
Somayeh Hajipour
Department of Biology, Faculty of Sciences, Isfehan Payamenoor University, Isfehan, Iran
Alireza Sarkaki
Physiology Research Center, Ahvaz Jundishapur University of Medical Sciences, Ahvaz, Iran
Seyed Mohammad Taghi Mansouri
Department of Pharmacology, Faculty of Medicine, Ahvaz Jundishapur University of Medical Sciences, Ahvaz, Iran
Asghar Pilevarian
Department of Biology, Faculty of Sciences, Isfehan Payamenoor University, Isfehan, Iran
Maryam RafieiRad
Department of Biology, Faculty of Sciences, Islamic Azad University, Izeh Branch, Izeh, Iran
Pakistan Journal of Biological Sciences
Year: 2014 | Volume: 17 | Issue: 8 | Page No.: 991-998







ABSTRACT
This study aimed to evaluate the effect of two weeks oral administration of Pomegranate Seed Extract (PGSE) on active avoidance memory and motor coordination activities after permanent bilateral common carotid arteries occlusion (2CCAO) in male adult rats. Adult male albino rats of Wistar strain (250±20 g, 3-4 months) were used. Animals were divided into eight groups with 10 in each: (1) Sham operated (Sh); (2) Ischemic (I); (3) Ischemic received 100 mg kg-1 PGSE, orally (I+E100); (4) Ischemic received 200 mg kg-1 PGSE, orally (I+E200); (5) Ischemic received 400 mg kg-1 PGSE, orally (I+E400); (6) Ischemic received 800 mg kg-1 PGSE, orally (I+ E800); (7) Ischemic received 2 mL kg-1 normal saline, orally (I+Veh); (8) Sham operated received 400 mg kg-1 PGSE, orally (Sh+E400). In order to make 2CCAO an animal Cerebral Hypoperfusion Ischemia (CHI) model, carotid arteries were ligatured and then bilaterally cut. To evaluate active avoidance task, Correct Response Percentages (CRP) was measured by Y-maze apparatus and motor coordination activity was evaluated using standard behavioral tests by rotarod apparatus in all the rats. It was found that memory. Memory and motor coordination activities were significantly impaired in the rats after CHI (p<0.01). PGSE treatment significantly improved impairment of memory and motor coordination in the rats with 2CCAO (p<0.001). PGSE exhibited therapeutic potential for memory and muscular coordination, which was most likely related at least in some part to its antioxidative and free radical scavenging actions.
PDF Abstract XML References Citation
Received: December 14, 2013;
Accepted: February 13, 2014;
Published: March 29, 2014
How to cite this article
Somayeh Hajipour, Alireza Sarkaki, Seyed Mohammad Taghi Mansouri, Asghar Pilevarian and Maryam RafieiRad, 2014. Motor and Cognitive Deficits due to Permanent Cerebral Hypoperfusion/ischemia Improve by Pomegranate Seed Extract in Rats. Pakistan Journal of Biological Sciences, 17: 991-998.
DOI: 10.3923/pjbs.2014.991.998
URL: https://scialert.net/abstract/?doi=pjbs.2014.991.998
DOI: 10.3923/pjbs.2014.991.998
URL: https://scialert.net/abstract/?doi=pjbs.2014.991.998
INTRODUCTION
Cerebral ischemia is induced by reduced cerebral blood flow due to transient or permanent occlusion of cerebral arteries (Dirnagl et al., 1999). Transient cerebral ischemia is known to result in neuronal death in some specific vulnerable regions such as hippocampus, neocortex and striatum (Kirino, 2000). Tissue damage following cerebral ischemia is caused by the interaction of complex pathophysiological processes such as excitotoxicity, depolarization, inflammation and apoptosis (Dirnagl et al., 1999). Neurons sensitive to ischemia include, neocortical neurons, striatal neurons, cerebellar purkinje cells and, most importantly, neurons in the CA1 zone of hippocampus, an area of brain thought to be critical for learning and memory (Brierley and Cooper, 1962).
Forebrain ischemia is caused by deficiency in blood supply to hippocampal neurons, which leads to disturbances in energy metabolism that leads to decreased glucose utilization with a consequent decrease in the production of ATP and phosphocreatine (Sims and Zaidan, 1995). Most of these changes are associated with massive production of Reactive Oxygen Species (ROS), which causes severe oxidative injury to the brain tissue (Chan, 2001). ROS are generated within brain tissue during cerebral hypoperfusion ischemia (CHI) and play a role in the development of cerebral damage (Pellegrini-Giampietro et al., 1989). Under normal conditions, ROS and antioxidant systems are in balance. Any imbalance between pro-oxidant and antioxidant factors, is called oxidative stress (Floyd and Hensley, 2002). Brain is believed to be particularly vulnerable to oxidative stress due to a relatively high rate of oxygen-free radical generation without commensurate levels of antioxidative defenses (Brewer, 1998). Earlier studies have reported that oxidative damage is a primary factor in various models of acute brain damage including forebrain ischemia (Al-Majed et al., 2006). Moreover, increase of oxidative stress biomarkers and depletion of enzymatic and non-enzymatic antioxidants have been reported in forebrain ischemia-induced neuronal damage (Gupta and Briyal, 2004). In ischemia, experimental evidence obtained from animals has causally linked selective necrosis of hippocampal Cornu Ammonis area 1 (CA1) pyramidal cells to an ischemia-induced release of the excitatory amino acid glutamate (Rothman and Olney, 1986). Glutamate may cause ischemic neuronal death by acting at excit atory N-methyl-D-Aspartate (NMDA) receptors (Simon et al., 1984) which play an important physiological role in long-term potentiation and memory (Wigstrom et al., 1986). Thus, high concentration of NMDA excitatory receptors on the endritic trees of hippocampal CA1 pyramidal cells is induced (Greenamyre et al., 1985).
Studies have shown potential neuroprotective effects of plant origin antioxidants such as flavonoid compounds against alcohol-induced injury (Antonio and Druse, 2008) and their beneficial effects on cognition have been demonstrated in animal studies (Farshchi et al., 2010; Taati et al., 2011) . Citrus fruits and juices are an important source of bioactive compounds including antioxidants such as ascorbic acid, flavonoids, phenolic compounds and pectins that are important in human nutrition (Fernandez-Lopez et al., 2005; Jayaprakasha and Patil, 2007). So, in this work, a well-characterized model of term CHI was used in adult male rats to investigate neuroprotective capability of pomegranate seed hydroalcoholic extract in a treatment paradigm.
MATERIALS AND METHODS
Animals: Eighty adult male albino rats of Wistar strain (250±20 g, 3-4 months) obtained from laboratory animal center of Ahvaz Jundishapur University of Medical Sciences (AJUMS) were used in this study. The animals were housed in standard cages at controlled room temperature 20±2°C, 55-60% humidity and light exposure conditions of 12:12 h light-dark cycle (lighted at 07:00 am). All experiments were carried out during light phase of the cycle (8:00 am to 6:00 pm). Access to food and water was ad libitum except during the experiments. Animal handling and experimental procedures were performed according the university and institutional legislation, controlled by local ethics committee for the purpose of control and supervision of experiments on laboratory animals. All efforts were made to minimize animal suffering by reduce the number of used animals. Prior to the onset of behavioral testing, all the rats were gently handled for 5 days (5 min every day) in order to adaptation with experiments condition. They were randomly divided into eight groups with 10 in each: (1) Sham operated (Sh) with manipulation of both common carotids arteries without occlusion, (2) Untreated ischemic (I) with occlusion of bilateral common carotids arteries, (3) Ischemic rats receiving pomegranate seed extract (PGSE 100 mg kg-1, orally) for 14 days (I+E100 ), (4) Ischemic rats receiving PGSE (200 mg kg-1, orally) for 14 days (I+E200), (5) Ischemic rats receiving PGSE (400 mg kg-1, orally) for 14 days (I+E400), (6) Ischemic rats receiving PGSE (800 mg kg-1, orally) for 14 days (I+E800), (7) Sham-treated animals (I+Veh), ischemic rats receiving the same volume of normal saline as PGSE vehicle (2 mL kg-1, orally) for 14 days and (8) Positive control group (Sh+E400), healthy rats receiving the best effective dose of PGSE (400 mg kg-1, orally) for 14 days.
Two common carotids arteries occlusion (2CCAO or 2VO) procedure: Cechetti et al. (2010) with little modification was used. Briefly, the rats were anesthetized using 50 mg kg-1 of Ketamine-HCl and 5 mg kg-1 Xylazine-HCl, intraperitoneal (IP) injection. A neck ventral midline incision was made; the common carotid arteries were then exposed and gently separated from the vagus nerve. The carotids were occluded with a one week interval between the interventions, the right common carotid was the first one to be assessed and the left one was occluded 1 week later. Sham-operated controls were under the same surgical procedures without carotid artery ligation and occlusion (Cechetti et al., 2010; Sarkaki et al., 2012).
PGSE preparation: Pomegranate fruits (Punica granatum L.) as a large fruit with red barriers were purchased from Shahreza granatum gardens, Isfahan province, Iran. The seeds removed from the fruits, air-dried in shade for one week and milled to fine powder (electric mill, Panasonic Co. Japan). Powdered seeds were macerated in 70% ethanol for 72 h at room temperature (1000 g 3 L-1). Ethanol extract was evaporated (Rotary Ovaporator, Heidolph Co. Germany) to remove ethanol and PGSE was obtained as a lyophilized powder (17±2% yield).
Treatment: Different doses of the extract were administrated to each animal in separate groups via forced oral administration (i.g.) everyday at 8:00-9:00 AM for 2 weeks, starting 5 days after ischemic injury. The sham-treated animals (I+Veh) received the same volume of normal saline for the same period.
Active avoidance task: The apparatus used for evaluating active avoidance memory, was the 3-equal arms Y-maze (made in Ahvaz, Iran). Besnard et al. (2012) with a little modification were used for evaluating active avoidance memory. Sham-operated, ischemic and all the treated rats were trained in a Y-maze. Training involved 30 trials every day for 4 consecutive days (sessions). The animals were conditioned using a 12 W light as the Conditioned Stimulus (CS) and 20-25 V, 3 mA electrical foot shock as the Unconditioned Stimulus (UCS). Inter-trial Interval (ITI) and inter-stimulus Interval (ISI) were 60 and 5 sec, respectively. The trained animals left the dark arms and entered the light arm before exposing to UCS as correct response. If this occurred within the 5 sec ISI, the effort was counted as a conditioned response. The Criterion Condition Response (CCR) was 90% (27 correct responses of 30 trials per training session).
Rotarod: For motor-coordination ability, 28 days after the CHI induction, the rats of all groups were evaluated on rotarod apparatus. The rotarod test served the purpose of detecting potential deleterious effects of the studied compounds on the rats's motor performance and coordination. The rotarod unit (Borj Sanaat Co., Tehran, Iran) consisted of a rotating rod, 75 mm in diameter, on which the rats were allowed to retain. The remaining time of each rat on the rotating bar was recorded. The maximum time was 15 min per trial. This apparatus automatically recorded the time in 0.1 sec and stopped counting when the rat fell back the rotating shaft. The animals were pre-trained to reach a stable performance in this test. During a familiar session, the animals were placed on a rod with constant 5 rpm for 3 min. The following day during the test session, the animals were placed on a rod with an initial constant rod speed of 5 rotations per minute (5 rpm) for 3 min; afterward, speed was programmatically increased to 40 rpm (5-10 rpm/next 3 min, 10-20 rpm/next 3 min, 20-30 rpm/next 3 min and 30-40 rpm/end 3 min) while the cut off time was 15 min. The test session consisted of three trials during one day. Inter-trial interval was 45 min. Data were presented as retention time (sec) on the rotating bar over the three test trials (Dekundy et al., 2006; Sameri et al., 2011).
Statistics: The data were expressed as Mean±S.E.M. of values for memory and motor coordination tests. Statistical analysis was performed by one-way ANOVA followed by LSD post-hoc test. p-value less than 0.05 assumed to denote a significant difference.
RESULTS
Active avoidance memory: Sham group animals were trained 4 sessions with 30 trials in each. They received to 90% criterion conditioned responses at 4th day of training (at least 27 corrected responses of 30 trials.
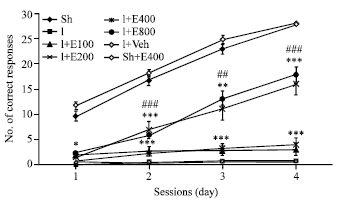 | |
| Fig. 1: | Mean±SEM of corrected conditioned responses (CCR) during 4 training sessions (days) with active avoidance test showed in sham operated (Sh), ischemic (I), ischemic rats treated with different doses of PGSE (100, 200, 400, 800 mg kg-1 and 2 mL kg-1 normal saline, orally) and sham operated groups receiving the best effective dose of PGSE (400 mg kg-1 2 mL-1). Data were analyzed by Repeated Measures (RM) ANOVA followed by LSD post-hoc test (n = 10). *: Significant difference with Sh group and #: Significant difference with I group, * or # p<0.05, ** or ## p<0.01 and *** or ###p<0.001 |
Figure 1 showed that number of Corrected Conditioned Responses (CCR) in Y-maze test during four training sessions decreased significantly (p<0.001) in I group two weeks after 2CCAO when compared with Sh group.
Treatment of 2CCAO rats with different doses PGSE for 14 days (100, 200, 400 and 800 mg kg-1, orally) improved significantly CCR when compared with I and I+Veh groups (p<0.05, p<0.001). But the effect of doses 400 and 800 mg kg-1 PGSE on CCR was same and also more potent than doses 100 and 200 of PGSE. So, with consideration this results in the sight of our dose 400 mg kg-1 of PGSE was an effective dose in this experiment. On the other hand, CCR of ischemic animals did not change after treatment with the same volume of PGSE vehicle (normal saline). Since the doses 400 and 800 mg kg-1 of PGSE had the same effect. Thus this dose, i.e., 400 mg kg-1 was administrated to sham operated rats as the positive control group; but PGSE was not significantly affected CCR in these rats (Fig. 1).
Motor coordination: The data obtained from all groups following motor balance test in rotarod system showed that bar descent latency in I severely decreased when compared with Sh (***p<0.001). Data of motor balance test in rotarod system showed that bar descending (s) in I group severely decreased when compared with Sh group (***p<0.001, Fig. 2).
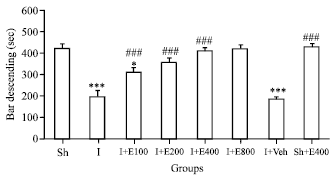 | |
| Fig. 2: | Mean±SEM of bar descending as motor coordination on rotarod in sham operated (Sh), ischemic (I) and rats suffering from cerebral hypoperfusion/ischemia which were treated with different doses of PGSE or Vehicle for 14 days (I+E100-800 , I+Veh) and (Sh+E 400 mg kg-1/2 mL) groups. Motor coordination in (I), (I+Veh) and (I+E100) were significantly weaker than Sh group (p<0.001 and p<0.05, n = 10) |
Treatment of 2CCAO rats with different doses PGSE for 14 days on 14th day of lesion (100, 200, 400 and 800 mg kg-1, orally) improved significantly motor coordination as bar descend latency (s) when compared with I and I+Veh groups (p<0.001). But the effect of doses 400 and 800 mg kg-1 PGSE on motor activity was same and also more potent than doses 100 and 200 of PGSE (Fig. 2). So, with consideration this result, in our sight the dose 400 mg kg-1 of PGSE was a best effective dose in this experiment. On the other hand, motor coordination of ischemic animals did not improve after treatment with the same volume of PGSE vehicle (normal saline). Since the doses 400 and 800 mg kg-1 of PGSE had the same effect on motor activity. Thus this dose, i.e., 400 mg kg-1 was administrated to healthy intact rats as the positive control group; but, PGSE was not significantly affected motor activity in related to untreated sham operated rats (Fig. 2).
DISCUSSION
According to the present findings, cognitive and motor activities were impaired in CHI rats. It was found that 14 days oral administration of different doses of PGSE could improve active avoidance learning and also motor activity in ischemic groups. However, in the related literature, few specific references (Sarkaki et al., 2012, 2013) were found to effects of PGSE on brain damages due to CHI or degeneration. So, based on results of this work, it could be suggested that PGSE with some compounds beside other useful contents such as ellagic acid, phytoestrogens and punicic acid, could act as an antioxidant for scavenge of free radicals from brain regions that influence cognition, especially hippocampus, striatum and cortex after CHI.
Brain ischaemia causes neuronal death in CA1 and CA3 regions of hippocampus (Hu and Wieloch, 1994). ROS is one of the most important factors that induce neuron death in CHI insult (Saeed et al., 2007). Behavioral studies on animals have demonstrated that hippocampus damage can produce learning and memory impairments (Zola-Morgan and Squire, 1986). Association of memory deficits with brain ischemia due to cardiac arrest or hypotension and the subsequent demonstration of hippocampal damage in the brain suggest that memory deficits are indeed due specifically to hippocampal damage (Sarkaki et al., 2013). Functional disturbance of hippocampus following mild ischemia is also reflected by behavioral changes. In agreement with earlier observations made in rats, hippocampal damage was associated with an impairment of learning behavior (Volpe et al., 1984). Brain is more vulnerable to oxidative stress than other organs due to its low antioxidant protection system and increased exposure of target molecules to reactive oxygen species. Nervous tissue has a high content of polyunsaturated fatty acids (Sun et al., 2002), which are easy targets for oxidative damage by free radicals due to their containing unsaturated bonds (Reiter, 1995). On the other hand, it has been revealed that brain structures supporting memory are uniquely sensitive to oxidative stress due to their elevated demand for oxygen (Floyd, 1999). Cells normally have a number of mechanisms to resist the damage induced by free radicals.
The major antioxidant defenses consist of antioxidant scavenger such as glutathione (GSH), vitamin C (ascorbic acid), vitamin E (alpha-tocopherol), carotenoids, flavonoids, polyphenols and antioxidant enzymes (Saeed et al., 2007).
Pomegranate contains some species of flavonoids and anthocyanidins in its seed oil and juice and shows antioxidant activity three times greater than green tea extract. Pomegrante juice contains tannins, ellagic tannis, anthocyanins, catechins, gallic and ellagic acid as antioxidant chemicals (Sarkaki et al., 2013). Gallic acid may directly combine with free radicals and inactivate them which may suppress intracellular concentration of free radicals (Volpe et al., 1984). Additionally, animal studies with pomegranate juice have provided further evidence that dietary flavonoids are beneficial in reversing the course of neuronal and behavioural aging (Sarkaki et al., 2013).
One mechanism underlying the neuroprotective effect of polyphenols is possibly through its effects on reducing levels of apoptotic markers. Pomegranate polyphenols and resveratrol protect neonatal mouse brain from ischemic injury by reducing caspase 3 and calpain activation (West et al., 2007). In neonatal rats, amentoflavone blocked the activation of caspase-3 and the proteolytic cleavage of its substrates following hypoxic-ischemic injury (Shin et al., 2006). Pomegranate juice also diminished caspase-3 activation in hippocampus and cortex of neonatal brain against a hypoxic-ischemic insult through supplementing maternal diet with pomegranate juice (Loren et al., 2005).
The second aim of the present study was to determine possible beneficial effects of PGSE on motor activity impairment after permanent CHI in rats. The rotarod test is a well-established procedure for testing balance and coordination aspects of motor performance in rats and mice (Jones and Roberts, 1968). Recent evidence has indicated that accelerating rotarod task is a more sensitive index for assessment of motor impairment induced by traumatic brain injury in rats than both beam-walking and beam-balancing tasks (Chang et al., 2013). Deficits in motor performance on the rotarod task had been observed from 24 h to 2 months after induction of focal cerebral ischemia in rats (Borlongan et al., 1995; Okada et al., 1995; Viswanatha et al., 2013). Recent evidence has demonstrated a marked impairment in skilled paw use after focal ischemia (Hamm et al., 1994; Borlongan et al., 1995; Tregub et al., 2013).
Decrements in motor function and decrements in memory are two main behavioral parameters that are altered in senescence in both humans and animals. Primary degenerative brain disease and diseases related to cerebral vascular disturbances are leading causes of disability at old age and can cause loss of autonomy, dependence and high social costs for people and society. There is growing evidence that free radicals are involved in the initiation of cellular injury observed in neurodegenerative diseases (Emerit et al., 2004; Pandi-Perumal et al., 2013). Experimental evidence has supported involvement of free radicals in CHI pathogenesis (Okada et al., 1995; Huy et al., 2013).
Biological properties of flavonoids are determined by the extent nature and position of substituents and the number of hydroxyl groups (Schroeter et al., 2000). Reactive nitrogen and oxygen species play important physiologic and pathologic roles in neonatal CHI and synthesis attenuation of these compounds during CHI has been shown to provide neuroprotection (Peeters-Scholte et al., 2002).
Accordingly, it is interesting to note that pomegranate juice has a significant antioxidant capacity (Youdim et al., 2002). Glutathione is a well characterized reactive nitrogen and oxygen species scavenger in brain and catechins (another constituent of pomegranate juice) have been shown to increase intracellular glutathione levels and modulate glutathione peroxidase activity (Savaskan et al., 2003; Chopra et al., 2004).
Although these compounds possess potent antioxidant capacities, they also increase superoxide dismutase activity and have been shown to protect brain's lipid peroxidation (Sun et al., 1999; Youdim et al., 2002 ). Efficient cerebral blood flow is also vital for optimal brain function and several studies have indicated a decrease in cerebral blood flow (CBF) among patients with dementia (Nagahama et al., 2003; Ruitenberg et al., 2005). Flavonoids have been shown to exert a positive effect on cerebral blood flow (CBF) in humans (Jones and Roberts, 1968; Pachauri et al., 2012).
CONCLUSION
Oxidative stress is a key feature for cerebral ischemia that affects neuronal viability after ischemia. There is tremendous potential for using polyphenols in preventing cell loss or damage and edema in cerebral ischemic injury. However, cellular and molecular actions of polyphenols involved in neuroprotection have to be further elucidated. PGSE improves memory and motor deficiency due to CHI in rats because its contents, such as vitamins, phytoestrogens, ellagic acid, punicic acid, polyphenols and flavonoids, act as antioxidant and remove free radicals in damaged brain tissue after CHI. Overall, exact mechanisms for effect of PGSE on cognition and motor activity needs more investigations.
ACKNOWLEDGMENTS
This study was extracted from Somaye Hajipour's M.Sc. thesis, Isfahan Payamenoor University. She was a guest student in Ahvaz Jundishapur University of Medical Sciences (AJUMS) during her thesis research work. This study was done in Physiology Research Center, Neurosciences Lab. (PRC-82).
REFERENCES
- Al-Majed, A.A., F.A. Al-Omar and M.N. Nagi, 2006. Neuroprotective effects of thymoquinone against transient forebrain ischemia in the rat hippocampus. Eur. J. Pharmacol., 543: 40-47.
CrossRefPubMedDirect Link - Antonio, A.M. and M.J. Druse, 2008. Antioxidants prevent ethanol-associated apoptosis in fetal rhombencephalic neurons. Brain Res., 1204: 16-23.
CrossRefPubMedDirect Link - Borlongan, C.V., D.W. Cahill and P.R. Sanberg, 1995. Locomotor and passive avoidance deficits following occlusion of the middle cerebral artery. Physiol. Behav., 58: 909-917.
CrossRefPubMedDirect Link - Brewer, G.J., 1998. Age-related toxicity to lactate, glutamate and β-amyloid in cultured adult neurons. Neurobiol. Aging, 19: 561-568.
CrossRefPubMedDirect Link - Brierley, J.B. and J.E. Cooper, 1962. Cerebral complications of hypotensive anaesthesia in a healthy adult. J. Neurol. Neurosurg. Psychiatry., 25: 24-30.
PubMedDirect Link - Cechetti, F., P.V. Worm, L.O. Pereira, I.R. Siqueira and C.A. Netto, 2010. The modified 2VO ischemia protocol causes cognitive impairment similar to that induced by the standard method, but with a better survival rate. Brazil. J. Med. Biol. Res., 43: 1178-1183.
CrossRefDirect Link - Chan, P.H., 2001. Reactive oxygen radicals in signaling and damage in the ischemic brain. J. Cerebral Blood Flow Metab., 21: 2-14.
CrossRef - Chopra, K., D. Singh and V. Chander, 2004. Nephrotoxicity and its prevention by catechin in ferric nitrilotriacetate promoted oxidative stress in rats. Hum. Exp. Toxicol., 23: 137-143.
CrossRefPubMedDirect Link - Dekundy, A., M. Pietraszek, D. Schaefer, M.A. Cenci and W. Danysz, 2006. Effects of group I metabotropic glutamate receptors blockade in experimental models of parkinson's disease. Brain Res. Bull., 69: 318-326.
PubMed - Dirnagl, U., C. Iadecola and M.A. Moskowitz, 1999. Pathobiology of ischaemic stroke: An integrated view. Trends Neurosci., 22: 391-397.
CrossRefPubMedDirect Link - Farshchi, A., G. Ghiasi, S. Farshchi and P.M. Khatabi, 2010. Effects of boswellia papyrifera gum extract on learning and memory in mice and rats. Iran. J. Basic Med. Sci., 13: 9-15.
Direct Link - Fernandez-Lopez, J., N. Zhi, L. Aleson-Carbonell, J.A. Perez-Alvarez and V. Kuri, 2005. Antioxidant and antibacterial activities of natural extracts: Application in beef meatballs. Meat Sci., 69: 371-380.
CrossRefDirect Link - Floyd, R.A., 1999. Antioxidants, oxidative stress and degenerative neurological disorders. Proc. Soc. Exp. Biol. Med., 222: 236-245.
PubMed - Floyd, R.A. and K. Hensley, 2002. Oxidative stress in brain aging: Implications for therapeutics of neurodegenerative diseases. Neurobiol. Aging, 23: 795-807.
CrossRefPubMedDirect Link - Greenamyre, J.T., J.M. Olson, J.B. Penney and A.B. Young, 1985. Autoradiographic characterization of N-methyl-D-aspartate-, quisqualate- and kainate-sensitive glutamate binding sites. J. Pharmacol. Exp. Ther., 233: 254-263.
PubMed - Gupta, Y.K. and S. Briyal, 2004. Animal models of cerebral ischemia for evaluation of drugs. Indian J. Physiol. Pharmacol., 48: 379-394.
PubMed - Hu, B.R. and T. Wieloch, 1994. Tyrosine phosphorylation and activation of mitogen-activated protein kinase in the rat brain following transient cerebral ischemia. J. Neurochem., 62: 1357-1367.
CrossRefDirect Link - Jayaprakasha, G.K. and B.S. Patil, 2007. In vitro evaluation of the antioxidant activities in fruit extracts from citron and blood orange. Food Chem., 101: 410-418.
CrossRef - Jones, B.J. and D.J. Roberts, 1968. The quantiative measurement of motor inco-ordination in naive mice using an accelerating rotarod. J. Pharm. Pharmaco., l20: 302-304.
CrossRefPubMedDirect Link - Loren, D.J., N.P. Seeram, R.N. Schulman and D.M. Holtzman, 2005. Maternal dietary supplementation with pomegranate juice is neuroprotective in an animal model of neonatal hypoxic-ischemic brain injury. Pediatr. Res., 57: 858-864.
CrossRefPubMedDirect Link - Nagahama, Y., H. Nabatame, T. Okina, H. Yamauchi and M. Narita et al., 2003. Cerebral correlates of the progression rate of the cognitive decline in probable Alzheimer's disease. Eur. Neurol., 50: 1-9.
CrossRefPubMedDirect Link - Peeters-Scholte, C., J. Koster, W. Veldhuis, E. van den Tweel and C. Zhu et al., 2002. Neuroprotection by selective nitric oxide synthase inhibition at 24 hours after perinatal hypoxia-ischemia. Stroke, 33: 2304-2309.
CrossRefPubMedDirect Link - Pellegrini-Giampietro, D.E., G. Cherici, M. Alesiani, V. Carla and F. Moroni, 1988. Excitatory amino acid release from rat hippocampal slices as a consequence of free-radical formation. J. Neurochem., 51: 1960-1963.
CrossRef - Reiter, R.J., 1995. Oxidative processes and antioxidative defense mechanisms in the aging brain. FASEB J., 9: 526-533.
PubMed - Rothman, S.M. and J.W. Olney, 1986. Glutamate and the pathophysiology of hypoxic-ischemic brain damage. Ann. Neurol., 19: 105-111.
CrossRef - Saeed, S.A., K.F. Shad, T. Saleem, F. Javed and M.U. Khan, 2007. Some new prospects in the understanding of the molecular basis of the pathogenesis of stroke. Exp. Brain Res., 182: 1-10.
PubMed - Sameri, M.J., A. Sarkaki, Y. Farbood and S.M.T. Mansouri, 2011. Motor disorders and impaired electrical power of pallidal EEG improved by gallic acid in animal model of Parkinson's disease. Pak. J. Biol. Sci., 14: 1109-1116.
CrossRefPubMedDirect Link - Sarkaki, A., S. Hajipour, T.M. Mansouri, A. Pilevarian and M.R. Rad, 2013. Pomegranate seed hydroalcoholic extract improves memory deficit due to permanent cerebral hypoperfusion/ischemia in male rats. HealthMed, 7: 863-871.
Direct Link - Sarkaki, A., M. Rafieirad, S.E. Hossini, Y. Farbood, S.M.T. Mansouri and F. Motamedi, 2012. Cognitive deficiency induced by cerebral hypoperfusion/ischemia improves by exercise and grape seed extract. Health Med. J., 6: 1097-1104.
Direct Link - Shin, D.H., Y.C. Bae, J.S. Kim-Han, J.H. Lee, I.Y. Choi, K.H. Son, S.S. Kang, W.K. Kim and B.H. Han, 2006. Polyphenol amentoflavone affords neuroprotection against neonatal hypoxic-ischemic brain damage via multiple mechanisms. J. Neurochem., 96: 561-572.
CrossRefPubMedDirect Link - Sims, N.R. and E. Zaidan, 1995. Biochemical changes associated with selective neuronal death following short-term cerebral ischaemia. Int. J. Biochem. Cell Biol., 27: 531-550.
CrossRefPubMedDirect Link - Sun, A.Y., A. Simonyi and G.Y. Sun, 2002. The French Paradox and beyond: Neuroprotective effects of polyphenols. Free Radical Biol. Med., 32: 314-318.
CrossRefPubMedDirect Link - Taati, M., M. Alirezaei, M.H. Moshkatalsadat, B. Rasoulian, M. Moghadasi, F. Sheikhzadeh and A. Sokhtezari, 2011. Protective effects of Ziziphus jujuba fruit extract against ethanol-induced hippocampal oxidative stress and spatial memory impairment in rats. J. Med. Plants Res., 5: 915-921.
Direct Link - Volpe, B.T., W.A. Pulsinelli, J. Tribuna and H.P. Davis, 1984. Behavioral performance of rats following transient forebrain ischemia. Stroke, 15: 558-565.
CrossRefPubMedDirect Link - West, T., M. Atzeva and D.M. Holtzman, 2007. Pomegranate polyphenols and resveratrol protect the neonatal brain against hypoxic-ischemic injury. Dev. Neurosci., 29: 363-372.
CrossRefPubMedDirect Link - Wigstrom, H., B. Gustafsson and Y.Y. Huang, 1986. Mode of action of excitatory amino acid receptor antagonists on hippocampal long-lasting potentiation. Neuroscience, 17: 1105-1115.
CrossRefPubMedDirect Link - Youdim, K.A., J.P.E., Spencer, H. Schroeter and C. Rice-Evans, 2002. Dietary flavonoids as potential neuroprotectants. Biol. Chem., 383: 503-519.
PubMed - Zola-Morgan, S. and L.R. Squire, 1986. Memory impairment in monkeys following lesions limited to the hippocampus. Behav. Neurosci., 100: 155-160.
CrossRefPubMedDirect Link - Pachauri, S.D., S. Tota, K. Khandelwal, P.R.P. Verma and C. Nath et al., 2012. Protective effect of fruits of Morinda citrifolia L. on scopolamine induced memory impairment in mice: A behavioral, biochemical and cerebral blood flow study. J. Ethnopharmacol., 139: 34-41.
CrossRefPubMedDirect Link - Pandi-Perumal, S.R., A.S. BaHammam, G.M. Brown, D.W. Spence and V.K. Bharti et al., 2013. Melatonin antioxidative defense: Therapeutical implications for aging and neurodegenerative processes. Neurotox. Res., 23: 267-300.
CrossRefPubMedDirect Link - Tregub, P.P., V.P. Kulikov, A.G. Bespalov, A.J. Vvedensky and I.S. Osipov, 2013. Neuroprotective effects of individual or combined exposure to hypoxia and hypercapnia in the experiment. Bull. Exp. Biol. Med., 155: 327-329.
PubMed - Viswanatha, G.L., H. Shylaja and C.G. Mohan, 2013. Alleviation of transient global ischemia/reperfusion-induced brain injury in rats with 1,2,3,4,6-penta-O-galloyl-β-d-glucopyranose isolated from Mangifera indica. Eur. J. Pharmacol., 720: 286-293.
CrossRefPubMedDirect Link - Huy, P.D., Y.C. Yu, S.T. Ngo, T.V. Thao, C.P. Chen, M.S. Li and Y.C. Chen, 2013. In silico and in vitro characterization of anti-amyloidogenic activity of vitamin K3 analogues for Alzheimer's disease. Biochim. Biophys. Acta General Subjects, 1830: 2960-2969.
CrossRefPubMedDirect Link - Chang, H.C., Y.R. Yang, P.S. Wang, C.H. Kuo and R.Y. Wang, 2013. The neuroprotective effects of intramuscular insulin-like growth factor-i treatment in brain ischemic rats. PLoS ONE, Vol. 8.
CrossRefDirect Link