
A. Sarkaki
Ahvaz Physiology Research Center, Ahvaz Jundishpur University of Medical Sciences, Ahvaz, Iran
H. Fathimoghaddam
Department of Physiology, Ahvaz Jundishpur University of Medical Sciences, Ahvaz, Iran
S.M.T. Mansouri
Department of Pharmacology, Ahvaz Jundishpur University of Medical Sciences, Ahvaz, Iran
M. Shahrani Korrani
Medical Plant Research Center and Cellular, Molecular Research Center of Shahrekord University of Medical Sciences, Shahrekord, Iran
G. Saki
Department of Anatomical Sciences, School of Medical Sciences, Ahvaz Jundishpur University of Medical Sciences, Ahvaz, Iran
Y. Farbood
Medicinal Plant Research Center, Ahvaz Jundishpur University of Medical Sciences, Ahvaz, Iran
Pakistan Journal of Biological Sciences
Year: 2014 | Volume: 17 | Issue: 8 | Page No.: 978-990







ABSTRACT
Cerebral Hypoperfusion Ischemia (CHI) has important role in neuronal damage and behavioral deficits, including memory and Long-term Potentiation (LTP) impairment. Protective effects of Gallic Acid (GA) on memory, hippocampus LTP and cell viability were examined in permanent bilateral common carotid artery occlusion in rats. Animals were divided into 9 groups: Control (Cont); sham operated (Sho); Cerebral Hypoperfusion Ischemia (CHI); CHI received normal saline (CHI +Veh); CHI treated with different doses gallic acid (50, 100, 200 mg kg-1 for 5 days before and 5 days after CHI induction, orally); CHI treated with phenytoin (50 mg kg-1, ip) (CHI+Phe); and sham operated received 100 mg kg-1, orally (Sho+GA100). CHI was induced by bilateral common carotid artery occlusion (2VO). Behavioral, electrophysiological and histological evaluations were performed. Data were analyzed by one-way and repeated measures ANOVA followed by tukey’s post-hoc test. GA improved passive avoidance memory, hippocampal LTP and cell viability in hippocampus and cortex of ischemic rats significantly (p<0.01). The results suggest that gallic acid via its antioxidative and free radicals scavenging properties attenuates CHI induced behavioral and electrophysiological deficits and has significant protective effect on brain cell viability. Dose of 100 mg kg-1 GA has affected the ischemic but not intact rats and its effect was more potent significantly than phenytoin, a routine drug for ischemic subjects.
PDF Abstract XML References Citation
Received: December 05, 2013;
Accepted: February 13, 2014;
Published: March 29, 2014
How to cite this article
A. Sarkaki, H. Fathimoghaddam, S.M.T. Mansouri, M. Shahrani Korrani, G. Saki and Y. Farbood, 2014. Gallic Acid Improves Cognitive, Hippocampal Long-term Potentiation Deficits and Brain Damage Induced by Chronic Cerebral Hypoperfusion in Rats. Pakistan Journal of Biological Sciences, 17: 978-990.
DOI: 10.3923/pjbs.2014.978.990
URL: https://scialert.net/abstract/?doi=pjbs.2014.978.990
DOI: 10.3923/pjbs.2014.978.990
URL: https://scialert.net/abstract/?doi=pjbs.2014.978.990
INTRODUCTION
One of common pathophysiological states associated with central nervous system is chronic cerebral hypoperfusion that frequently occurs in conditions such as vascular dementia and Alzheimer's disease (Ozacmak et al., 2007). Hypoperfusion brain injury is caused by the interruption of cerebral blood flow leading to both acute and delayed degeneration of brain cells (Xu et al., 2009a). Oxidative stress plays an important role in animal models of brain ischemia (Horecky et al., 2011). Reactive Oxygen Species (ROS) formation represents the first key step to initiate oxidative damage (Silva-Adaya et al., 2008). Oxidative stress is defined as an imbalance between cellular antioxidant capacity and ROS (Kasparova et al., 2005). Focal disturbance of blood flow after brain ischemia involves increase of extracellular glutamate (Liu et al., 2010). Activation of glutamate receptors after cerebral hypoperfusion increases excitotoxicity and neuronal damage (Saransaari and Oja, 2010).
Vascular endothelial nitric oxide may play a critical role in spatial memory function during cerebral brain hypoperfusion and can result in spatial memory impairment (De La Torre and Aliev, 2005). Chronic cerebral hypoperfusion (CHI) caused progressive and long-lasting cognitive deficits (Tohda et al., 2004), perturbation in the memory and exacerbate cognitive impairment in rat (Choi et al., 2011b).
GA (2, 3 ,4-trihydroxybenzoic acid) is a natural phenolic compound which finds application in chemical industries such as dye-making, the tanning of leather and organic synthesis (Qi et al., 2009). It also has a wide range of biological activities such as antioxidant, anti-inflammatory, anti-microbial and anti-cancer. The free GA content of plants is very low, at a level which could not meet the demands of chemical industry (Kratz et al., 2008b). The synthetic n-alkyl esters of GA, also known as gallates, especially propyl, octyl and dodecyl gallates, are widely employed as antioxidants by food and pharmaceutical industries (Kratz et al., 2008a). GA has strong anti tyrosine’s activity (Kim, 2007) and protects the brain by improving antioxidant capacity and reducing inflammation in a rat model of permanent brain hypoperfusion (Mansouri et al., 2013).
Phenytoin is an anticonvulsant that has been used successfully to treat epilepsy for many years, but its potential as a mood stabilizer has only recently been evaluated in a controlled manner. Phenytoin up-regulates the expression of genes related to glutamate neurotransmission (Mariotti et al., 2010). On the other hand, phenytoin is known to interact with the voltage-dependent Na+ channels responsible for action potential generation and has positive effect on dura healing (Ergun et al., 2011). Chronic cerebral hypoperfusion, induced by 2VO, is related to neurological disorders and contributes to cognitive decline (He et al., 2012).
The aim of present study was to evaluate the effects of GA on memory, hippocampal long-term potentiation (LTP) and cell viability in rats with cerebral hypoperfusion/ischemi.
MATERIALS AND METHODS
Animals: Adult male Wistar rats (250-300 g) were obtained from the central animal house of the Shahrekord University of Medical Sciences, Shahrekord-Iran). They were maintained in a temperature-controlled room (21±2°C), on a 12/12 h light/dark cycle and with food and water available ad libitum. Animals were randomly divided into 9 experimental groups with 14 in each as following: (1) Control (Cont) (2) Sham-operated (Sho) (3) With Cerebral Hypoperfusion Ischemia (CHI) in which bilateral common carotid arteries were occluded (4) CHI received normal saline (2 mL kg-1, orally) (5-7) CHI received different doses gallic acid (CHI+GA50-200) (8) CHI received 50 mg kg-1, i.p. phenytoin (CHI+Phe) (Wu et al., 1989) (9) sham operated received 100 mg kg-1 GA (Sh+GA100) orally as positive control group. All procedures were done in accordance with the guides for the care and use of laboratory animals adopted by National Institute of Health and with the Federation of Iranian Societies for Experimental Biology.
Experimental design: GA administration was started 5 days before surgery to make CHI and continued for 5 consecutive days after CHI induction. On 5th day the 2VO surgery was applied. The treatment schedule and the intervals for estimation of various parameters have been presented in Fig. 1.
Permanent cerebral hypoperfusion induction: Permanent cerebral hypoperfusion was induced by occlusion of the bilateral common carotid arteries (2VO) by ligation with silk threads and cutting (Xu et al., 2012) under appropriate anesthesia with ketamine/xylazine (50/5 mg kg-1) (Roohbakhsh et al., 2007). A neck ventral midline incision was made and the common carotid arteries were then exposed and gently separated from the vagus nerve. The sham operated animals underwent a similar surgery but vessel ligation was excluded.
Neurological evaluation: Behavioral assessment such as sensorimotor tests were done to prove the ischemic brain damage in all groups of animals 5 days after the operation by an examiner who was blind to the type of surgical procedure. It consisted of six tests developed and described by (Garcia et al., 1995) with some modifications. The scores assigned to each rat at the end of the examination is the sum of the six tests scores. Each animal with summation score at least 3 for all six tests assign as CHI model in this study.
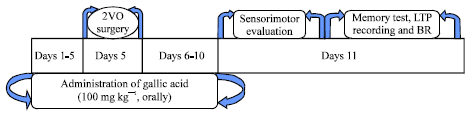 | |
| Fig. 1: | Design of experimental protocols. BR: Brain removing for histological and molecular studies |
Spontaneous activity: The animal was observed for 5 min in its normal environment. The rat’s activity was assessed by its ability to approach all four walls of the cage. Scores were given as following: score 3, rat moved around, explored the environment and approached at least three walls of the cage; score 2, slightly affected rat moved about in the cage but did not approach all sides and hesitated to move, although it eventually reached at least one upper rim of the cage; score 1, severely affected rat did not rise up at all and barely moved in the cage and score 0, rat did not move at all.
Symmetry in the movement of four limbs: The rat was held in the air by the tail to observe symmetry in the movement of the four limbs. Scores were given as following: score 3, all four limbs extended symmetrically; score 2, limbs on left side extended less or more slowly than those on the right; score 1, limbs on left side showed minimal movement and score 0, forelimb on left side did not move at all.
Forepaw outstretching: The rat was brought up to the edge of the table and made to walk on forelimbs while being held by the tail. Symmetry in the outstretching of both forelimbs was observed while the rat reached the table and the hind limbs were kept in the air. Scores were given as following: score 3, both forelimbs were outstretched and the rat walked symmetrically on forepaws; score 2, left side outstretched less than the right and forepaw walking was impaired; score 1, left forelimb moved minimally and score 0, left forelimb did not move.
Climbing: The rat was placed on the wall of a wire cage. Normally the rat uses all four limbs to climb up the wall. When the rat was removed from the wire cage by pulling it off by the tail, the strength of attachment was noted. Scores were given as following: score 3, rat climbed easily and gripped tightly to the wire; score 2, left side was impaired while climbing or did not grip as hard as the right side and score 1, rat failed to climb or tended to circle instead of climbing.
Body proprioception: The rat was touched with a blunt stick on each side of the body and the reaction to the stimulus was observed. Scores were given as following: score 3, rat reacted by turning head and was equally startled by the stimulus on both sides; score 2, rat reacted slowly to stimulus on left side and score 1, rat did not respond to the stimulus placed on the left side.
Response to vibrissae touch: A blunt stick was brushed against the vibrissae on each side; the stick was moved toward the whiskers from the rear of the animal to avoid entering the visual fields. Scores were given as following: score 3, rat reacted by turning head or was equally startled by the stimulus on both sides; score 2, rat reacted slowly to stimulus on left side and score 1, rat did not respond to stimulus on the left side.
The score given to each rat at the completion of the evaluation is the summation of all six individual test scores. The minimum neurological score is 3 and the maximum is 18.
Passive avoidance test: The passive avoidance apparatus (Shuttle box) consisted of two illuminated/darken compartments. The cognitive test was performed at 8:00-11:00 am. The rat was placed in the lighted compartment while a guillotine door was opened and allowed to explore into both compartments for 5 min. After 10 min the rat was placed in the lighted compartment again facing away from the closed guillotine door and ten seconds later the door was raised and the entering delay of rat into the darken compartment was recorded as Initial Latency (IL). After then the guillotine door was closed and a 50 Hz square wave, 1.2 mA constant current electrical single shock was delivered to rat foot paws for 1.5 sec. On the retention test that given 24 h after the acquisition trial, the rat was again placed into the illuminated compartment and the step-through latency (STL) and the time spent in the darken compartment were recorded as a measure of retention performance (Lashgari et al., 2006). The maximum latency was 300 sec.
Electrophysiological studies
Surgery: Rats were anesthetized with intraperitoneal injection of ketamine/xylazine (90/10 mg kg-1) and their heads mounted in a stereotaxic device for surgery (electrode implantation and EPSP recording). A heating pad was used to maintain the animals’ body temperature at 36.5±0.5°C. The bipolar metal wire recording and stimulating microelectrodes were positioned in the granular cells of DG (tungsten wire, CFW, USA) with stereotaxic coordination of AP= -3.8 mm from bregma; ML= -2.3 mm; DV= -3.5 mm from dura (Paxinos and Waston, 2006) and Perforant Pathway (PP) (stainless steel; 0.125 mm diameter, Advent Co., UK) at AP: -7.5 from bregma, ML: -4, DV: -3.9 mm from the dura, respectively (Roohbakhsh et al., 2007). Implantation of electrodes at the correct position was determined by fEPSP recording (Lashgari et al., 2006) and histological verification at the end of experiments.
Electrophysiological recordings and LTP induction: The field potential recordings were obtained in the granular cells of the DG following stimulation of the PP. Test stimuli were delivered to the PP every 30 sec. Electrodes were positioned to elicit a maximal field Population Spike (PS) and field excitatory post synaptic potential (fEPSP). The PS amplitude was measured as the difference of voltage between the peak of the first positive wave and the peak of the first negative deflection. The slope of fEPSP was measured as the maximum slope between initial point of fEPSP and the first positive peak of wave in order to measure synaptic efficacy. Extracellular field potentials were amplified (1000x); band pass filtered between 0.1Hz-3kHz, digitized and recorded, after then analyzed with using potentialize software (Science Beam Co. Version 1.107, Iran). LTP was induced by using high-frequency stimuli (HFS) protocols of 400 Hz (10 bursts of 20 stimuli, 0.2 ms stimulus duration, 10 sec interburst interval) at a stimulus intensity that evoked a PS amplitude of approximately 80% of maximum response. All potentials employed as baseline before and after HFS were evoked at a stimulus intensity which produced 40% of its maximum amplitude by input/output (I/O) curve with different intensities for LTP recording. Both fEPSP and PS were recorded in the periods of 5, 15, 30, 45 and 120 min after the HFS in order to determine any changes in the synaptic response of DG neurons (Lashgari et al., 2006).
Histology: At the end of electrophysiological evaluations rats were deeply anesthetized with chloral hydrate (350 mg kg-1) and perfused transcardially by 200 mL of normal saline followed by 600 mL of 4% paraformaldehyde solution. The brains were removed and immersion-fixed in 5% paraformaldehyde solution for 10 days. Afterward, each brain was dehydrated in graded ethanol solutions and embedded in paraffin. Coronal serial sections with 5 μm thickness were cut. Ten sections including hippocampal formation and cerebral cortex were selected by random systemic sampling from each animal and stained by hematoxylin-eosin. The cytoarchitectonic borders of granular layer of DG were defined according to standard cytoarchitectonical criteria. The granular layer of hippocampus in both hemispheres was studied under a light microscope (BX51, Olympus, Japan) coupled to a digital camera (objective lensx100; Olympus, Japan). The magnification was calculated by an objective micrometer. For counting of dark neurons on each section, at least ten microscopic fields were selected by uniform systematic random sampling in granular layer of hippocampus. Glial cells were distinguished from neurons based on their nuclear shape, size, cytoplasm location and characteristic staining (Nazem et al., 2012).
Statistical analysis: Data were expressed as Mean±SEM of mean and processed by SPSS ver.17. Data from the behavioral and electrophysiological experiments were analyzed by one-way and two-way Analysis of Variance (ANOVA) with repeated-measures for lesion and treatments as independent variables and session as the repeated measure. All analyses were followed by post hoc Tukey’s test. P-value less than 0.05 considered to be statistically significant. The symbols * and # mean differences with sham operated and cerebral hypoferfusion/ischemia groups, respectively.
RESULTS
No differences were seen between control (Cont) and sham operated (Sho) groups during analyzing the all parameters. So, the other groups were compared only with Sho group in all experiments.
Sensorimotor score: Figure 2 shows the sensorimotor scores 5 days after 2VO operation. There was significant sensorimotor impairment in CHI rats when compared to Sho group (p<0.001). Treatment with GA started 5 days before the 2VO surgery for 10 days improved sensorimotor scores in CHI+GA100 and CHI+GA 200, while there was not any difference between them (p<0.001 vs. CHI). No significant differences were between CHI+Phe with CHI groups and between Sho with Sho+GA100 groups, respectively.
Passive avoidance memory: The Step Through Latency (STL) in sham operated (Sho), CHI without any treatment and CHI groups treated with GA, normal saline or phenytoin have shown in Fig. 3.
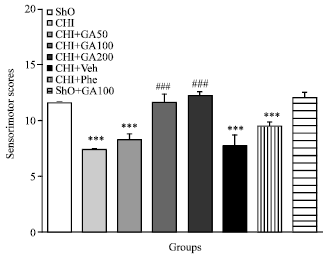 | |
| Fig. 2: | Sensorimotor scores 5 days after 2VO operation in CHI group. Treatment with GA started 5 days before the 2VO surgery for 10 days improved sensorimotor scores in CHI+GA100 and CHI+GA 200 (p<0.001 vs. CHI). No significant differences were between CHI+Phe with CHI groups and between Sho with Sho+GA100 groups, respectively |
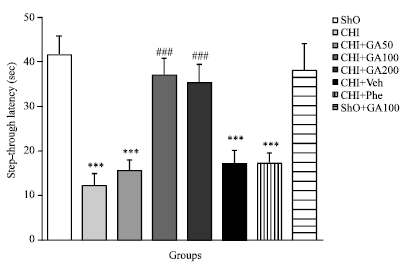 | |
| Fig. 3: | Mean±SEM of step down Latency (SDL) in Sham operated (ShO), CHI, CHI+Veh, CHI+GA (50, 100 and 200 mg kg-1, orally), CHI+Phenytoin (CHI+Phe) and Sho+GA100 groups during passive avoidance memory (***p<0.001 vs. Sho and ### p<0.001 vs. CHI group, n = 14, one way ANOVA followed by Tukey’s post hoc test) |
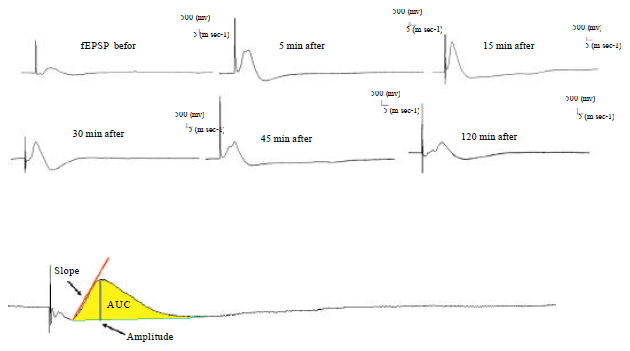 | |
| Fig. 4: | Recorded PS (LTP) traces before (as base EPSP) and 15, 30, 45 and 120 min after high frequency stimulation (HFS, 400 Hz) in different groups |
STL decreased significantly in CHI and CHI+Veh groups when compared with Sho group (p<0.001). STL was significantly increased in CHI+GA100 and CHI+GA200 groups (with same effect) when compared to CHI group (p<0.001). In this case there was no significant difference between CHI+Phe with CHI or CHI+Veh groups.
Electrophysiological results: Figure 4 shows that recorded PS traces before and 15, 30, 45 and 120 min after high frequency stimulation (HFS, 400 Hz) in different groups.
PS amplitude: As shown in Fig. 5, the PS amplitude (% PS Amp) was not difference between control (Cont) and sham operated (Sho) groups. Amp decreased significantly in CHI during all LTP recording times when compared with sham operated group (p<0.001, Fig. 5a). PS amplitude was improved significantly (p<0.00, Fig. 5b) in CHI+GA100 and CHI+GA200 groups (with same effect) during all LTP recording times when compared with CHI or CHI+Veh (CHI+GA200 group didn’t show here).
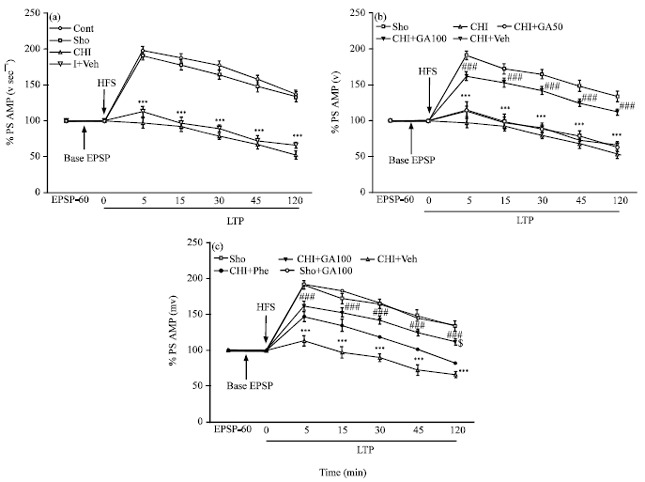 | |
| Fig. 5(a-c): | Mean±SEM of percents of amplitude of Population Spikes (PS) in different groups during basal fEPSP and LTP recorded from hippocampal Dentate Gyrus (DG) at different times after High Frequency Stimulation (HFS) to brain perforant path (PP), (a) Cont, Sho, CHI and CHI+Veh groups (b), in Sho, CHI, CHI+GA50, CHI+GA100 and CHI+Veh groups and (c) Sho, CHI+GA100, CHI+Phe and Sho+GA100 groups. (Repeated measures two-way ANOVA, followed by Tukey’s post hoc test, (n=14, ***p<0.001 vs. Sho group and ### p<0.001 vs. CHI group) |
The effect of gallic acid was more effective than phenytoin as a routine drug used in cerebral ischemic subjects to improve PS amplitude during all LTP recording times in 2VO rats, but it was significant only during 120th recording time (p<0.05). There was no difference between PS amplitude of Sho+GA100 and Sho group (Fig. 5c).
PS slope: As shown in Fig. 6, the PS slope has no difference between control (Cont) and sham operated (Sho) groups. The slope of population spikes in CHI group was significantly lower than Sho group during all LTP recording times (p<0.001, Fig. 6a). The PS slope in CHI group treated with 100 mg kg-1 GA (CHI+GA100) was improved significantly when compared with CHI or CHI+Veh groups (p<0.001, Fig. 6b). Treatment ischemic groups with gallic acid 100 and 200 mg kg-1 (with same effects) improved the PS slope more power than currently medication with 50 mg kg-1 phenytoin for brain ischemia (CHI+GA200 groups didn’t show here) (p<0.001, Fig. 6c).
PS AUC: As shown in Fig. 7, Area Under the Curve (AUC) of Population Spike (PS) wasn’t different in control and sham operated groups while AUC in CHI group significantly decreased in comparing to sham operated group (Sho) during all LTP recording times after HFS (p<0.001, Fig. 7a). There was no significance difference between AUC of PS in CHI, CHI+Veh and CHI+GA50 groups. Treatment with GA 100 and 200 mg kg-1 (with same effects) increased AUC of PS during all LTP recording times after HFS in comparing to CHI or CHI+Veh groups (p<0.001, Fig. 7b).
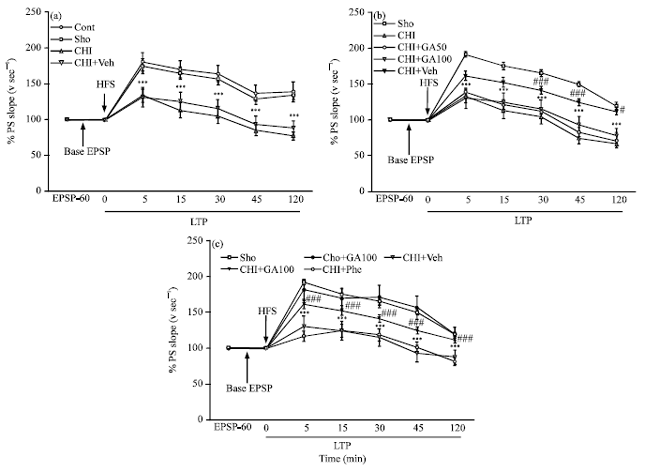 | |
| Fig. 6(a-c): | Mean±SEM of percents of slope of Population Spikes (PS) in different groups during basal fEPSP and LTP recorded from hippocampal Dentate Gyrus (DG) at different times after High Frequency Stimulation (HFS) to brain Perforant Path (PP), (a) Cont, Sho, CHI and CHI+Veh groups, (b) Sho, CHI, CHI+GA50, CHI+GA100 and CHI+Veh groups and (c) Sho, CHI+GA100, CHI+Phe and Sho+GA100 groups. (Repeated measures two-way ANOVA, followed by Tukey’s post hoc test, (n=14, ***p<0.001 vs. Sho group and ### p<0.001 vs. CHI group) |
As shown in Fig. 6c there were no differences between CHI+Veh with CHI+Phe and Sho with Sho+GA100 groups, but CHI groups treated with doses 100 and 200 mg kg-1 GA (not shown GA+200 group at here) improved AUC of PS during 5, 15 and 30 min after HFS significantly (p<0.05, Fig. 7c). While treatment the ischemic group with phenytoin didn’t change the AUC when compared to CHI+Veh group.
Cell viability in hippocampus and cerebral cortex: The cell viability was measured to evaluate the protective potential of GA on the cells against oxidative stress in brain regions. As shown in Fig. 8 cell viability level in CHI rats were found to be significantly depleted when compared with Sho group in the hippocampus and cerebral cortex (p<0.001, Fig. 8a and b). Treatment the CHI rats with GA but not phenytoin (CHI+GA100 or CHI+GA200) was able to raise hippocampus cells viability levels in comparing to CHI group (p<0.05, p<0.001, Fig. 8a). Cell viability level in CHI+GA200 significantly prevented neuronal depletion in cerebral cortex (p<0.01, Fig. 8b).
Figure 9 shows a high number of dead neurons in CHI group when compared with Sho (Fig. 9a, b). Treatment the CHI rats with GA (100 or 200 mg kg-1, orally) significantly prevented neuronal damage in both cerebral cortex and hippocampus (Fig. 9c).
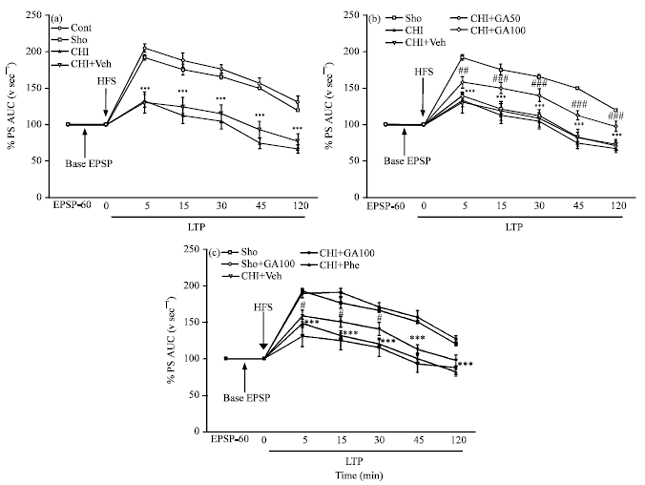 | |
| Fig. 7(a-c): | Mean±SEM of percents of Area Under Curve (AUC) of Population Spikes (PS) in different groups during basal fEPSP and LTP recorded from hippocampal Dentate Gyrus (DG) at different times after High Frequency Stimulation (HFS) to brain Perforant Path (PP) (a) Cont, Sho, CHI and CHI+Veh groups, (b) Sho, CHI, CHI+GA50, CHI+GA100 and CHI+Veh groups and (c) Sho, CHI+GA100, CHI+Phe and Sho+GA100 groups. (Repeated measures two-way ANOVA, followed by Tukey’s post hoc test, (n = 14, ***p<0.001 vs. Sho group and #p<0.05 and ### p<0.001 vs. CHI group) |
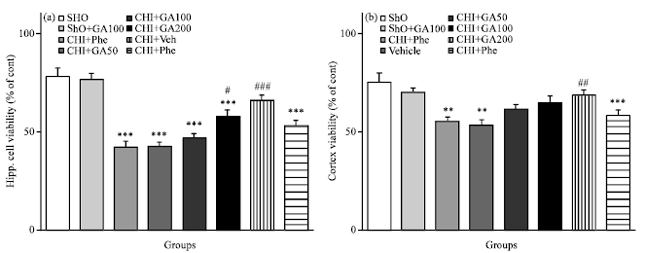 | |
| Fig. 8(a-b): | Cell viability level of hippocampus and cerebral cortex in different groups. Cell viability in CHI rats were found to be significantly depleted when compared with Sho group (p<0.001, (a and b). Treatment the CHI rats with GA but not phenytoin was able to raise cells viability levels in hippocampus and cerebral cortex when compared to CHI group (p<0.05, p<0.001) |
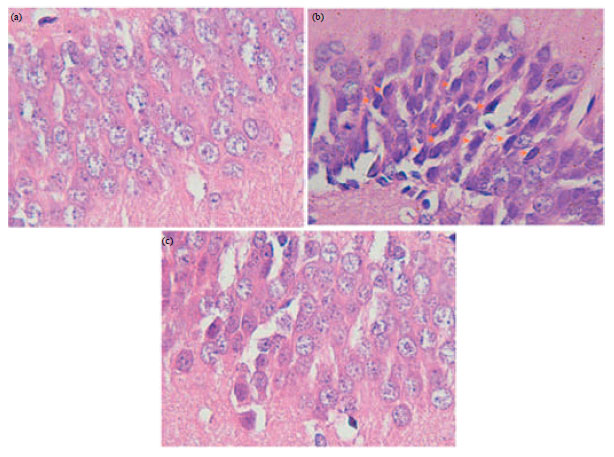 | |
| Fig. 9(a-c): | (a) High No. of dead neurons in CHI group (b) When compared with Sho group and (c) Treatment the CHI rats with GA (100 or 200 mg kg-1, orally) significantly prevented neuronal damage in hippocampus (c) |
DISCUSSION
Permanent occlusion of the common carotid arteries in rats is an established procedure to investigate the effects of chronic cerebral hypoperfusion on brain cognitive, electrophysiological dysfunctions and neurodegenerative processes (Cechetti et al., 2012b). Here we have reported that GA is effective in recovering avoidance memory deficit in rats when applied before and after the ischemic event (Cechetti et al., 2012a). Reduction of cerebral blood flow is an important risk factor for dementia states and other brain dysfunctions. We demonstrate that chronic brain hypoperfusion leads to memory impairment, brain electrophysiology, sensorimotor disturbance, decrease spontaneous activity, loss of symmetry in the movement of four limbs and abnormality in gait performance. Cognitive deficits were described earlier for this experimental model and indicate the adequacy for studying vascular dementia (Cechetti et al., 2010). It has been suggested that the functional cognitive deficits after hypoperfusion are associated with impairment of synaptic transmission, lower number of pyramidal neurons and induced neurodegeneration in the hippocampal CA1 region (Cechetti et al., 2012b). The present study indicated that GA dramatically protected the brain against chronic brain hypoperfusion-induced deterioration of memory in rat. Our study showed that the chronic brain hypoperfusion model was an efficient model of neurodegenerative disorders as shown before and well established experimental model to investigate neuronal damage and cognitive impairment that occurs in human after CHI, ageing and Alzheimer's disease (Vicente et al., 2009).
Cognition is a collective term for higher cortical functions such as thinking, remembering, knowing, planning and analyzing. Cognition is crucial for a person to become aware of his/her situation, needs and goals and meet the challenges of daily life (Dardiotis et al., 2012). Pathophysiology of cognitive impairment in heart failure is also the development of cerebral abnormalities as a result of chronic hypoperfusion (Shibata et al., 2004). Chronic cerebral hypoperfusion induces microvascular changes that could contribute to the progression of vascular cognitive impairment and dementia in the aging brain (Hai et al., 2013; Marquez-Martin et al., 2012). Our results in this experiment are consistent with previous studies that replicated memory impairment in rats with CHI (Zhang et al., 2013). General cognitive dysfunction were most correlated with cerebral hypoperfusion (Yoon et al., 2012).
Hippocampus is very sensitive to ischemia and the some its subdivisions are characterized by a low capillary density as compared with the neighboring other subdivisions (Cavaglia et al., 2001; Farkas et al., 2007). In other hand, on base of knowledge hippocampus plays a crucial role in learning and memory processing. Thus it is very sensitive to CHI in relation to other areas of brain. Our results showed that 2VO to weaken memory and Population Spikes (PS) recorded from hippocampus DG subdivision. Several investigations such as following reports have already established neuronal damage due to cerebral ischemia. Neuronal cell death is caused by a serial pathophysiological events after cerebral ischemic stroke, so called ‘ischemic cascade’ like energy failure, excitotoxicity, oxidative stress, inflammation, apoptosis (Dahiya et al., 2010). Ischemia causes acute necrotic death in the "core" of the ischemic area by leading to resting membrane potential disruption and neuronal swelling (Dirnagl et al., 1999; Saulle et al., 2002). So, cerebral ischemia could weaken and disrupt the synaptic transition and hippocampal PS in animals as well as humans (Squire and Zola, 1996). In addition, it seems a few cells of brain will damage or die after ischemia due to intracellular Ca+accumulation induced by activation of NMDA receptors in important brain area. Certain studies showed that 2VO causes an increase in the NMDA receptor density in the hippocampus and excessive Ca+2 influx through NMDA receptors is a major mechanism for neurodegeneration following stroke and brain trauma (Shinno et al., 1997). So, brain ischemia causes the rise in the glutamate (Glu) level in the brain interstitial fluid (Davalos et al., 2000). Decrease of this excess Glu may improve the consequences and outcome of ischemic conditions (Gottlieb et al., 2003). This finding is consistent with some previous findings (Shinno et al., 1997; Farkas et al., 2007).
GA is a well known natural compound with potent antioxidant and free radical scavenging abilities (Ding et al., 2012). In nature, GA metabolites are found in two forms, methylated gallic acids form and galloyl conjugates of catechin (Nabavi et al., 2012). In addition to their antioxidant effects, there are numerous reports on their other biological activities such as its anti-cancer and anti-apoptotic potentials. Ligustilide was a neuroprotective agent for treating chronic cerebral hypoperfusion injury, which may be attributed to its anti-apoptosis of neuron and anti-proliferation of astrocyte both in cortex and in hippocampus of 2VO rats (Feng et al., 2012).
So, GA likely has a protective effect against neurotoxicity due to NMDA receptors sensitivity and excitotoxicity induced by glutamate after cerebral ischemia that followed by ca+ influx and thereby intracellular ca+ accumulation induced neuronal apoptosis. In other hand, GA with its antioxidative effect may oppose with NMDA receptors activation and thereby has a protective effect for neurotoxicity and/or excititoxicity following CHI.
Hypothesized that the cerebral injury of CHI may causes inflammation and demyelination (Rosenberg, 2009) and excess Reactive Oxygen Species (ROS) (Xu et al., 2009b) in the brain tissues and has been suggested as a key pathological mechanisms of chronic cerebral hypoperfusion. Highly adhesive glycoprotein von Willebrand factor multimer induces platelet aggregation and leukocyte tethering on the injured vascular wall, contributing to microvascular plugging and inflammation in brain hypoperfusion (Fujioka et al., 2012). Previous studies shown that hippocampal, septohippocampal or reticular thalamic nucleus lesions, have detrimental effects on performances in many memory tasks (Karson et al., 2012) and in this study cognition deficit, LTP reduction, attenuate cell viability in cerebral cortex and hippocampus after CHI can be result of oxidative stress in this parts of brain (data have not shown here). Also cognitive decline induced by CHI could be related to dysfunction of the basal forebrain cholinergic system and reduction of hippocampal mitogen-activated protein kinases activities (Choi et al., 2011a). Chronic cerebral hypoperfusion is thought to induce white matter lesions with oligodendrocyte death and myelin breakdown. There is consistent evidence that cerebral hypoperfusion causes in memory deficits (Cai et al., 2011). Activation of the Rho/Rho-kinase pathway is related to the neuronal damage and the pathogenesis of the memory impairment in CHI rats (Huang et al., 2008).
CONCLUSION
Our data show that GA could improve behavioral, electrophysiological and cell viability of hippocampus and cerebral cortex by attenuate oxidative stress in the hypoperfusion brain tissues. In other hand, GA is more effective than phenytoin, a routine drug for cerebral ischemia in CHI rats. GA had no effects on behavioral, electrophysiological properties and brain cell viability in healthy rats.
ACKNOWLEDGMENT
This article was extracted as a part of Mr. Mehrdad shahrani Korrani's Ph.D. thesis. This research was supported by Physiology Research Center, Vice Chancellor of Research, Ahvaz Jundishapur University of Medical Sciences, Iran (Grant No: PRC-75). The authors would like to thank Researchers Eimani, Rafieian, hodjati, Rabiei and Alibabaei, our colleagues in Shahrekord University of Medical Sciences (Iran), for kindly help in technical support.
REFERENCES
- Cai, Q.Y., X.S. Chen, X.L. Zhan and Z.X. Yao, 2011. Protective effects of catalpol on oligodendrocyte death and myelin breakdown in a rat model of chronic cerebral hypoperfusion. Neurosci. Lett., 497: 22-26.
CrossRef - Cechetti, F., A.S. Pagnussat, P.V. Worm, V.R. Elsner and J. Ben et al., 2012. Chronic brain hypoperfusion causes early glial activation and neuronal death and subsequent long-term memory impairment. Brain Res. Bull., 87: 109-116.
CrossRef - Cechetti, F., P.V. Worm, G. Lovatel, F. Moyses, I.R. Siqueira and C.A. Netto, 2012. Environmental enrichment prevents behavioral deficits and oxidative stress caused by chronic cerebral hypoperfusion in the rat. Life Sci., 91: 29-36.
CrossRef - Cechetti, F., P.V. Worm, L.O. Pereira, I.R. Siqueira and C.A. Netto, 2010. The modified 2VO ischemia protocol causes cognitive impairment similar to that induced by the standard method, but with a better survival rate. Brazil. J. Med. Biol. Res., 43: 1178-1183.
CrossRefDirect Link - Choi, B.R., K.J. Kwon, S.H. Park, W.K. Jeon, S.H. Han, H.Y. Kim and J.S. Han, 2011. Alternations of septal-hippocampal system in the adult wistar rat with spatial memory impairments induced by chronic cerebral hypoperfusion. Exp. Neurobiol., 20: 92-99.
CrossRefDirect Link - Choi, B.R., S.R. Lee, J.S. Han, S.K. Woo and K.M. Kim et al., 2011. Synergistic memory impairment through the interaction of chronic cerebral hypoperfusion and amlyloid toxicity in a rat model. Stroke, 42: 2595-2604.
Direct Link - Dahiya, K., K. Aggarwal, S. Seth, V. Singh and T.K. Sharma, 2010. Type 2 diabetes mellitus without vascular complications and ischemia modified albumin. Clin. Lab., 56: 187-190.
PubMed - Dardiotis, E., G. Giamouzis, D. Mastrogiannis, C. Vogiatzi, J. Skoularigis, F. Triposkiadis and G.M. Hadjigeorgiou, 2012. Cognitive impairment in heart failure. Cardiol. Res. Pract.
CrossRef - Davalos, A., A. Shuaib and N.G. Wahlgren, 2000. Neurotransmitters and pathophysiology of stroke: Evidence for the release of glutamate and other transmitters/mediators in animals and humans. J. Stroke Cerebrovasc. Dis., 9: 2-8.
CrossRefDirect Link - De La Torre, J.C. and G. Aliev, 2005. Inhibition of vascular nitric oxide after rat chronic brain hypoperfusion: Spatial memory and immunocytochemical changes. J. Cerebral Blood Flow Metab., 25: 663-672.
CrossRef - Ding, W., D. Qian, J. Duan, E. Shang, Y. Tang, Y. Qian and N. Luo, 2012. Study on metabolites of ferulic acid and gallic acid compatible with danggui and chishao in rat plasma and urine. Zhongguo Zhong Yao Za Zhi, 37: 366-372.
PubMed - Dirnagl, U., C. Iadecola and M.A. Moskowitz, 1999. Pathobiology of ischaemic stroke: An integrated view. Trends Neurosci., 22: 391-397.
CrossRefPubMedDirect Link - Ergun, E., G. Kurt, M. Tonge, H. Aytar, M. Tas, K. Baykaner and N. Ceviker, 2011. Effects of phenytoin sodium on dura mater healing in a rat model of CSF leakage. Turk. Neurosurg., 21: 471-476.
PubMedDirect Link - Farkas, E., P.G. Luiten and F. Bari, 2007. Permanent, bilateral common carotid artery occlusion in the rat: A model for chronic cerebral hypoperfusion-related neurodegenerative diseases. Brain Res. Rev., 54: 162-180.
CrossRef - Feng, Z., Y. Lu, X. Wu, P. Zhao and J. Li et al., 2012. Ligustilide alleviates brain damage and improves cognitive function in rats of chronic cerebral hypoperfusion. J. Ethnopharmacol., 144: 313-321.
CrossRefDirect Link - Fujioka, M., T. Nakano, K. Hayakawa, K. Irie and Y. Akitake et al., 2012. ADAMTS13 gene deletion enhances plasma high-mobility group box1 elevation and neuroinflammation in brain ischemia-reperfusion injury. Neurol. Sci., 33: 1107-1115.
CrossRef - Garcia, J.H., S. Wagner, K.F. Liu and X.J. Hu, 1995. Neurological deficit and extent of neuronal necrosis attributable to middle cerebral artery occlusion in rats. Statistical validation. Stroke, 26: 627-634.
PubMedDirect Link - Gottlieb, M., Y. Wang and V.I. Teichberg, 2003. Blood‐mediated scavenging of cerebrospinal fluid glutamate. J. Neurochem., 87: 119-126.
CrossRef - He, X.L., Y.H. Wang, M.G. Bi and G.H. Du, 2012. Chrysin improves cognitive deficits and brain damage induced by chronic cerebral hypoperfusion in rats. Eur. J. Pharmacol., 680: 41-48.
CrossRef - Huang, L., Z. He, L. Guo and H. Wang, 2008. Improvement of cognitive deficit and neuronal damage in rats with chronic cerebral ischemia via relative long-term inhibition of rho-kinase. Cell. Mol. Neurobiol., 28: 757-768.
CrossRef - Hai, J., Y.F. Wu, Q. Lin, X.S. Huang and G.Y. Zhang, 2013. Cerebral blood flow and metabolic changes in hippocampal regions of a modified rat model with chronic cerebral hypoperfusion. Acta Neurologica Belgica, 113: 313-317.
CrossRef - Karson, A., T. Utkan, F. Balci, F. Aricioglu and N. Ates, 2012. Age-dependent decline in learning and memory performances of WAG/Rij rat model of absence epilepsy. Behav. Brain Funct., Vol. 8.
CrossRef - Kim, Y.J., 2007. Antimelanogenic and antioxidant properties of gallic acid. Biol. Pharmaceut. Bull., 30: 1052-1055.
CrossRefDirect Link - Kratz, J.M., C.R. Andrighetti-Frohner, D.J. Kolling, P.C. Leal and C.C. Cirne-Santos et al., 2008. Anti-HSV-1 and anti-HIV-1 activity of gallic acid and pentyl gallate. Mem. Inst. Oswaldo. Cruz., 103: 437-442.
CrossRef - Kratz, J.M., C.R. Andrighetti-Frohner, P.C. Leal, R.J. Nunes and R.A. Yunes et al., 2008. Evaluation of anti-HSV-2 activity of gallic acid and pentyl gallate. Biol. Pharmaceut. Bull., 31: 903-907.
CrossRefDirect Link - Liu, X., C. Hunter, H.R. Weiss and O.Z. Chi, 2010. Effects of blockade of ionotropic glutamate receptors on blood-brain barrier disruption in focal cerebral ischemia. Neurol. Sci., 31: 699-703.
CrossRef - Mansouri, M.T., Y. Farbood, M.J. Sameri, A. Sarkaki, B. Naghizadeh and M. Rafeirad, 2013. Neuroprotective effects of oral gallic acid against oxidative stress induced by 6-hydroxydopamine in rats. Food Chem., 138: 1028-1033.
CrossRef - Mariotti, V., E. Melissari, S. Amar, A. Conte, R.H. Belmaker, G. Agam and S. Pellegrini, 2010. Effect of prolonged phenytoin administration on rat brain gene expression assessed by DNA microarrays. Expt. Biol. Med., 235: 300-310.
PubMedDirect Link - Marquez-Martin, A., F. Jimenez-Altayo, A.P. Dantas, L. Caracuel, A.M. Planas and E. Vila, 2012. Middle cerebral artery alterations in a rat chronic hypoperfusion model. J. Applied Physiol., 112: 511-518.
CrossRefPubMedDirect Link - Nabavi, S.F., S. Habtemariam, M. Jafari, A. Sureda and S.M. Nabavi, 2012. Protective role of gallic acid on sodium fluoride induced oxidative stress in rat brain. Bull. Environ. Contamin. Toxicol., 89: 73-77.
CrossRef - Nazem, A., A.H. Jafarian, S.H. Sadraie, A. Gorji, H. Kheradmand, M. Radmard and H. Haghir, 2012. Neuronal injury and cytogenesis after simple febrile seizures in the hippocampal dentate gyrus of juvenile rat. Child Nerv. Syst., 28: 1931-1936.
CrossRef - Ozacmak, V.H., H. Sayan, A. Cetin and A. Akyildiz-Igdem, 2007. AT1 receptor blocker candesartan-induced attenuation of brain injury of rats subjected to chronic cerebral hypoperfusion. Neurochem. Res., 32: 1314-1321.
CrossRef - Qi, F.H., T.Z. Jing, Z.X. Wang and Y.G. Zhan, 2009. Fungal endophytes from Acer ginnala Maxim: Isolation, identification and their yield of gallic acid. Lett. Applied Microbiol., 49: 98-104.
CrossRefDirect Link - Roohbakhsh, A., A.H. Moghaddam, R. Massoudi and M.R. Zarrindast, 2007. Role of dorsal hippocampal cannabinoid receptors and nitric oxide in anxiety like behaviours in rats using the elevated plus‐maze test. Clin. Exp. Pharmacol. Physiol., 34: 223-229.
CrossRef - Rosenberg, G.A., 2009. Inflammation and white matter damage in vascular cognitive impairment. Stroke, 40: S20-S23.
PubMedDirect Link - Saransaari, P. and S.S. Oja, 2010. Modulation of taurine release in ischemia by glutamate receptors in mouse brain stem slices. Amino Acids, 38: 739-746.
CrossRef - Saulle, E., D. Centonze, A.B. Martin, R. Moratalla, G. Bernardi and P. Calabresi, 2002. Endogenous dopamine amplifies ischemic long-term potentiation via D1 receptors. Stroke, 33: 2978-2984.
CrossRef - Shibata, M., R. Ohtani, M. Ihara and H. Tomimoto, 2004. White matter lesions and glial activation in a novel mouse model of chronic cerebral hypoperfusion. Stroke, 35: 2598-2603.
CrossRef - Shinno, K., L. Zhang, J.H. Eubanks, P.L. Carlen and M.C. Wallace, 1997. Transient ischemia induces an early decrease of synaptic transmission in CA1 neurons of rat hippocampus: Electrophysiologic study in brain slices. J. Cerebral Blood Flow Metab., 17: 955-966.
CrossRef - Silva-Adaya, D., V. Perez-De La Cruz, M.N. Herrera-Mundo, K. Mendoza-Macedo and J. Villeda-Hernandez et al., 2008. Excitotoxic damage, disrupted energy metabolism, and oxidative stress in the rat brain: Antioxidant and neuroprotective effects of l‐carnitine. J. Neurochemistry, 105: 677-689.
CrossRefDirect Link - Squire, L.R. and S.M. Zola, 1996. Ischemic brain damage and memory impairment: A commentary. Hippocampus, 6: 546-552.
Direct Link - Tohda, M., P. Suwanakitch, R. Jeenapongsa, H. Hayashi, H. Watanabe and K. Matsumoto, 2004. Expression changes of the mRNA of Alzheimer's disease related factors in the permanent ischemic rat brain. Biol. Pharm. Bull., 27: 2021-2023.
CrossRef - Vicente, E., D. Degerone, L. Bohn, F. Scornavaca and A. Pimentel et al., 2009. Astroglial and cognitive effects of chronic cerebral hypoperfusion in the rat. Brain Res., 1251: 204-212.
CrossRef - Wu, H.Q., L. Xie, X.N. Jin, Q. Ge, H. Jin and G.Q. Liu, 1989. The effect of vanillin on the fully amygdala-kindled seizures in the rat. Yao Xue Xue Bao, 24: 482-486.
PubMed - Xu, L., L.A. Voloboueva, Y. Ouyang, J.F. Emery and R.G. Giffard, 2009. Overexpression of mitochondrial Hsp70/Hsp75 in rat brain protects mitochondria, reduces oxidative stress and protects from focal ischemia. J. Cerebral Blood Flow Metab., 29: 365-374.
CrossRefDirect Link - Xu, X., Z. Li, Z. Yang and T. Zhang, 2012. Decrease of synaptic plasticity associated with alteration of information flow in a rat model of vascular dementia. Neuroscience, 206: 136-143.
CrossRef - Xu, Y., S.M. Liachenko, P. Tang and P.H. Chan, 2009. Faster recovery of cerebral perfusion in SOD1-overexpressed rats after cardiac arrest and resuscitation. Stroke, 40: 2512-2518.
CrossRef - Yoon, H.J., K.W. Park, Y.J. Jeong and D.Y. Kang, 2012. Correlation between neuropsychological tests and hypoperfusion in MCI patients: Anatomical labeling using xjView and talairach daemon software. Ann. Nuclear Med., 26: 656-664.
CrossRef - Zhang, H.N., J. Wu, T. Jin, M. Chang and L. Sun, 2013. Transient elevation of synaptosomal mitoenergetic proteins and Hsp70 early in a rat model of chronic cerebrovascular hypoperfusion. Neurol. Sci., 34: 471-477.
CrossRef