This study discovers the activities of enzyme protease, lipase and amylase in M. ladigesi larvae that can be beneficial for improving larval rearing techniques of M. ladigesi. This study will help the researcher to uncover the critical areas of the phase of ontogeny in M. ladigesi larvae that many researchers were not able to explore. Thus, a new theory of digestive enzyme activity of M. ladigesi larvae to 35 DAH has been found that protease enzyme increases with increasing larval age.
ACKNOWLEDGMENTS
The authors would like to thank the Director of Research and Community Service, Kemenristek-Dikti who provided research grants competing in the contract: DIPA-042.06-0/2016. We also thank to Muhammad Rajiman Hasbi as the head of Freshwater Ornamental Fish Hatchery in Manggala village, Makassar City as a place to conduct research. The authors express their gratitude to the head of research and development of brackish water aquaculture, Maros, for the use of laboratory facilities in analysing enzymes.
REFERENCES
- Parenti, L.R., 2011. Endemism and Conservation of the Native Freshwater Fish Fauna of Sulawesi, Indonesia, In: Proceedings of the 6th Fish National Seminar and 3rd Congress of the Indonesian Society of Ichthyologists, Simanjuntak, C.P.H., A. Zahid, M.F. Rahardjo, K.H. Hadiaty and Krismono et al. (Eds.). Indonesian Society of Ichthyologists, Cibinong, pp: 1-10.
- Kariyanti, S.B., A. Omar and J. Tresnati, 2014. Analysis of fecundity and egg diameter of Celebes rainbow fish (Marosatherina ladigesi Ahl, 1936) on the river Bantimurung and Pattunuang Asue, Maros, South Sulawesi. Proceedings of the National Symposium on Marine and Fisheries, May 3, 2014, Makassar, pp: 1-10.
- Jayadi, A. Husma, Nursahran, Ardiansyah and Sriwahidah, 2016. Domestication of Celebes rainbow fish (Marosatherina ladigesi). AACL Bioflux, 9: 1067-1077.
Direct Link - Yufera, M. and M.J. Darias, 2007. The onset of exogenous feeding in marine fish larvae. Aquaculture, 268: 53-63.
CrossRefDirect Link - Spirita, S.V. and A.J. Ahila, 2015. Stages of embryonic development of the Zebrafish Danio rerio (Hamilton). Eur. J. Biotechnol. Biosci., 3: 6-11.
Direct Link - Chen, B.N., J.G. Qin, S.M. Kumar, W.G. Hutchinson and S.M. Clarke, 2006. Ontogenetic development of digestive enzymes in yellowtail kingfish Seriola lalandi larvae. Aquaculture, 260: 264-271.
CrossRefDirect Link - Naz, M., 2009. Ontogeny of biochemical phases of fertilized eggs and yolk sac larvae of gilthead seabream (Sparus aurata L.). Turk. J. Fish. Aquat. Sci., 9: 77-83.
Direct Link - Bakhtiarvandi, N.K. and A.M. Abedian-Kenari, 2015. Changes of digestive enzymes activity in Caspian Kutum (Rutilus frisii Kutum) during larval developmental stages. Irian J. Fish. Sci., 14: 158-175.
Direct Link - Kamaci, H.O., D. Coban, C. Suzer, B. Aksu, S. Suka and K. Firat, 2010. Exocrine pancreas development and trypsin expression in cultured European Sea Bass (Dicentrarchus labrax) larvae. Turk. J. Fish. Aquat. Sci., 10: 123-130.
Direct Link - Srichanun, M., C. Tantikitti, V. Vatanakul and P. Musikarune, 2012. Digestive enzyme activity during ontogenetic development and effect of live feed in green catfish larvae (Mystus nemurus Cuv. & Val.). Songklanakarin J. Sci. Technol., 34: 247-254.
Direct Link - Farhoudi, A., A.M.A. Kenari, R.M. Nazari and C.H. Makhdoomi, 2013. Changes of digestive enzymes activity in common carp (Cyprinus carpio) during larval ontogeny. Iran. J. Fish. Sci., 12: 320-334.
Direct Link - Park, S.J., S.G. Lee and W.S. Gwak, 2015. Ontogenetic development of the digestive system in chub mackerel Scomber japonicas Larvae and Juveniles. Fish. Aquat. Sci., 18: 301-309.
CrossRefDirect Link - Buentello, J.A., C. Pohlenz, D. Margulies, V.P. Scholey and J.B. Wexler et al., 2011. A preliminary study of digestive enzyme activities and amino acid composition of early juvenile yellowfin tuna (Thunnus albacares). Aquaculture, 312: 205-211.
CrossRefDirect Link - Zavala-Leal, I., S. Dumas, R. Pena, M. Contreras-Olguin and D. Hernandez-Ceballos, 2013. Effects of culture conditions on feeding response of larval Pacific red snapper (Lutjanus peru, Nichols & Murphy) at first feeding. Aquac. Res., 44: 1399-1406.
CrossRefDirect Link - Kamarudin, M.S., S. Otoi and C.R. Saad, 2011. Changes in growth, survival and digestive enzyme activities of Asian redtail catfish, Mystus nemurus, larvae fed on different diets. Afr. J. Biotechnol., 10: 4484-4493.
Direct Link - Borlongan, I.G., 1990. Studies on the digestive lipases of milkfish, Chanos chanos. Aquaculture, 89: 315-325.
CrossRefDirect Link - Blumenthal, E., 2010. Introduction to SPSS 16.00. In: Center for Social Science Computation and Research. Blumenthal, E. (Eds.). University of Washington, Seattle, WA, USA., pp: 1-15.
Direct Link - Gisbert, E., G. Gimenez, I. Fernandez, Y. Kotzamanis and A. Estevez, 2009. Development of digestive enzymes in common dentex Dentex dentex during early ontogeny. Aquaculture., 287: 381-387.
CrossRefDirect Link - Cruz, E.M. and C.M. James, 1989. The effects of feeding rotifers (Brachionus plicatilis typicus) on the yield and growth of tilapia (Oreochromis spilurus) fry. Aquaculture, 77: 353-361.
CrossRefDirect Link - Bakhtiyar, Y., S. Langer, S.K. Karlopia and I. Ahmed, 2011. Growth, survival and proximate body composition of Labeo rohita larvae fed artificial food and natural food organisms under laboratory condition. Int. J. Fish. Aquacult., 3: 114-117.
Direct Link - Honji, R.M., C.E. Tolussi, P.H. Mello, D. Caneppele and R.G. Moreira, 2012. Embryonic development and larval stages of Steindachneridion parahybae (Siluriformes: Pimelodidae)-implications for the conservation and rearing of this endangered Neotropical species. Neotrop. Ichthy., 10: 313-327.
CrossRefDirect Link - Suzer, C., H.O. Kamaci, D. Coban, S. Saka, K. Firat, B. Ozkara and A. Ozkara, 2007. Digestive enzyme activity of the red porgy (Pagrus pagrus, L.) during larval development under culture conditions. Aquacult. Res., 38: 1178-1785.
CrossRefDirect Link - Kuz'mina, V.V., 1996. Influence of age on digestive enzyme activity in some freshwater teleosts. Aquaculture, 148: 25-37.
CrossRefDirect Link - Dbrowski, K., 1982. Proteolytic enzyme activity decline in starving fish alevins and larvae. Environ. Biol. Fish., 7: 73-76.
CrossRefDirect Link - Munilla-Moran, R., J.R. Stark and A. Barbour, 1990. The role of exogenous enzymes in digestion in cultured turbot larvae (Scophthalmus maximus L.). Aquaculture, 88: 337-350.
CrossRefDirect Link - Applebaum, S.L. and G.J. Holt, 2003. The digestive protease, chymotrypsin, as an indicator of nutritional condition in larval red drum (Sciaenops ocellatus). Mar. Biol., 142: 1159 -1167.
CrossRefDirect Link - Rathore, R.M., S. Kumar and R. Chakrabarti, 2005. Digestive enzyme patterns and evaluation of protease classes in Catla catla (Family: Cyprinidae) during early developmental stages. Comp. Biochem. Physiol. Part B: Biochem. Mol. Biol., 142: 98-106.
CrossRefPubMedDirect Link - Al-Tameemi, R., A. Aldubaikul and N.A. Salman, 2010. Comparative study of α-amylase activity in three Cyprinid species of different feeding habits from Southern Iraq. Turk. J. Fish. Aquat. Sci., 10: 411-414.
CrossRef - Ma, H.M., C. Cahu, J. Zambonino, H. Yu, Q.Y. Duan, M.M. Le Gall and K. Mai, 2005. Activities of selected digestive enzymes during larval development of large yellow croaker (Pseudosciaena crocea). Aquaculture, 245: 239-248.
CrossRefDirect Link - Sahlmann, C., J. Gu, T.M. Kortner, I. Lein, A. Krogdahl and A.M. Bakke, 2015. Ontogeny of the digestive system of atlantic salmon (Salmo salar L.) and effects of soybean meal from start-feeding. Plos One.
CrossRefDirect Link
Jayadi Reply
I am very satisfied