
S.N. Fatihah
Institute of Tropical Aquaculture, Universiti Malaysia Terengganu, 21030, Kuala Terengganu, Terengganu, Malaysia
J. Safiah
Institute of Tropical Aquaculture, Universiti Malaysia Terengganu, 21030, Kuala Terengganu, Terengganu, Malaysia
A.B. Abol-Munafi
School of Fisheries and Aquaculture Sciences, Universiti Malaysia Terengganu, 21030, Kuala Terengganu, Malaysia
M. Ikhwanuddin
Institute of Tropical Aquaculture, Universiti Malaysia Terengganu, 21030, Kuala Terengganu, Terengganu, Malaysia
Pakistan Journal of Biological Sciences
Year: 2014 | Volume: 17 | Issue: 7 | Page No.: 937-941







ABSTRACT
The present study aimed to determine the effect of testosterone undecanoate hormone on sperm quality (sperm viability) and sperm quantity (sperm counts) and its levels in the hemolymph of male mud spiny lobster, Panulirus polyphagus. Male P. polyphagus was injected laterally in fifth abdominal segment of pure hormone, Testosterone Undecanoate (TU) and ethanol at days 1, 8, 15, 22 and 29. Hemolymph of P. polyphagus was taken every two weeks and checked with Enzyme-linked immunosorbent assay (ELISA) to measure hormone levels. The mean sperm quality and quantity were increased due to increase the TU dose and TU levels also increase. The sperm quality, quantity and hormone levels were relevance each others. These findings indicate that TU injection should be evaluated as a practical way of improving sperm quality and quantity in commercial operations.
PDF Abstract XML References Citation
Received: September 27, 2013;
Accepted: January 24, 2014;
Published: March 29, 2014
How to cite this article
S.N. Fatihah, J. Safiah, A.B. Abol-Munafi and M. Ikhwanuddin, 2014. Effect of Testosterone Undecanoate Hormone on Sperm and its Level in the Hemolymph of Male Mud Spiny Lobster, Panulirus polyphagus. Pakistan Journal of Biological Sciences, 17: 937-941.
DOI: 10.3923/pjbs.2014.937.941
URL: https://scialert.net/abstract/?doi=pjbs.2014.937.941
DOI: 10.3923/pjbs.2014.937.941
URL: https://scialert.net/abstract/?doi=pjbs.2014.937.941
INTRODUCTION
Spiny lobster, Panulirus spp., is one of the most important seafood products with high economic value and high prices in various international markets. In Malaysia, the culture of spiny lobsters is still in its early years with activities based in the State of Sabah mostly concentrated in Lahad Datu, Semporna, Kudat and Kota Kinabalu. Spiny lobster culture occupied both grow-out of juveniles in marine cages and hold of adults in cement tanks and marine cages (Tuan and Mao, 2004). The problems in spiny lobster including mud spiny lobster, P. polyphagus breeding technology are low survival rate in the larvae rearing of pueruli stage, difficult to maintain from pueruli until juvenile stages and very difficult to obtained the mature broodstock especially the male P. polyphagus. The availability of the mature males is important for induced mating trials in captivity or further trials on artificial insemination using ready sperm of P. polyphagus.
A number of steroids such as progesterone, 17β-hydroxyprogesterone, 6α-hydroxyprogesterone, 20β-hydroxyprogesterone, pregnenolone, 17β-hydroxypregnenolone, 17α-estradiol, estrone, testosterone and others have been well-known in diverse crustaceans (Fingerman et al., 1993; Subramoniam, 2000; Tsukimura, 2001; Wilder et al., 2002; Oetken et al., 2004). In crustaceans, steroid hormones levels have been studied and reported for many species such as the American lobster, Homarus americanus (Burns et al., 1984; Couch et al., 1987), European Green crab Cacinus mianus (Hazel, 1986) and the black tiger shrimp Penaeus monodon (Fairs et al., 1990; Yashiro and Chaisriha, 1991). The existence of vertebrate-type steroids has been recognized in almost all invertebrate groups including crustaceans (Lehoux and Sandor, 1970; Lafont, 1991; Cardoso et al., 1997).
Testosterone which a single sex differentiating hormones of vertebrates is commonly initiates in the hemolymph and tissues of decapods crustaceans (Ollevier et al., 1986; Fairs et al., 1989; Mackevieciene and Chibisova, 1995). Even though testosterone is not the major factor dependable for sex differentiation in decapods but, it was reported to stimulate growth of male reproductive organs (Nagabhushanam and Kulkarni, 1981), to alter sex ratios towards males (Baghel et al., 2004) and still to transform ovaries into testes (Sarojini, 1963). Therefore, there is a chance for use Testosterone Undecanoate (TU) for enhancing sperm in mature male P. polyphagus. The objectives of the present study were to determine the effect of TU on the sperm quality and quantity and TU levels in the hemolymph of P. polyphagus.
MATERIALS AND METHODS
Fifteen healthy and sexually matured P. polyphagus males were used in the experiment with 10 males for TU and 5 males for control treatments. One week acclimatization was adopted in order to reduce stress associated with handling and laboratory environment. The brood stocks were fed daily at 10% biomass with fresh squid, Loligo sp.
Two fiberglass tanks (5 tonnes capacity) were used for the TU treatments with dose of 0.01 μg g-1 body weight (BW) (5 males) and 0.1 μg g-1 body weight (BW) (five males). One fiberglass tank (5 tonnes capacity) was also used for control treatment (5 males). The hormone dose at 0.01 and 0.1 μg g-1 BW used was modified from Alfaro (1996). Male P. polyphagus was injected laterally in fifth abdominal segment with 0.5 mL dilution of pure hormone TU and ethanol (control) at days 1, 8, 15, 22, 29 and 39. Day 42, the male P. polyphagus was disected to check sperm quality and quantity and before disect, hemolymph from male P. polyphagus was taken. The sperm quality and quantity of P. polyphagus at day 42 were determined using the percentage of viable sperm (sperm viability) and sperm count (sperm number), respectively. Percentages of viable sperm (sperm viability) was known when a live and death of sperm were counted. Percentages of viable sperm were calculated by following equation:
Hemolymph of P. polyphagus was taken every 2 weeks and checked with Enzyme-linked immunosorbent assay (ELISA) (Cusabio Biotech Co. Ltd.,) to measure hormone level. For the ELISA assays, the microtiter plate provided in the kit has been pre-coated with goat-anti-rabbit antibody. Standards or samples were added to the appropriate microtitre plate wells with a HRP-conjugated testosterone and antibody preparation specific for testosterone and incubated. The substrate solutions were then added to each well. The enzyme-substrate reaction was accomplished by the addition of a sulphuric acid solution and the color change was measured spectrophotometically at a wavelength of 450±2 nm. The concentration of testosterone in the samples was determined by comparing the Optical Density (OD) of the samples to the standard curve (Cusabio Biotech Co. Ltd.).
One way ANOVA (Bonferroni) was used for the statistical evaluation of mean values. Significance was accepted at p<0.05.
RESULTS
The result for mean sperm viability percentage were 72.11±5.38% in control, 91.84±1.95% in dose 0.01 μg g-1 BW and 92.14±3.56% in dose 0.1 μg g-1 BW (Fig. 1). The mean sperm counts were 38.84x106 cells mL-1 (control), 62.64x106 cells mL-1 (dose 0.01 μg g-1 BW) and 102.83x106 cells mL-1 (dose 0.1 μg g-1 BW) (Fig. 2). There was significant difference between each dose (p<0.05) for the mean sperm quantity (sperm count).
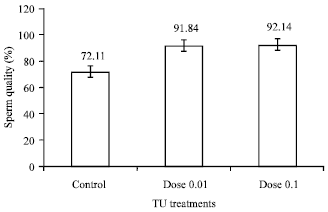 | |
| Fig. 1: | Mean percentage of sperm viability (sperm quality) of P. polyphagus treated with difference dose of testosterone undecanoate (TU) for 42 days study period |
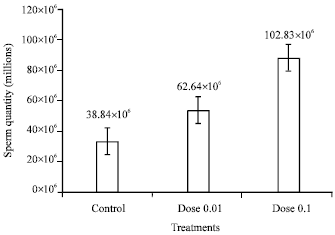 | |
| Fig. 2: | Mean percentage of sperm counts (sperm quantity) of P. polyphagus treated with different dose of testosterone undecanoate (TU) for 42 days study period |
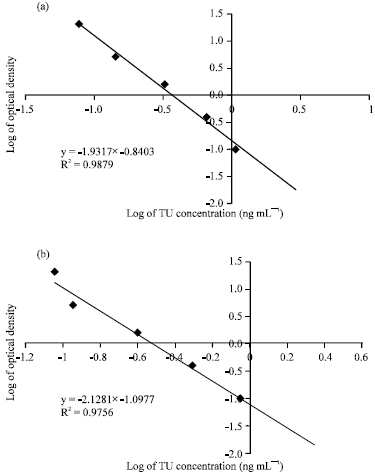 | |
| Fig. 3(a-b): | ELISA standard curve of TU concentration (ng mL-1) |
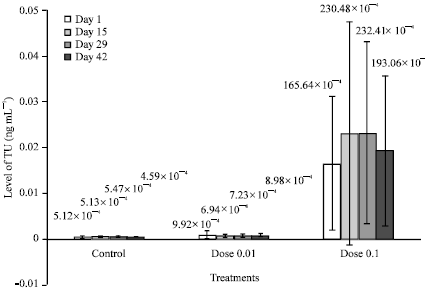 | |
| Fig. 4: | Mean testosterone undecanoate (TU) levels in control, dose 0.01 and 0.1 μg g-1 BW in days 1, 15, 29 and 42 of P. polyphagus |
The mean sperm counts in dose 0.1 μg g-1 BW was very higher than the other two treatments of control and dose 0.01 μg g-1 BW. The standard curves of TU were obtained (Fig. 3) before the TU levels in the hemolymph were determined. In the hemolymph, TU levels in control were low compared to the treatment that injected with 0.01 and 0.1 μg g-1 BW. The mean TU levels of control were 5.12x10-4, 5.13x10-4, 5.47x10-4 and 4.59x10-4 ng mL-1 for days 1, 15, 29 and 42. The means of TU levels of treatments for days 1, 15, 29 and 42 were 9.92x10-4, 6.94x10-4, 7.23x10-4 and 8.98x10-4 ng mL-1, respectively for 0.01 μg g-1 BW dose and 165.64x10-4, 230.48x10-4, 232.41x10-4 and 193.06x10-4 ng mL-1 respectively for 0.1 μg g-1 BW dose (Fig. 4). The mean TU concentration for control and dose 0.1 μg g-1 BW was increased in day 15 and 29 and decreased in day 42 (Fig. 4). But, in dose 0.01 μg g-1 BW, the mean TU concentration was decreased in day 15 and increased in day 29 and 42 (Fig. 4).
DISCUSSION
The mean sperm quality (sperm viability) had increase after been injected with 0.01 and 0.1 μg g-1 BW compared to the control. The sperm quantity (sperm counts) in control treatment was lower than the dose 0.01 and 0.1 μg g-1 BW. Therefore, the sperm quantity was an improvement by injecting of TU. There was significant difference (p<0.05) in sperm quantity in dose 0.01 and 0.1 μg g-1 BW from control treatment with significant improvement in injection of TU in dose 0.1 than dose 0.01 μg g-1 BW.
In the present study, TU levels of male P. polyphagus measured using ELISA. There are a number of different techniques have been well-known to establish vitellogenin (VTG) levels in crustaceans including immunodiffusion (Quinitio et al., 1990; Tom et al., 1987), rocket immunoelectrophoresis (Yano, 1987) and ELISA (Sagi et al., 1999; Tsukimura et al., 2000; Vincent et al., 2001). In ELISA, there were numerous advantages in terms of wellbeing, simplicity and speed and thus, is a method of choice for testing at hatcheries or on site (Mendoza et al., 1993). One more advantage of the ELISA is that hemolymph samples can be assayed directly, not including extraction or hormone enrichment procedures (Chang et al., 1998).
TU has significant effect on hormone level in P. polyphagus. There was no significant effect on hemolymph vitelogenin (Vg) levels of ridgeback shrimp, Sicyonia ingentis when injection of progesterone, 17β-HP and 17α-estradiol (Tsukimura et al., 2000) were given or used. There were difference between TU level in control and dose 0.01 with dose 0.1 μg g-1 BW. For the control treatment, TU level lower than dose 0.01 and 0.1 μg g-1 BW because TU level was naturally present in the P. polyphagus body with no other source of artificial injection. For dose 0.1 μg g-1 BW, TU levels increase dramatically because dose 0.1 μg g-1 BW was enough to stimulate TU level in the body P. polyphagus and thus, the sperm quantity was increased. The TU levels and sperm quantity was an ascendancy each others.
CONCLUSION
The present study shows that TU has affected the sperm quality (sperm viability) and sperm quantity (sperm counts) and hormone levels in mud spiny lobster, P. polyphagus. The injection of dose 0.1 μg g-1 BW was higher and produced significant different (p<0.05) as compared to control and dose 0.01 μg g-1 BW in mean TU levels.
ACKNOWLEDGMENTS
This research was funded by the Malaysia’s Ministry of Education under Exploratory Research Grant Scheme (ERGS) (Vot. No. 55051). Our great appreciation to Institute of Tropical Aquaculture, Universiti Malaysia Terengganu and all who involved directly or indirectly during this study.
REFERENCES
- Alfaro, J., 1996. Effect of 17α-methyltestosterone and 17α-hydroxyprogesterone on the quality of white shrimp Penaeus vannamei spermatophores. J. World Aquacult. Soc., 27: 487-492.
CrossRefDirect Link - Baghel, D.S., W.S. Lakra and G.P. Satyanarayana Rao, 2004. Altered sex ratio in giant fresh water prawn, Macrobrachium rosenbergii (de Man) using hormone bioencapsulated live Artemia feed. Aquacult. Res., 35: 943-947.
CrossRef - Burns, B.G., G.B. Sangalang, H.C. Freeman and M. McMenemy, 1984. Isolation and identification of testosterone from the serum and testes of the American lobster (Homarus americanus). Gen. Comp. Endocrinol., 54: 429-432.
CrossRef - Cardoso, A.M., C.M. Barros, A.J.F. Correia, J.M. Cardoso, A. Cortez, F. Carvalho and L. Baldaia, 1997. Identification of vertebrate type steroid hormones in the shrimp Penaeus japonicus by tandem mass spectrometry and sequential product ion scanning. J. Am. Soc. Mass Spectrom., 8: 365-370.
CrossRefDirect Link - Couch, E.F., N. Hagino and J.W. Lee, 1987. Changes in estradiol and progesterone immunoreactivity in tissues of the lobster, Homarus americanus, with developing and immature ovaries. Comp. Biochem. Physiol. Part A: Physiol., 87: 765-770.
CrossRefDirect Link - Chang, E.S., R. Keller and S.A. Chang, 1998. Quantification of crustacean hyperglycemic hormone by ELISA in hemolymph of the lobster, Homarus americanus, following various stresses. Gen. Comp. Endocrinol., 111: 359-366.
CrossRefDirect Link - Fairs, N.J., R.P. Evershed, P.T. Quinlan and L.J. Goad, 1989. Detection of unconjugated and conjugated steroids in the ovary, eggs and haemolymph of the decapod crustacean Nephrops norvegicus. Gen. Comp. Endocrinol., 74: 199-208.
CrossRef - Fairs, N.J., P.T. Quinlan and L.J. Goad, 1990. Changes in ovarian unconjugated and conjugated steroid titers during vitellogenesis in Penaeus monodon. Aquaculture, 89: 83-99.
CrossRef - Fingerman, M., R. Nagabhushanam and R. Sarojini, 1993. Vertebrate-type hormones in crustaceans: Localization, identification and functional significance. Zool. Sci., 10: 13-29.
Direct Link - Lehoux, J.G. and T. Sandor, 1970. The occurrence of steroids and steroid metabolizing enzyme systems in invertebrates: A review. Steroids, 16: 141-171.
CrossRefDirect Link - Lafont, R., 1991. Reverse endocrinology, or hormones seeking functions. Insect Biochem., 21: 697-721.
CrossRefDirect Link - Mendoza, R., J.C. Guillaume and C. Fauvel, 1993. Homologous ELISA procedure for the determination of penaeid shrimp vitellogenin. Aquat. Living Resour., 6: 39-48.
Direct Link - Nagabhushanam, R. and G.K. Kulkarni, 1981. Effect of exogenous testosterone on the androgenic gland and testis of a marine penaeid prawn, Parapenaeopsis hardwickii(Miers)(Crustacea, Decapoda, Penaeidae). Aquaculture, 23: 19-27.
CrossRef - Ollevier, F., D. De Clerck, H. Diederik and A. De Loof, 1986. Identification of nonecdysteroid steroids in hemolymph of both male and female Astacus leptodactylus(Crustacea) by gas chromatography-mass spectrometry. Gen. Comp. Endocrinol., 61: 214-228.
CrossRef - Oetken, M., J. Bachmann, U. Schulte-Oehlmann and J. Oehlmann, 2004. Evidence for endocrine disruption in invertebrates. Int. Rev. Cytol., 236: 1-44.
CrossRef - Quinitio, E.T., A. Hara, K. Yamauchi and A. Fuji, 1990. Isolation and characterization of vitellin from the ovary of Penaeus monodon. Invertebrate Reprod. Dev., 17: 221-227.
CrossRefDirect Link - Sagi, A., I. Khalaila, U. Abdu, R. Shoukrun and S. Weil, 1999. A newly established ELISA showing the effect of the androgenic gland on secondary-vitellogenic-specific protein in the hemolymph of the crayfish Cherax quadricarinatus. Gen. Comp. Endocrinol., 115: 37-45.
CrossRefDirect Link - Subramoniam, T., 2000. Crustacean ecdysteriods in reproduction and embryogenesis. Comp. Biochem. Physiol. C: Pharmacol. Toxicol. Endocrinol., 125: 135-156.
CrossRefDirect Link - Tom, M., M. Goren and M. Ovadia, 1987. Purification and partial characterization of vitellin from the ovaries of Parapenaeus lingirostris (Crustacea, Decapoda, Panaeidae). Comp. Biochem. Physiol., 87: 17-23.
CrossRef - Tsukimura, B., J.S. Bender and C.J. Linder, 2000. Development of an anti-vitellin ELISA for the assessment of reproduction in the ridgeback shrimp, Sicyonia ingentis. Comp. Biochem. Physiol. A: Mol. Integr. Physiol., 127: 215-224.
CrossRefDirect Link - Tsukimura, B., 2001. Crustacean vitellogenesis: Its role in oocyte development. Am. Zool., 41: 465-476.
CrossRefDirect Link - Vincent, S.G.P., R. Keller and T. Subramoniam, 2001. Development of vitellogenin-ELISA, an in vivo bioassay and identification of two vitellogenesis-inhibiting hormones of the tiger shrimp Penaeus monodon. Mar. Biotechnol., 3: 561-571.
CrossRefDirect Link - Yano, I., 1987. Effect of 17α-hydroxy-progesterone on vitellogenin secretion in kuruma prawn, Penaeus japonicus. Aquaculture, 61: 49-57.
CrossRefDirect Link