
Nina Artanti
Research Center for Chemistry, Indonesian Institute of Sciences, Kawasan PUSPIPTEK, 15314, Serpong, Indonesia
Sanro Tachibana
Faculty of Agriculture Ehime University, 3-5-7 Tarumi, Matsuyama, 790-8566, Ehime, Japan
Leonardus B.S. Kardono
Research Center for Chemistry, Indonesian Institute of Sciences, Kawasan PUSPIPTEK, 15314, Serpong, Indonesia
Pakistan Journal of Biological Sciences
Year: 2014 | Volume: 17 | Issue: 7 | Page No.: 884-890







ABSTRACT
The active α-glucosidase inhibitor compounds in the endophytic fungus Colletotrichum sp. TSC13 were found to be the unsaturated fatty acids (oleic, linoleic and linolenic acids). These compounds have potential as antidiabetic agents. The aim of the present study is to investigate the effects of various media composition on growth (mycelium dry weight) and the fatty acids content (μg mg-1 mycelium DW) of Colletotrichum sp. TSC13 in relation to its α-glucosidase inhibitory activity. For that purpose, the experiments were set up by varying the carbon and nitrogen sources, metal ions and desaturase and fatty acid synthase inhibitors in the media. Colletotrichum sp. TSC13 grown on potato dextrose broth (PDB) was used as control. The α-glucosidase inhibitory activities were (range from 43.9±2.5 to 88.6±5.2%) at 10 μg mL-1. This activity seemed to correlate with the unsaturated fatty acids content of the samples. Different sugars as carbon source experiment showed that xylose gave the highest growth (938.7±141.6 mg). However, the highest fatty acids content was obtained from fructose medium which containing linoleic acid (38.8±4.9 μg mg-1 DW). Soluble starch gave better growth (672.5±62.3 mg) but very low fatty acids content (2.8±0.1 μg mg-1 DW) was obtained. Yeast extract was the best nitrogen source. Fatty acids production was better as compared to beef extract and soytone. This is the first report of various media compositions on fatty acids content in Colletotrichum sp. TSC13 in relation to its α-glucosidase inhibitory activity.
PDF Abstract XML References Citation
Received: July 23, 2013;
Accepted: January 06, 2014;
Published: February 12, 2014
How to cite this article
Nina Artanti, Sanro Tachibana and Leonardus B.S. Kardono, 2014. Effect of Media Compositions on α-glucosidase Inhibitory Activity,
Growth and Fatty Acid Content in Mycelium Extracts of Colletotrichum sp. TSC13 from Taxus Sumatrana (Miq.) de Laub. Pakistan Journal of Biological Sciences, 17: 884-890.
DOI: 10.3923/pjbs.2014.884.890
URL: https://scialert.net/abstract/?doi=pjbs.2014.884.890
DOI: 10.3923/pjbs.2014.884.890
URL: https://scialert.net/abstract/?doi=pjbs.2014.884.890
INTRODUCTION
Colletotrichum sp. TSC13 is an endophytic fungi isolated from Taxus sumatrana. The active α-glucosidase inhibitor compounds in this endophytic fungus were found to be the unsaturated fatty acids (oleic, linoleic and linolenic acids) (Artanti et al., 2012). These compounds may have potential for the development of antidiabetic agents (Artanti et al., 2012). Fatty acid contents in microbial cultures were shown to be affected by media compositions such as the carbon source, nitrogen source, metal ions and precursors or inhibitors of fatty acid biosynthesis (Certik and Shimizu, 1999; Aki et al., 2001; Dyal et al., 2005).
Manipulating the fatty acid content in a culture can be achieved by creating desaturase defective microorganisms (Certik et al., 1998) or the application of desaturase inhibitors (Certik and Shimizu, 1999). Kapok seeds were previously show to contain sterculic acid, a desaturase inhibitor from stearic to oleic acid (Moreton, 1985; Certik and Shimizu, 1999). Other fatty acid desaturase inhibitors were the non-oil fractions of sesame and curcumin from turmeric (Certik and Shimizu, 1999). Green tea extract w as also shown to be a mammalian fatty acid synthase inhibitor (Wang and Tian, 2001; Puig et al., 2008). Although mammalian and fungal fatty acid synthases are different, the presence of green tea extract in the fungal media may have an effect on the fatty acids conten produced by the cultures. In present study, investigation on the effects of the carbon source, nitrogen source, metal ions and desaturase and fatty acid synthase inhibitors on growth and the fatty acids content of Colletotrichum sp. TSC13 in relation to its α-glucosidase inhibitory activity were reported.
MATERIALS AND METHODS
This research was conducted at the Faculty of Agriculture, Ehime University, Japan and Research Center for Chemistry, Indonesian Institute of Sciences (LIPI), Indonesia in 2012-2013.
Fungi and culture conditions: The endophytic fungus, Colletotrichum sp. TSC13 from Taxus sumatrana (Miq.) de Laub. used in this study was kindly provided by Ms. Harmastini Sukiman (Research Center for Biotechnology, Indonesian Institute of Sciences). Maintenance of this culture was described in a previous study (Artanti et al., 2011, 2012). All experiments were conducted in a 500 mL Erlenmeyer-flask containing 150 mL of medium. Colletotrichum sp. TSC13 grown on PDB (potato dextrose broth) (Artanti et al., 2012) was used as a control medium. The flasks were incubated at 25°C with shaking at 75 rpm for seven days. Mycelia were harvested by being filtered through Whatman filtered study, washed with water and dried until a constant dry weight. The mycelium dry weight was measured as growth and dried mycelia were extracted with methanol.
Effect of the addition of different sugars: The control medium was PDB (potato dextrose broth) containing 4 g L-1 potato extract and 20 g L-1 dextrose (glucose). The 20 g L-1 each of fructose, sucrose and xylose was added to the media to replace glucose and their effects on α-glucosidase inhibitory activity, growth and fatty acid content were determined.
Effect of the addition of soluble starch as a carbon source: The control medium was PDB (potato dextrose broth) containing 4 g L-1 potato extract and 20 g L-1 dextrose (glucose). Soluble starch 20 g L-1 was added to the media and its effect on α-glucosidase inhibitory activity, growth and fatty acid content was determined.
Effect of the addition of different nitrogen sources: The control medium was PDB (potato dextrose broth) containing 4 g L-1 potato extract and 20 g L-1 dextrose (glucose). A total of 1.5 g L-1 each of beef extract, soytone and yeast extract were added to the PDB media and their effects on α-glucosidase inhibitory activity, growth and fatty acid content were determined.
Effect of the addition of different metal Ions: The control medium was PDB (potato dextrose broth) containing 4 g L-1 potato extract and 20 g L-1 dextrose (glucose). Five mg L-1 each of CuSO4.5H2O, FeSO4.7H2O and Mg SO4 were added to the PDB media and their effects on α-glucosidase inhibitory activity, growth and fatty acid content were determined.
Effect of the the adition of inhibitors: He control medium was PDB (potato dextrose broth) containing 4 g L-1 potato extract and 20 g L-1 dextrose (glucose).. Kapok water extract (filtrate of 10g L-1 Kapok), 2 g L-1 sesame oil, turmeric water extract (filtrate of 30g L-1 turmeric) and green tea extract (commercial non-sugar bottled green tea) were each added to the PDB media and their effects on α-glucosidase inhibitory activity. The growth and fatty acid content were determined.
Analytical methods: Samples were methylated to form Fatty Acid Methyl Esters (FAME) to determine the fatty acid content, as described in a previous report (Artanti et al., 2012) using GC Shimadzu-QP-2014 equipped with a SPB-50 column (30 m x 0.25 mm I.D., 0.25 μm film thickness). Quantification of the FAME content in samples was conducted by comparing the peak area of samples with the standard curves of FAME standards.
Alpha-glucosidase inhibitory assay: The α-glucosidase inhibitory assay was conducted as described in a previous report (Artanti et al., 2012).
Data analysis: Data analysis was performed using Microsoft Office Excel 2003. Values are shown as the average of duplicate measurements±standard deviation.
RESULTS AND DISCUSSION
Effect of the addition of different sugars: Figure 1 shows the effects of the different sugars type used in the media on growth and fatty acid content in the Colletotrichum sp. TSC13 mycelium extract.
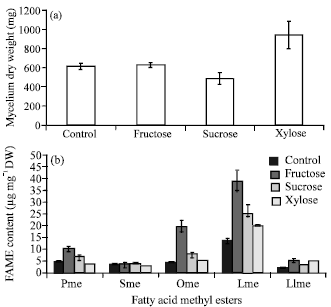 | |
| Fig. 1(a-b): | Effect of the addition of different sugars on (a) Growth and (b) Fatty acid content, Pme: Palmitic acid methyl ester; Sme: Stearic acid methyl ester; Ome: Oleic acid methyl ester; Lme: Linoleic acid methyl ester; Llme: Linolenic acid methyl ester. Values are shown as the average of duplicate measurements±standard deviation |
The highest mycelium dry weight (938.7±141.6 mg) was obtained when xylose was used. Similar growth was obtained when glucose and fructose were used, whereas the lowest growth was obtain with sucrose. The effects of glucose, fructose and sucrose on growth were consistent with the effects of these sugars on the growth of Mortierella ramanniana var. ramanniana (Dyal et al., 2005). Higher growth with fructose as the carbon source than with glucose was also observed in Colletotrichum gloeosporioides cultures (Sangeetha and Rawal, 2008), while lower growth was observed in Colletotrichum sp. (DM06) and Alternaria sp. (DM09) (Dey et al., 2011). Lower growth with sucrose as the carbon source than with glucose was previously reported in cultures of Mucor sp. 1b (Somashekar et al., 2003), Motrierella alliaceae YN-15 (Aki et al., 2001) and Colletotrichum sp. (DM06) and Alternaria sp. (DM09) (Dey et al., 2011) and Colletotrichum gloeosporioides (Deshmukh et al., 2012). However, higher growth in sucrose media than in the glucose was reported in Motrierella rouxii MTCC 386 (Somashekar et al., 2003) and Colletotrichum gloeosporioides (Sangeetha and Rawal, 2008). Higher growth with xylose as the carbon source than with glucose was also observed in Colletotrichum gloeosporioides cultures (Deshmukh et al., 2012). In contrast, lower growth was observed in cultures of Colletotrichum sp. (DM06) and Alternaria sp. (DM09) (Dey et al., 2011), Mortierella isabellina (Fakas et al., 2009; Ruan et al., 2012) and Pleurotus pulmonarius (Smiderle et al., 2012). The proportion pattern of fatty acid content palmitic acid and stearic acid (saturated fatty acids), oleic acid, linoleic acid and linolenic acid (unsaturated fatty acid)) was similar with different sugars as carbon source, whereas the fatty acid content of linoleic acid was always the highest. The highest linoleic acid content (38.8±4.9 μg g-1 DW) was obtained from fructose media and the lowest (20.4±0.1 μg g-1 DW) was obtained from glucose (control) media. Therefore, the effect of different sugar types on growth did not have a linear correlation with their effects on fatty acid production. In contrast to the results of this study relative to glucose, lower fatty acids contents were observed with fructose and sucrose as carbon source in cultures of Mortierella ramanniana var. ramanniana (Dyal et al., 2005). Colletotrichum sp. (DM06) and Alternaria sp. (DM09) (Dey et al., 2011) also showed lower fatty acid content with xylose as the carbon source. Xylose, exhibited the best effect on growth, however, the fatty acid content in this media were significantly lower than that of fructose but was still higher than of glucose.
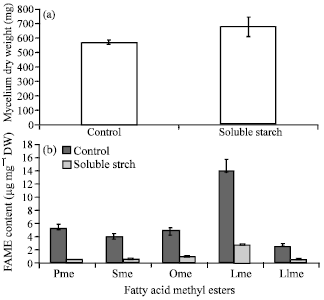 | |
| Fig. 2(a-b): | Effect of the addition of soluble starch on (a) Growth and (b) Fatty acid content, Pme: Palmitic acid methyl ester, Sme: Stearic acid methyl ester, Ome: Oleic acid methyl ester, Lme: Linoleic acid methyl ester, Llme: Linolenic acid methyl ester values are shown as the average of duplicate measurements±standard deviation |
Therefore, any fungal strain may have unique preferences to the type of sugar and the effect of different sugar types on growth does not have a linear correlation with their effects on fatty acid production in the culture. Good growth and fatty acid production also reported on Mortierella isabellina with xylose as the carbon source (Gao et al., 2013).
Effect of the addition of soluble starch: Figure 2 shows the effect of soluble starch when it was used as the carbon source on growth and fatty acid content in Colletotrichum sp. TSC13 mycelium extract. The results showed that compare to control (glucose), soluble starch as the complex carbohydrate gave slightly better growth (19% increased in mycelium DW) but was significantly lower fatty acid production (around 76-90% decreased in fatty acids content). This result differed from that in Mortierella alliaceae YN-15. The growth and fatty acid content of this culture was slightly lower when soluble starch was used than when glucose was used (Aki et al., 2001). Significantly lower fatty acid content in soluble starch media than in glucose media has also reported in Mortierella plumbeus, although two other Mortierella species (M. ramanniana and M. mucedo) showed similar fatty acid contents in both media (Sajbidor et al., 1992; Dyal and Narine, 2005).
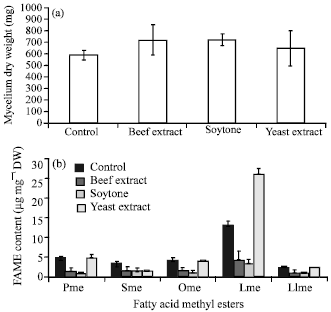 | |
| Fig. 3(a-b): | Effect of the addition of diferent nitrogen sources on (a) Growth and (b) Fatty acid content, Pme: Palmitic acid methyl ester, Sme: Stearic acid methyl ester, Ome: Oleic acid methyl ester, Lme: Linoleic acid methyl ester, Llme: Linolenic acid methyl ester; Values are shown as the average of duplicate measurements±standard deviation |
Significantly higher fatty acid production was observed in soluble starch media than in glucose media and other carbon sources has been reported in M. alpine (Jang et al., 2005). This finding suggest that soluble starch and other carbon sources affect growth and fatty acid content depending on the fungal species.
Effect of the addition of different nitrogen sources: Figure 3 shows the effects of different nitrogen sources on growth and fatty acid content of Colletotrichum sp. TSC13 mycelium extract. The results show that the addition of different types of nitrogen sources from an animal (beef extract), plant (soytone) and microorganism (yeast extract) slightly increased growth (around 10-22% ) relative to the control without the addition of nitrogen source. However, nitrogen sources had a significant effect on fatty acid content. Compare to control, adding beef extract and soytone significantly decreased the content of all fatty acids (around 48-79%). Yeast extract significantly increased linoleic acid content (100%) but only has little effect on palmitic, oleic and linolenic acids and decreased the content of stearic acid. Using yeast extract as nitrogen source to grow fungi is more common than beef extract and soytone.
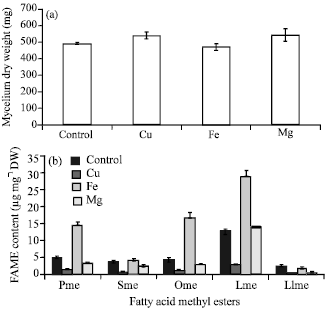 | |
| Fig. 4(a-b): | Effect of the addition of metal ions on (a) Growth and (b) Fatty acid content, Pme: Palmitic acid methyl ester, Sme: Stearic acid methyl ester, Ome: Oleic acid methyl ester, Lme: Linoleic acid methyl ester, Llme: Linolenic acid methyl ester values are shown as the average of duplicate measurements±standard deviation |
Higher fatty acid or lipid contents in the media containing yeast extract were reported in cultures of M. ramanniana var. ramanniana (Dyal et al., 2005) and M. alliaceae YN-15 (Aki et al., 2001). Significant differences in growth and fatty acid with the presence of yeast extract in the cultures of M. alliaceae YN-15 depended on the type of carbon source used (Aki et al., 2001).
Effect of the addition of different metal Ions: Figure 4 shows the effects of metal ion on growth and fatty acid content in the Colletotrichum sp. TSC13 mycelium extract. The results showed that similar growth was obtained in control media and media with the addition of Cu2+, Fe2+ and Mg2+. However, various effects on fatty acid content were observed due to the addition of different metal ions. Adding Cu2+ resulted in a significant reduction in the fatty acid content (around 72-84%). Adding Fe2+ significantly increased the content of palmitic acid (3 fold), oleic acid (4 fold) and linoleic acid (2 fold), however, it decreased the content of linolenic acid (more than half) and had little effect on the content of stearic acid. Adding Mg2+ reduced the fatty acid content but not by as much as that with Cu2+. These results suggested that different metal ions have different effects.
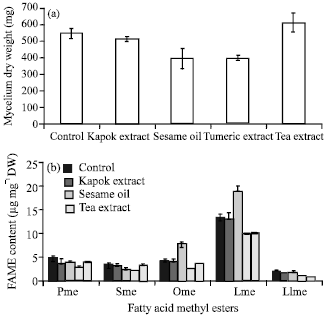 | |
| Fig. 5(a-b): | Effect of the addition of desaturase and fatty acid synthase inhibitors on (a) Growth and (b) Fatty acid content, Pme: Palmitic acid methyl ester, Sme: Stearic acid methyl ester, Ome: Oleic acid methyl ester, Lme: Linoleic acid methyl ester, Llme: Linolenic acid methyl ester values are shown as the average of duplicate measurements±standard deviation |
Lipid accumulation in Cunninghamella sp. 2A1 was shown to be increased in the presence of Mg2+ and Fe2+, while Cu2+ decreased lipid accumulation, especially the content of α-linolenic acid (Muhid et al., 2008). Similar results have also been reported in the culture of Aspergillus niger (Jernejc and Legis, 2002). Different patterns were observed in M. ramanniana var. ramanniana (Dyal et al., 2005) in which the addition of Fe2+ gave the lowest lipid accumulation relative to that of Cu2+ and Mg2+.
Effect of the addition of desaturase and fatty acid synthase inhibitors: Figure 5 shows the effect of desaturase and fatty acid synthase inhibitors on growth and fatty acid content in the Colletotrichum sp. TSC13 mycelium extract. Adding kapok extract had a little effect on growth (only 6% decreased) and stearic, oleic and linoleic acid content (3-10% decreased) to that of the control except slightly bigger effect for reduction on the content of palmitic acid (18%) and linolenic acid (25%). This suggested that kapok extract used in this experiment might be not contain enough sterculic acid to exhibit its effect as a desaturase inhibitor in the conversion of stearic acid to oleic acid. The effect of kapok seed oil containing sterculic acid on modifying fatty acid content was reported in cultures of Candida sp. 107, Trichosporon cutaneum and Rhodosporidium toruloides (Moreton, 1985).
| Table 1: | α-glucosidase inhibitory activity of samples from experiments of various carbon and nitrogen sources, metal ions and desaturase and fatty acid synthase inhibitors in the media. The assay of α-glucosidase inhibitor was conducted at 10 μg mL-1 sample |
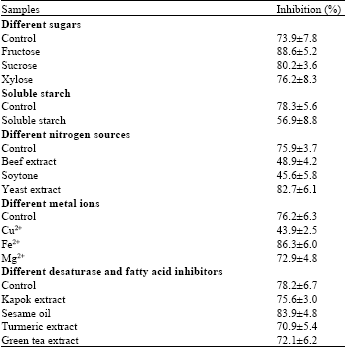 | |
| Values are shown as the average of duplicate measurements±standard deviation | |
Adding sesame oil caused a decrease in growth (28%) and the content of palmitic acid (16%), stearic acid (30%) and linolenic acid (15%), however, this treatment increased the content of oleic acid (77%) and linoleic acid (42%). A high content of linoleic acid was previously observed in M. ramanniana var. ramanniana with the addition of sesame oil (Dyal et al., 2005). Adding turmeric extract also decreased growth (27%) as well as the fatty acid content (around 28-48%). Adding tea extract slightly increased growth (12%) but decreased the fatty acids content (around 10-65%). This result suggested that tea extract may also inhibit fatty acid production in this endophytic fungi culture, although fungal fatty acid synthase differs from mammalian fatty acid synthase.
Effect of various media composition treatments on α-glucosidase inhibitory activity of Colletotrichum sp. TSC13 mycelium extracts: Table 1 shows that α-glucosidase inhibitor activities ranged from 43.9±2.5 to 88.6±5.2% in a 10 μg mL-1 sample. Samples with high unsaturated fatty acids content such as samples from fructose, yeast and Fe2+ media, showed higher α-glucosidase inhibitory activity than samples with a lower unsaturated fatty acids content such as samples from soluble starch media. This suggested that α-glucosidase inhibitory activity appeared to correlate well with the unsaturated fatty acid content of the samples.
CONCLUSION
Various compositions of media resulted in different effects on growth, fatty acid content and α-glucosidase inhibitory activity in endophytic fungi Colletotrichum sp. TSC13. Xylose as the carbon source gave the highest growth, whilst fructose as the carbon source gave the highest fatty acid content. Yeast extract as the microbial nitrogen source had the best effect on fatty acid production. Fe2+ was the best metal ion for fatty acid production although growth was slightly lower than that with Cu2+ and Mg2+. Adding of sesame oil significantly increased the content of oleic and linoleic acid. Green tea extract decreased fatty acid content which indicated that this extract may inhibit fungal fatty acid synthase. This is the first report to show the effect of various media compositions on growth and fatty acid content in relation to the α-glucosidase inhibitory activity of Colletotrichum sp. TSC13.
ACKNOWLEDGMENTS
The authors would like to thank JSPS (The Japan Society for the Promotion of Science) under the Ronpaku Dissertation Ph.D. program for providing fellowship during Fiscal Year 2012 to conduct research in the Faculty of Agriculture, Ehime University, Japan and the Research Center for Chemistry - LIPI, Indonesia.
REFERENCES
- Aki, T., Y. Nagahata, K. Ishihara, Y. Tanaka and T. Morinaga et al., 2001. Production of arachidonic acid by filamentous fungus, Mortierella alliacea strain YN-15. J. Am. Oil Chem. Soc., 78: 599-604.
CrossRef - Artanti, N., S. Tachibana, L.B.S. Kardono and H. Sukiman, 2011. Screening of endophytic fungi having ability for antioxidative and α-glucosidase inhibitor activities Isolated from Taxus sumatrana. Pak. J. Biol. Sci., 14: 1019-1023.
CrossRefDirect Link - Artanti, N., S. Tachibana, L.B.S. Kardono and H. Sukiman, 2012. Isolation of α-glucosidase inhibitors produced by an endophytic fungus, Colletotrichum sp. TSC13 from Taxus sumatrana. Pak. J. Biol. Sci., 15: 673-679.
CrossRefDirect Link - Certik, M., E. Sakuradani and S. Shimizu, 1998. Desaturase-defective fungal mutants: Useful tools for the regulation and overproduction of polyunsaturated fatty acids. Trends Biotechnol., 16: 500-505.
CrossRef - Certik, M. and S. Shimizu, 1999. Biosynthesis and regulation of microbial polyunsaturated fatty acid production. J. Biosci. Bioeng., 87: 1-14.
CrossRefPubMedDirect Link - Deshmukh, B., P. Mehta, A.N. Sabalpara and V.A. Patil, 2012. In vitro effect of various nitrogen, carbon sources and pH regimes on the growth and sporulation of Colletotrichum gloeosporioides Penz. and Sacc causing anthracnose of Indian bean. J. Biopest, 5: 46-49.
Direct Link - Dyal, S.D. and S.S. Narine, 2005. Implications for the use of Mortierella fungi in the industrial production of essential fatty acids. Food Res. Int., 38: 445-467.
CrossRefDirect Link - Dyal, S.D., L. Bouzidi and S.S. Narine, 2005. Maximizing the production of γ-linolenic acid in Mortierella ramanniana var. ramanniana as a function of pH, temperature and carbon source, nitrogen source, metal ions and oil supplementation. Food Res. Int., 38: 815-829.
CrossRefDirect Link - Fakas, S., S. Papanikolaou, A. Batsos, M. Galiotou-Panayotou, A. Mallouchos and G. Aggelis, 2009. Evaluating renewable carbon sources as substrates for single cell oil production by Cunninghamella echinulata and Mortierella isabellina. Biomass Bioenergy, 33: 573-580.
CrossRefDirect Link - Gao, D., J. Zeng, Y. Zheng, X. Yu and S. Chen, 2013. Microbial lipid production from xylose by Mortierella isabellina. Bioresour. Technol., 133: 315-321.
CrossRef - Jang, H.D., Y.Y. Lin and S.S. Yang, 2005. Effect of culture media and conditions on polyunsaturated fatty acids production by Mortierella alpina. Bioresour. Technol., 96: 1633-1644.
CrossRef - Jernejc, K. and M. Legis, 2002. The influence of metal ions on malic enzyme activity and lipid synthesis in Aspergillus niger. FEMS Microbiol. Lett., 217: 185-190.
CrossRef - Moreton, R.S., 1985. Modification of fatty acid composition of lipid accumulating yeasts with cyclopropene fatty acid desaturase inhibitors. Applied Microbiol. Biotechnol., 22: 42-45.
CrossRefDirect Link - Puig, T., J. Relat, P.F. Marrero, D. Haro, J. Brunet and R. Colomer, 2008. Green tea catechin inhibits fatty acid synthase without stimulating carnitine palmitoyltransferase-1 or inducing weight loss in experimental animals. Anticancer Res., 28: 3671-3676.
Direct Link - Ruan, Z., M. Zanotti, X. Wang, C. Ducey and Y. Liu, 2012. Evaluation of lipid accumulation from lignocellulosic sugars by Mortierella isabellina for biodiesel production. Bioresour. Technol., 110: 198-205.
CrossRef - Sajbidor, J., D. Kozelouhov'a and M. Cert'ik, 1992. Influence of some metal ions on the lipid content and arachidonic acid production by Mortierella sp. Folia Microbiologica, 37: 404-406.
CrossRef - Smiderle, F.R., L.M. Olsen, A.C. Ruthes, P.A. Czelusniak and A.P. Santana-Filho et al., 2012. Exopolysaccharides, proteins and lipids in Pleurotus pulmonarius submerged culture using different carbon sources. Carbohydr. Polym., 87: 368-376.
CrossRef - Somashekar, D., G. Venkateshwaran, K. Sambaiah and B.R. Lokesh, 2003. Effect of culture conditions on lipid and γ-linolenic acid production by mucoraceous fungi. Process Biochem., 38: 1719-1724.
CrossRefDirect Link - Wang, X and W. Tian, 2001. Green tea epigallocatechin gallate: A natural inhibitor of fatty-acid synthase. Biochem. Biophys. Res. Commun., 288: 1200-1206.
CrossRef