
Reza Zarghami
Agricultural Biotechnology Research Institute of Iran (ABRII), Shahid Fahmideh Blv. Karaj, 31535-1897, Iran
Ali Salari
National Research Centre on Plant Biotechnology, Indian Agricultural Research Institute, New Delhi, 110012, India
Pakistan Journal of Biological Sciences
Year: 2015 | Volume: 18 | Issue: 6 | Page No.: 260-266







ABSTRACT
Present study is set to develop a method to proliferate and rooting the Persian walnut (Juglans regia L.) in in vitro condition. To evaluate the proliferation of three genotypes, the DKW basal medium supplemented with four different concentrations of BAP (0, 0.5, 1 and 1.5 mg L–1) and IBA (0.01 mg L–1) conducted in three replications under factorial randomized block design were used. The root induction evaluated by using DKW medium supplemented by three different concentrations of IBA (0, 3 and 4 mg L–1) for one week. By study of growth factors, we conclude, the BAP (1.5 mg L–1) have shown appreciable results and the Chandler genotype were more responsive to the concentration compare to Hartley and Z60 genotypes. To root induction and callus initiation, IBA (3 mg L–1) were found to be responsive hormone with 7 days in dark incubation. The Chandler genotype compare to the other genotypes studied here, were more responsive to this hormonal level by showing high root percentage.
PDF Abstract XML References Citation
Received: August 03, 2015;
Accepted: September 23, 2015;
Published: October 14, 2015
How to cite this article
Reza Zarghami and Ali Salari, 2015. Effect of Different Hormonal Treatments on Proliferation and Rooting of Three Persian Walnut (Juglans regia L.) Genotypes. Pakistan Journal of Biological Sciences, 18: 260-266.
DOI: 10.3923/pjbs.2015.260.266
URL: https://scialert.net/abstract/?doi=pjbs.2015.260.266
DOI: 10.3923/pjbs.2015.260.266
URL: https://scialert.net/abstract/?doi=pjbs.2015.260.266
INTRODUCTION
Walnut belongs to Juglandaceae family and Juglans genus. The Persian walnut (Juglans regia L.), a dominant species of Juglandaceae family is originated from high hills of central Asia. Grafting is the most widely used way to propagate the walnut plants, globally. It can be propagated by seed and cutting but the seed originated offspring will often results variation and the other ways are slow and time consuming (Al-Mizory and Mayi, 2012). Tissue culture techniques are often use for fast plants proliferation. In vitro proliferation is an efficient alternative to the other methods, like grafting, which is fast and fulfill the genetic stability. Therefore, micro propagation soon became one of the biggest research fields to walnut propagation (Gruselle and Boxus, 1990; Silva and Dias, 1997; Navatel and Bourrain, 2001; Lopez, 2004; Breton et al., 2004; Vahdati et al., 2009).
Slow regeneration and rooting, phenolic compounds and endogenous bacterial contaminations are the main barriers to successful in vitro proliferation in Persian walnut protocol. Study on in vitro propagation of chandler genotypes showed early fruiting of these plants compare to the genotypes proliferated with conventional methods. In addition, they had well rooting system and no incompatibility between stock and scion were seen (Hasey et al., 2001; Lopez, 2001). Successful use of fresh wing tips, nodal and meristem as explant were reported (Leslie and McGranahan, 1998; Vahdati et al., 2009; Meynier, 1984; Heile-Sudholt et al., 1986). The culture contamination during in vitro culture should be consider. Seventy percent alcohol, sodium hypochlorite (2.5%), calcium hypochlorite and mercuric chloride (0.01%) for exogenous contaminations and antibiotics recommended to control the inner tissues adulterations (Sticklen and Oraby, 2005; Ainsley et al., 2001; Koubouris and Vasilakakis, 2006; Sharma et al., 1999). Also, nano silver particles suggested to control the bacterial contaminations (Nomiya et al., 2004; Sondi and Salopek-Sondi, 2004).
The toxic substances released from walnut explants are one of the other problem to explant establishment (Preece et al., 1989). Therefore, establishment phase in in vitro culture is very crucial to successful walnut micro propagation. Addition of antioxidants (ascorbic acid, citric acid, polyvinylpyrrolidone (PVP), dithiothreitol (DTT) and albumin) in to the culture medium, incubation of explants before culturing, low light or dark condition in initial days of culturing and repeated sub culturing are successful percussions to this stage (Leslie and McGranahan, 1998; Liu and Han, 1986; Heile et al., 1984; Navatel and Bourrain, 2001).
Proliferation and rooting are the main aims of in vitro protocols. The DKW medium known to reach to this target. Moreover, different concentrations of BAP, IBA and BA found to be stimulant to increase the walnut vegetative buds induction, propagation and rooting (Saadat and Hennerty, 2002; Driver and Kuniyuki, 1984; Penuela et al., 1988; Amiri, 2005; Jay-Allemand et al., 1992; Tetsumura et al., 2002; Sanchez-Zamora et al., 2006). In recent years, the walnut micro propagation methods are highly developed but successful shoot and root developments still in demand to commercialize the protocol. Therefore, present study is set to develop a method to proliferate and rooting the Persian walnut in in vitro condition.
MATERIALS AND METHODS
Explant preparation, sterilization and establishment: The newly grown shoots (40-60 cm) were collected from three high yielded genotypes (2-3 years old) (Chandler, Hartley and Z60) from the institute of horticultural research in Karaj, Iran in April 2014. The shoots were cut to the nodal segments.The explants were kept under three stages of sterilization treatment. Because of different explants sensitivity to the sterilization chemicals and their contamination levels, the soft stems (70% ethanol for 30 sec, 1.5% sodium hypochlorite combine with 2-3 drops of twin 20, 2.5% nano silver for 15 min, distilled water wash for 3 times, 5, 10 and 15 min) and the semi hard stems (70% ethanol for 40 sec, 1.5% sodium hypochlorite combine with 2-3 drops of twin 20 for 10-12 min, 2.5% nano silver for 20 min, distilled water wash for 3 times, 5, 10 and 15 min) surface sterilized separately. The explants dried up with filter paper before culturing. Driver-Kuniyuki-Walnut (DKW) medium supplemented with 2.1 g L–1 Phytagel (Sigma Chemical Co.), 30 g L–1 sucrose, 1 mg L–1 BAP and 0.01 mg L–1 IBA was used for establishment phase. To control the negative effects of phenols, the cultured explants incubated for 24 h at 4°C in dark. The explants were maintained in canning jars (0.3 l), at 25±2°C under 16 h photoperiod and light intensity of 40-60 μmol m‾2 sec–1 supplied by cool white fluorescent lamp.
Proliferation: The contaminated free in vitro samples which grown well in establishment phase were used. To evaluate the proliferation of three genotypes, the DKW basal medium supplemented with four different concentrations of BAP (0, 0.5, 1 and 1.5 mg L–1) and IBA (0.01 mg L–1) conducted in three replications under factorial randomized block design were used (Driver and Kuniyuki, 1984). All the cultures were sub cultured to the fresh medium of the same composition every four weeks. At the end of multiplication phase, shoot length, shoot number, number of leaves and callus size, were recorded (Fig. 1).
 | |
| Fig. 1(a-f): | In vitro propagation of Persian walnut (a, b) Explant preparation, (c) Establishment, (d, e) Proliferation and (f) Rooting |
Rooting: In root induction phase, regenerated shoots (3-5 cm), were excised and transferred onto rooting media. The root induction evaluated by using DKW medium supplemented by three different concentrations of IBA (0, 3 and 4 mg L–1) and were kept in darkness at 24±1°C for one week (Driver and Kuniyuki, 1984). In root development phase, the mix of hormone free, quarter-strength DKW: vermiculite to conduct soil aeration and root penetration were used. After 21 days, the root percentage, root length and number of roots conducted in three replication under factorial randomized block design were examined.
Statistical analysis: All statistical analysis of this experiment were analyzed using SAS software. The Duncan’s multiple test range were used for mean comparison and all graphs draws by Excel program.
RESULTS AND DISCUSSION
Effect of benzylaminopurine on proliferation: Analysis of variance shows the significant difference on different BAP levels (1%) and the interaction effects between BAP and genotypes on shoot length and callus size (5%) and shoot number (1%) were significant (Table 1). According to the mean comparison results (Table 2), there were significant difference among BAP levels on studied factors. In all factors, between control and other hormonal (BAP) treatments level were significant difference and by increasing the BAP levels, shoot length, shoot number and leaf number also increased. But, two BAP levels (0.5 and 1 mg L–1) did not show any significant difference while , the highest callus size were found in 1 mg L–1 BAP and decreased by 1.5 mg L–1.
Factors observations (shoot number and callus size) after 30 days of culturing showed the significant difference among genotypes and factors. The Chandler genotype showed more number of shoots (2.35) but two other genotypes, Hartley (1.33) and Z60 (1.48), did not show any significant difference in shoot number (Fig. 2). Genotype Z60 (2.91 mm) had lowest callus size and were significantly different with chandler (3.99 mm) and Hartley (4.15 mm) genotypes (Fig. 3). In similar study on two different walnut genotypes (pre mature dwarf and normal), the average number of shoots (3-3.3) were observed in dwarf plants (G4, G12 and G16), which compare to the normal genotypes (G8) (2.3) were high. Also, the lowest callus accumulation reported in dwarf, compare to normal genotypes which found closely similar to this study results (Vahdati et al., 2009).
The interaction between hormonal (BAP) treatments and walnut genotypes for the factors; shoot length, callus size and shoot number were significant at 5 and 1%, respectively. According to the results (Fig. 4), by increasing the hormonal level (BAP) a notable increase were observed in chandler genotype on shoot length (4.08 cm) and shoot number (4) in 1.5 mg L–1 BAP concentration, followed by genotype Z60 (3.42 cm) which were significantly different with Hartley genotype in shoot length. The best treatment in callus size (showed the lowest callus size) was control treatment in all genotypes followed by genotype Z60 (3.14 mm) at BAP (0.5 mg L–1) level.
The treatment effects (BAP) on genotypes had significant difference with control in all factors, which by increasing the BAP level up to 1.5 mg L–1, shoot length, shoot number and leaf number gradually increased, unlike callus size which this concentration had less effect compare to 1 mg L–1 on it. The interaction effect of BAP and genotypes on shoot length and callus size (1%) and shoot number (5%) were significant. Observations confirms that, the best proliferation hormonal (BAP) concentration was BAP (1.5 mg L–1) for chandler genotype with highest shoot number.
Induction of cell division, in vitro initiation and growth are the roll of cytokinins. Benzylaminopurine (BAP), zeatin, kinetin, 2ip and TDZ are the widely used cytokinins (Neuman et al., 1993; Makara et al., 2010). In this study, the treatment of explants with BAP hormone showed significant difference in studied factors.
| Table 1: | Analysis of variance for different benzylaminopurine levels and genotypes on proliferation and their interactions |
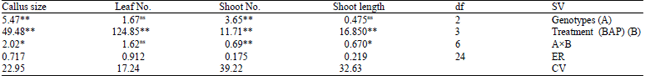 | |
*’**Significant at 5 and 1%, respectively, ns: Non-significant, BAP: Benzylaminopurine, df: Degrees of freedom | |
| Table 2: | Mean comparison of studied factors for proliferation under different benzylaminopurine levels |
 | |
| BAP: Benzylaminopurine | |
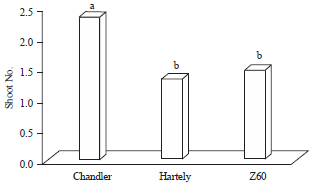 | |
| Fig. 2: | Mean comparison of shoot numbers on different genotypes |
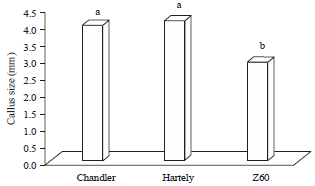 | |
| Fig. 3: | Mean comparison of callus size on different genotypes |
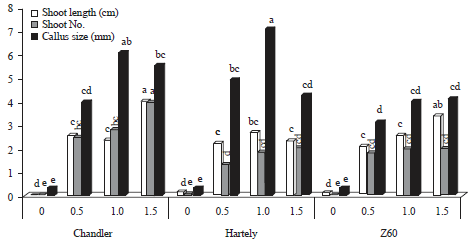 | |
| Fig. 4: | Mean comparison of BAP (0, 0.5, 1 and 1.5 mg L–1) and genotypes interaction on different factors |
By increasing the BAP concentration, shoot length, shoot number and leaf number were increased which were similar to previously studied results (Bosela and Michler, 2008; Gotea et al., 2012). Recently, the best results achieved by BAP combine with IBA and high concentrations of BA with IBA in walnut in vitro culture (Rios et al., 2005; Yari et al., 2014). These support the results of this study and show the importance of cytokinins in walnut proliferation.
Effect of indole butyric acid (IBA) on rooting: The walnut rooting potential is depend on endogenous levels of auxin and its external concentration and initiate by plantlets callus formation (Feito et al., 1997; Rios et al., 2005). Well grown plantlets selected to transfer to the rooting medium supplemented with hormone. After 21 days, root percentage, root length and root number were studied. The analysis of variance showed the significant difference (1%) among different IBA levels in all factors. The interaction effect between hormonal treatments (IBA) and genotypes were significant (1%) in root number. Also, in genotypes, as a studied factor, the root percentage and root number showed significant difference (1%) (Table 3). The mean comparison of IBA levels in different factors shows significant deference (Table 4). There were significant difference between control and treatments in all factors, which IBA (3 mg L–1) had highest root percentage and root length and an increase in root number.
| Table 3: | Analysis of variance of different indole butyric acid levels, genotypes and their interactions for rooting study |
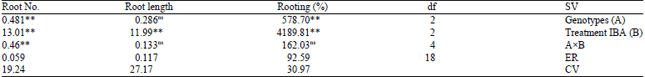 | |
*’**Significant at 5 and 1%, respectively, ns: non-significant, df: Degrees of freedom, IBA: Indole butyric acid | |
| Table 4: | Mean comparison of studied factors under different indole butyric acid levels on rooting |
 | |
| IBA: Indole butyric acid | |
 | |
| Fig. 5: | Mean comparison of rooting percentage on different genotypes |
But, by increasing the IBA level a decrease in those factors were observed.
Results showed, in callus percentage, the chandler genotype (33.33%) gained more average in this factor but there were no any difference between Z60 and Hartley genotype (19.44%) (Fig. 5). Chandler genotype also found more number of roots per explant (1.53) which were significantly different with Z60 (1.14) and Hartley (1.11). While, the difference between those last genotypes were not significant (Fig. 6). The interaction effect of IBA and walnut genotypes for root number factor were significant (1%). Hence, there were no significant difference among hartley (2.1) and Z60 (1.91) but the chandler genotype (3.16) in effect of IBA (3 mg L–1) were shown significant difference (Fig. 7).
According to the results of this study, we conclude that, the IBA (3 mg L–1) were best hormonal treatment for rooting of walnut micro-shoots which showed significant difference with rest of the treatments. The similar experiments showed the highest rooting rate treated with IBA compare to NAA. Also, the effect of genotype on rooting in different plant species were examined with several other experiments, which are compatible with present study.
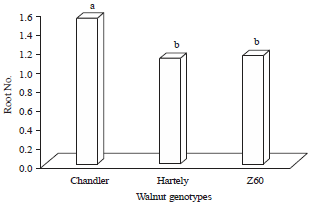 | |
| Fig. 6: | Mean comparison of root number on different genotypes |
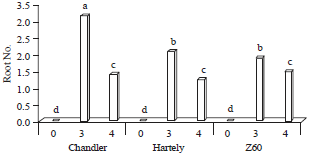 | |
| Fig. 7: | Mean comparison of root number on IBA (0, 3 and 4 mg L–1) and genotypes interaction |
This shows the need of different hormonal concentrations and rooting induction period for different genotypes and also shows the effect of endogenous hormonal level on rooting (Pruski et al., 2005; Vahdati et al., 2002; Saadat and Hennerty, 2002; Rugini et al., 1993). The effect of basal MS medium supplemented with IBA found to be more effective than IAA in some cases (Al-Mizory and Mayi, 2012; James et al., 1980).
By study of growth factors, we conclude, the BAP (1.5 mg L–1) have shown appreciable results and the chandler genotype were more responsive to that concentration compare to Hartley and Z60 genotypes. To root induction and callus initiation, IBA (3 mg L–1) were found to be responsive hormone with 7 days in dark incubation. The Chandler genotype compare to the other genotypes studied here, were more responsive to this hormonal level by showing high root percentage.
REFERENCES
- Ainsley, P.J., G.G. Collins and M. Sedgely, 2001. In vitro rooting of almond (Prunus dulcis Mill.). ln vitro Cell. Dev. Biology-Plant, 37: 778-785.
CrossRefDirect Link - Vahdati, K., R. Razaee and M. Mirmasoomi, 2009. Micropropagation of some dwarf and early mature walnut genotypes. Biotechnology, 8: 171-175.
CrossRefDirect Link - Al-Mizory, L.S.M. and A.A. Mayi, 2012. In vitro propagation of walnut (Juglans regia) by nodal explants. J. Agric. Sci. Technol. B, 2: 665-670.
Direct Link - Amiri, M.E., 2005. Effect of mineral concentration on tissue-cultured walnut (Juglans regia var. Zeiabadi) growth. Acta Horticulture, 705: 383-386.
CrossRefDirect Link - Bosela, M.J. and C.H. Michler, 2008. Media effects on black walnut (Juglans nigra L.) shoot culture growth in vitro: Evaluation of multiple nutrient formulations and cytokinin types. In vitro Cell. Dev. Biol. Plant, 44: 316-329.
CrossRefDirect Link - Breton, C., D. Cornu, D. Chriqui, A. Sauvanet, P. Capelli, E. Germain and C. Jay-Allemand, 2004. Somatic embryogenesis, micropropagation and plant regeneration of early mature walnut trees (Juglans regia) that flower in vitro. Tree Physiol., 24: 425-435.
CrossRefDirect Link - Feito, I., M.A. Gea, D. Rios, M. Albuerne, B. Fernandez and R. Rodriguez, 1997. Physiological studies during walnut rooting. Acta Hortic., 442: 137-142.
CrossRefDirect Link - Gotea, R., I. Gotea, R.E. Sestras and K. Vahdati, 2012. In vitro propagation of several walnut cultivars. Bull. UASVM Hortic., 69: 167-171.
Direct Link - Gruselle, R. and P. Boxus, 1990. Walnut micropropagation. Acta Horticulturae, 284: 45-52.
CrossRefDirect Link - Hasey, J.K., B.B. Westerdahl, W.C. Micke, D.E. Ramos and J.T. Yeager, 2001. Yield performance of own-rooted Chandler walnut versus Chandler walnut on paradox rootstock. Acta Horticulturae, 544: 489-493.
CrossRefDirect Link - Heile-Sudholt, C., C.A. Huetteman, J.E. Preece, J.W. van Sambeek and G.R. Gaffney, 1986. In vitro embryonic axis and seedling shoot tip culture of Juglans nigra L. Plant Cell Tissue Organ Culture, 6: 189-197.
CrossRefDirect Link - James, D.J., V.H. Knight and I.J. Thurbon, 1980. Micropropagation of red raspberry and the influence of phloroglucinol. Scientia Horticulturae, 12: 313-319.
CrossRefDirect Link - Jay-Allemand, C., P. Capelli and D. Cornu, 1992. Root development of in vitro hybrid walnut microcuttings in a vermiculite-containing gelrite medium. Scientia Horticulturae, 51: 335-342.
CrossRefDirect Link - Koubouris, G. and M. Vasilakakis, 2006. Improvement of in vitro propagation of apricot cultivar Bebecou. Plant Cell Tissue Orang Cult., 85: 173-180.
CrossRefDirect Link - Lopez, J.M., 2001. Field behaviour of self-rooted walnut trees of different cultivars produced by tissue culture and planted in Murcia (Spain). Acta Horticulturae, 544: 543-546.
CrossRefDirect Link - Makara, A.M., P.R. Rubaihayo and J.S. Magambo, 2010. Carry-over effect of Thidiazuron on banana in vitro proliferation at different culture cycles and light incubation conditions. Afr. J. Biotechnol., 21: 3079-3085.
Direct Link - Silva, D.J.M. and J.S.D. Dias, 1997. In vitro shoot culture on Juglans regia L.: Repeated subculturing on juvenile and adult material. Acta Hortic., 442: 251-256.
CrossRefDirect Link - Navatel, J.C. and L. Bourrain, 2001. Plant production of walnut (Juglans regia L.) by in vitro multiplication. Acta Hortic., 544: 465-471.
CrossRefDirect Link - Neuman, M.C., J.E. Preece, J.W. Van Sambeek and G.R. Gaffney, 1993. Somatic embryogenesis and callus production from cotyledon explants of Eastern black walnut. Plant Cell Tissue Org. Cult., 32: 9-18.
CrossRefDirect Link - Nomiya, K., A. Yoshizawa, K. Tsukagoshi, N.C. Kasug, S. Hirakawa and J. Watanabe, 2004. Synthesis and structural characterization of silver(I), aluminium(III) and cobalt(II) complexes with 4-isopropyltropolone (hinokitiol) showing noteworthy biological activities. Action of silver(I)-oxygen bonding complexes on the antimicrobial activities. J. Inorg. Biochem., 98: 46-60.
CrossRefDirect Link - Penuela, R., C. Garavito, R. Sanchez-Tames and R. Rodriguez, 1988. Multiple shoot-bud stimulation and rhizogenesis induction of embryogenic and juvenile explants of walnut. Acta Hortic. 227: 457-459.
CrossRefDirect Link - Pruski, K., T. Astatkie and J. Nowak, 2005. Tissue culture propagation of Mongolian cherry (Prunus fruticosa) and Nanking cherry (Prunus tomentosa). Plant Cell Tissue Orang Cult., 82: 207-211.
CrossRefDirect Link - Sanchez-Zamora, M.A., J. Cos-Terrer, D. Frutos-Tomas and R. Garcia-Lopez, 2006. Embryo germination and proliferation in vitro of Juglans regia L. Scientia Horticulturae, 108: 317-321.
CrossRefDirect Link - Saadat, Y.A. and M.J. Hennerty, 2002. Factors affecting the shoot multiplication of Persian walnut (Juglans regia L.). Sci. Hortic., 95: 251-260.
CrossRefDirect Link - Sharma, M., A. Sood, P.K. Nagar, O. Prakash and P.S. Ahuja, 1999. Direct rooting and hardening of tea microshoots in the field. Plant Cell Tissue Organ Cult., 58: 111-118.
CrossRefDirect Link - Sondi, I. and B. Salopek-Sondi, 2004. Silver nanoparticles as antimicrobial agent: A case study on E. coli as a model for Gram-negative bacteria. J. Colloid Interface Sci., 275: 177-182.
CrossRefDirect Link - Sticklen, M.B. and H.F. Oraby, 2005. Shoot apical meristem: A sustainable explant for genetic transformation of cereal crops. In vitro Cell. Dev. Biology-Plant, 41: 187-200.
CrossRefDirect Link - Tetsumura, T., K. Tsukuda and K. Kawase, 2002. Micropropagation of shinano walnut (Juglans regia L.). J. Jpn. Soc. Hortic. Sci., 17: 661-666.
CrossRef - Yari, M.B., M. Gholami and I. Khazaei, 2014. Impact of media and different cytokinins concentrations on in vitro shoot multiplication of persian walnut (Juglans regia L.). Int. J. Farm. Allied Sci., 3: 203-209.
Direct Link - Vahdati, K., J.R. McKenna, A.M. Dandekar, C.A. Leslie and S.L. Uratsu et al., 2002. Rooting and other characteristics of a transgenic walnut hybrid (Juglans hindsii x J. regia) rootstock expressing rolABC. J. Am. Soc. Hortic. Sci., 127: 724-728.
Direct Link - Rugini, E., A. Jacoboni and M. Luppino, 1993. Role of basal shoot darkening and exogenous putrescine treatments on in vitro rooting and on endogenous polyamine changes in difficult-to-root woody species. Scientia Horticulturae, 53: 63-72.
CrossRefDirect Link