
Shabnam Nadjafi
Faculty of Medicine, Iran University of Medical Sciences, Tehran, Iran
Soltan-Ahmad Ebrahimi
Faculty of Medicine, Iran University of Medical Sciences, Tehran, Iran
Nahid Rahbar- Roshandel
Faculty of Medicine, Iran University of Medical Sciences, Tehran, Iran
Pakistan Journal of Biological Sciences
Year: 2014 | Volume: 17 | Issue: 11 | Page No.: 1185-1189







ABSTRACT
In this study, the effect of berberine, an isoquinoline alkaloid isolated from Coptidis rhizoma, on Nitric Oxide (NO) production, as a possible involved factor, during excitotoxic injury in oligodendroglial cells were evaluated. The overactivation of ionotropic glutamate receptors which is known as the excitotoxicity, is an important phenomenon because of the contribution in acute injury to the central nervous system, chronic neurodegenerative disorders, oligodendrocyte loss and demyelinating diseases as Multiple Sclerosis (MS). Intracellular Ca2+ overload, have a key role during excitotoxic injury and such increase in cytoplasmic Ca2+ triggers a series of events such as production of NO that end to cell death. Previous report showed the protective effects of berberine on ischemic-induced excitotoxic insult in oligodendrocytes. Hereby, we intended to know if the NO production could be associated with oxygen-glucose deprivation/reperfusion-induced excitotoxic damage in oligodendrocyte; moreover, the alteration of NO production could be considered as an involved mechanism for protective effect of berberine in such condition. Therefore, the effect of berberine (2 μM) on NO production during oxygen-glucose deprivation/24 h reperfusion in oligodendrocytes were examined. The OLN-93 cell line (a permanent immature rat oligodendrocyte) was used as a model of oligodendrocyte. Thirty minutes-oxygen-glucose deprivation/24 h reperfusion was used to induce excitotoxicity. NO production was evaluated by Griess method. Our results demonstrated that berberine (2 μM) significantly decreased NO production during 30 min oxygen-glucose deprivation/reperfusion. It seems that blockade of NO production by berberine may also participate in oligodendroglial cell protection against oxygen-glucose deprivation/ reperfusion-induced insult.
PDF Abstract XML References Citation
Received: June 16, 2014;
Accepted: August 23, 2014;
Published: September 17, 2014
How to cite this article
Shabnam Nadjafi, Soltan-Ahmad Ebrahimi and Nahid Rahbar- Roshandel, 2014. Effect of Berberine on Nitric Oxide Production During Oxygen-Glucose Deprivation/Reperfusion in OLN-93 Oligodendrocytes. Pakistan Journal of Biological Sciences, 17: 1185-1189.
DOI: 10.3923/pjbs.2014.1185.1189
URL: https://scialert.net/abstract/?doi=pjbs.2014.1185.1189
DOI: 10.3923/pjbs.2014.1185.1189
URL: https://scialert.net/abstract/?doi=pjbs.2014.1185.1189
INTRODUCTION
Berberine is an isoquinoline alkaloid isolated from Coptidis rhizoma and Cortex phellodendri (Hsu et al., 2013). Recent studies have revealed that berberine has multiple pharmacological activities including hypoglycemia, hypolipidemia, anti-inflammation and anti-oxidation (Hsu et al., 2013; Cui et al., 2009). Moreover, the neuroprotective effects of berberine in Alzheimer’s disease, transient forebrain ischemia and mental depression have been demonstrated by in vivo and in vitro studies (Cui et al., 2009; Hsu et al., 2013; Chai et al., 2014). Also, berberine presents high blood brain barrier permeability (Wang et al., 2005; Hsu et al., 2013). The protective effect of berberine on ischemic/reperfusion-induced excitotoxic injury in OLN-93 oligodendrocyte was recently reported; furthermore, berberine could attenuate the intracellular Ca2+ overload during the short time ischemia (Nadjafi et al., 2014).
Oligodendrocytes, a type of the glial cells in the central nervous system, wrap axons by forming the myelin sheaths which provide electrical insulation around the axon (Agresti et al., 2005; Giaume et al., 2007; Matute, 2010). Oligodendrocyte death results in demyelination, impaired axonal conduction and finally axon death (Matute, 2010).
L-Glutamate is the major excitatory neurotransmitter in the central nervous system and mediates various physiological or pathophysiological actions via activation of its receptor families (Matute et al., 2007; Kew and Kemp, 2005). Ionotropic glutamate receptors are ligand-gated cation channels which are classified in glutamate receptor families (Kew and Kemp, 2005). Notably, glutamate and the ionotropic glutamate receptors are involved in neuronal and glial cells death (Matute et al., 2001, 2007).
Excitotoxic cell death occurs through sustained activation of ionotropic glutamate receptors by glutamate (Matute et al., 2007). Excitotoxicity leads to cellular Ca2+ overload and the signaling cascades which ultimately end to cell death (Giaume et al., 2007; Matute et al., 2007). Excitotoxicity can occur in almost all neurons which express ionotropic glutamate receptors; in addition, this event can cause injury to glial cells including oligodendrocytes (Matute et al., 2007). All three major types of ionotropic glutamate receptors, alpha-amino-3-hydroxy-5-methylisoxazole-4-propionic acid (AMPA), kainate and N-methyl-D-aspartate (NMDA), participate in glutamate receptor-mediated oligodendroglial injury (Kew and Kemp, 2005; Matute et al., 2007). The importance of excitotoxicity is due to the involvement of this phenomenon in acute injury to the central nervous system and in chronic neurodegenerative disorders, also it can cause oligodendrocyte loss and demyelination (Matute et al., 2007; Matute, 2010).
Nitric Oxide (NO), a messenger molecule with dual action, contributes both in normal physiologic and pathophysiologic conditions (Knott and Bossy-Wetzel, 2009). The NO is generated by the Nitric Oxide Synthase (NOS), an enzyme that exists in three forms, neuronal (nNOS), endothelial (eNOS) and inducible (iNOS) (Yao et al., 2010). Also, mtNOS is a post translational modification of nNOS and is present in the mitochondria (Yao et al., 2010).
Impairment of intracellular calcium homeostasis and increase in NO production are the components of multiple adverse effects which resulted from over-stimulation of glutamate receptors during excitotoxicity phenomenon (Wang and Qin, 2010).
In this study, the effect of berberine on NO production during short time oxygen-glucose deprivation/reperfusion-induced injury in OLN-93 oligodendrocytes was evaluated. OLN-93 cell line, derived from spontaneously transformed cells in primary rat brain glial cultures, is used as the model of immature oligodendrocytes for in vitro studies (Richter-Landsberg and Heinrich, 1996; Wilkins et al., 2003).
MATERIALS AND METHODS
Materials: Berberine chloride was obtained from Sigma Chemical Co. (St. Louis, MO, USA). The stock solution was prepared at 1 mmol L¯1 in deionized water and kept at -20°C. Dulbecco’s Modified Eagle’s medium (DMEM) with glutamine, glucose/glutamine-free DMEM, Fetal Bovine Serum (FBS) were purchased from GIBCO. Penicillin-streptomycin, poly-L-lysine, Dizocilpine (MK-801), NG nitro-L-arginine methyl ester (L-NAME) and modified Griess reagent were purchased from Sigma Chemical Co. (St. Louis, MO, USA). All other chemicals are of the analytical grade available from regular commercial sources.
Cell culture: OLN-93 cell line was purchased from Pasteur Institute of Iran (Tehran, Iran). The cells were grown in DMEM medium supplemented with 10% FBS and 100 IU mL¯1 penicillin and100 μg mL¯1 streptomycin in a humidified 95% air and 5% CO2 incubator at 37°C. The cells were sub cultured twice a week by gentle scraping and were cultured on poly-L-lysine-coated 12-well plates at a density of 5<×104 mg L¯1. Culture dishes were coated with poly-L-lysine 24 h before the experiment. Poly-L-lysine (150 000-300 000 MW) was dissolved in deionized water and the plate wells were filled with this solution (20 μg mL¯1). After 5 min standing at room temperature, the solution was aspirated and the plates were left to dry in a laminar flow hood overnight. Cells were used for experiments 24 h after seeding.
Oxygen-glucose deprivation and drug exposure: The Oxygen-Glucose Deprivation (OGD) condition was performed as described previously (Frantseva et al., 1999). Briefly, the culture medium was replaced with glucose/glutamine-free DMEM and cells were exposed to hypoxia for 30 min in a small anaerobic chamber filled with 95% (v/v) N2 and 5% (v/v) CO2 at 37°C. To terminate the oxygen-glucose deprivation, the chamber was opened and the medium was replaced with DMEM and the cultures were then placed in an incubator with 5% CO2 for 24 h. To examine the drug effects, cell cultures were treated 3 h before oxygen-glucose deprivation with berberine (2 μM). This concentration was selected based on the previous study by our group and berberine presented the peak of protective effect at this concentration (Nadjafi et al., 2014). The pre-incubation time of 3 h was selected based upon previous studies (Rahbar-Roshandel et al., 2008). To investigate the effects of the inhibitors on oxygen-glucose deprivation-induced cell death, MK-801, a non-competitive antagonist of the NMDA receptor and L-NMAE, a NOS inhibitor, was added to the medium 3 h before oxygen-glucose deprivation (Tavakoli-Far et al., 2009). Plates not exposed to oxygen-glucose deprivation were used as external control and those exposed to oxygen-glucose deprivation conditions without the addition of any drug were used as internal control (OGD control). Each experiment was repeated at least three times, so every data point is therefore the mean of at least three measurements.
Measurement of nitrite: The level of nitrite as a measure of NO production in the culture medium was quantified using modified Griess reagent. In brief, the medium in each well was removed and centrifuged at 10,000 rpm for 10 min at 4°C.
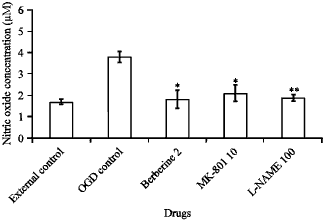 | |
| Fig. 1: | Effect of berberine on Nitric Oxide (NO) production during ischemia/reperfusion-induced injury in OLN-93 oligodendrocytes. The effects of berberine (2 μM), MK-801 (10 μM) and L-NAME (100 μM) were evaluated on NO production during a 30 min Oxygen-Glucose Deprivation (OGD)/24 h reperfusion-induced cell injury in cultured OLN-93 oligodendrocytes. The plates not exposed to oxygen-glucose deprivation were considered as external control and those exposed to oxygen-glucose deprivation conditions without the addition of any drug were used as OGD control. The values are presented as Mean"SE compared with OGD control (*p<0.05; **p<0.01) |
Then, the supernatant were mixed with an equal volume of modified Griess reagent at room temperature for 10 min and the absorbance was measured at 540 nm using an Ultrospec ®3000 UV/Visible spectrophotometer (Pharmacia Biotech, Cambridge, England). The nitrite concentration was determined from a sodium nitrite standard curve.
Statistical analysis: All results are expressed as Mean±SE. The significance of differences was evaluated using Student’s t-test by Excel 2007 (Microsoft; Seattle, WA, USA) software. The p<0.05 was considered statistically significant.
RESULTS
Effects of berberine, L-NAME and MK-801 on nitric oxide production during 30 min oxygen-glucose deprivation/reperfusion in OLN-93 cells: NO production in the culture medium was measured by modified Griess reagent after 30 min oxygen-glucose deprivation/reperfusion. Exposure of OLN-93 cells to 30 min of oxygen-glucose deprivation/reperfusion significantly increased nitrite production compared with the external control (p<0.01). Pretreatment of cultures with berberine (2 μM), significantly decreased NO production, as compared with the oxygen-glucose deprivation control (p<0.05). The effects of L-NAME (100 μM), a NOS inhibitor and MK-801 (10 μM), a non-competitive antagonist of the NMDA receptor, on NO production were also evaluated and both compounds reduced the NO production, significantly (p<0.01; p<0.05, respectively) (Fig. 1).
DISCUSSION
Berberine, an isoquinoline alkaloid, presents anti-apoptotic effects on the neurons suffering ischemia in addition to its other pharmacologic properties (Cui et al., 2009; Zhang et al., 2012; Chai et al., 2014). The previous report showed that berebrine could present protective effects and attenuate the intracellular Ca2+ elevation during oxygen-glucose deprivation/reperfusion-induced injury in OLN-93 oligodendrocytes (Nadjafi et al., 2014).
The results of this study revealed that berberine could attenuate the NO production during a short-term ischemic/reoxygenation-induced injury in OLN-93 oligodendrocytes. Moreover, L-NAME, a NOS inhibitor and MK-801, the non-competitive NMDA receptor antagonist, decreased the NO production through same experiments.
Glutamate-mediated injury to oligodendrocytes contributes to mental or physical impairment in acute and chronic diseases as periventricular leukomalacia (prenatal white matter injury leading to cerebral palsy), spinal cord injury, multiple sclerosis and stroke (Karadottir et al., 2005; Matute et al., 2007). It is known that, oligodendrocytes express Ca2+-permeable ionotropic glutamate receptors including NMDA, AMPA and kainate receptors (Salter and Fern, 2005; Matute, 2006; Wang and Qin, 2010) whereas, AMPA/kainate receptors are expressed on somata and NMDA receptors on processes (Salter and Fern, 2005).
Some evidences demonstrated a direct link between ischemic damage to oligodendrocytes and excitotoxicity (Matute, 2010). Also, the intracellular Ca2+ overload, is the key mediator of the excitotoxic oligodendroglial injury (Matute et al., 2001, 2007; Matute, 2010; Wang and Qin, 2010). The subsequent excitotoxic oligodendroglial death due to overactivation of AMPA and kainate receptors had been demonstrated, earlier (Matute et al., 2001). Now it is well known that, the expressed NMDA receptors in the oligodendrocytes are also activated in ischemia (Karadottir et al., 2005).
In the case of excitotoxicity and/or ischemia, it is considered that there is a relation between intracellular Ca2+ overload and NO production (Szydlowska and Tymianski, 2010; Wang and Qin, 2010). Hereof, calcium ions can enter into the neuronal cell through various mechanisms; these include the over-activation of ionotropic glutamate receptors or of a range of channels and transporters (Szydlowska and Tymianski, 2010). Potentially toxic cytoplasmic calcium concentrations can also occur due to release from intracellular sources (Szydlowska and Tymianski, 2010). Such increases of cytoplasmic calcium concentrations can trigger a range of downstream neurotoxic events such as activation of nitric oxide synthase (Szydlowska and Tymianski, 2010). Notably, in neurons, NMDA receptor is linked to the neurotoxic molecules such as nNOS (Szydlowska and Tymianski, 2010). After ischemia, over-activation of NMDA receptor results in activation of nNOS leads to the production of NO and reactive oxygen species that can end to neuronal cell death (Szydlowska and Tymianski, 2010). Over all, through glutamate-induced neuronal death, NMDA receptor activation leads to increased cytosolic calcium and the intracellular Ca2+ elevation may stimulate nNOS production of NO and peroxynitrite which take part in the events that end to cell death (Brown, 2010; Szydlowska and Tymianski, 2010).
The results of present study showed the attenuation of NO production by MK-801, the non-competitive NMDA receptor antagonist and L-NAME, a NOS inhibitor, in OLN-93 oligodendrocytes during short-term ischemia/reperfusion. Thus, it revealed that NO production might contribute in ischemic-induced excitotoxicity in oligodendrocyte similar to neuron. On this subject, one study has also suggested the involvement of nitric oxide and excitotoxicity in hypoxic damage to the periventricular white matter in rat neonatal brain (Kaur et al., 2006).
Recently, neuroprotective effect of berberine against cerebral ischemia and reperfusion has been demonstrated by many studies (Cui et al., 2009; Hu et al., 2012; Chai et al., 2014). Briefly, the effects of berberine on this condition are due to its anti-oxidative and the anti-apoptotic effects (Hu et al., 2012; Chai et al., 2013). Also, berberine can be neuroprotective against high glucose-induced damage (Hsu et al., 2013). Interestingly, it seems that the facilitation of glucose metabolism by berberine may contribute to the neuroprotective effect of this alkaloid against in vitro ischemia-induced neuronal cell damage in mouse organotypic hippocampal slice cultures (Cui et al., 2009).
CONCLUSION
In conclusion, it seems that reduction of NO production may also be one of the mechanisms by which berberine protects the OLN-93 oligodendrocytes against ischemic/reperfusion-induced excitotoxic injury.
REFERENCES
- Agresti, C., M.E. Meomartini, S. Amadio, E. Ambrosini and C. Volonte et al., 2005. ATP regulates oligodendrocyte progenitor migration, proliferation and differentiation: Involvement of metabotropic P2 receptors. Brain Res. Rev., 48: 157-165.
CrossRefDirect Link - Chai, Y.S., J. Hu, F. Lei, Y.G. Wang and Z.Y. Yuan et al., 2013. Effect of berberine on cell cycle arrest and cell survival during cerebral ischemia and reperfusion and correlations with p53/cyclin D1 and PI3K/Akt. Eur. J. Pharmacol., 708: 44-55.
CrossRefDirect Link - Chai, Y.S., Z.Y. Yuan, F. Lei, Y.G. Wang and J. Hu et al., 2014. Inhibition of retinoblastoma mRNA degradation through Poly (A) involved in the neuroprotective effect of berberine against cerebral ischemia. PLoS ONE, Vol. 9.
CrossRefDirect Link - Cui, H.S., K. Matsumoto, Y. Murakami, H. Hori and Q. Zhao et al., 2009. Berberine exerts neuroprotective actions against in vitro ischemia-induced neuronal cell damage in organotypic hippocampal slice cultures: Involvement of B-cell lymphoma 2 phosphorylation suppression. Biol. Pharm. Bull., 32: 79-85.
CrossRefDirect Link - Frantseva, M.V., P.L. Carlen and H. El-Beheiry, 1999. A submersion method to induce hypoxic damage in organotypic hippocampal cultures. J. Neurosci. Methods, 89: 25-31.
CrossRefDirect Link - Giaume, C., F. Kirchhoff, C. Matute, A. Reichenbach and A. Verkhratsky, 2007. Glia: The fulcrum of brain diseases. Cell Death Differ., 14: 1324-1335.
CrossRefDirect Link - Hsu, Y.Y., Y.T. Tseng and Y.C. Lo, 2013. Berberine, a natural antidiabetes drug, attenuates glucose neurotoxicity and promotes Nrf2-related neurite outgrowth. Toxicol. Applied Pharmacol., 272: 787-796.
CrossRefDirect Link - Hu, J., Y. Chai, Y. Wang, M.M. Kheir and H. Li et al., 2012. PI3K p55γ promoter activity enhancement is involved in the anti-apoptotic effect of berberine against cerebral ischemia-reperfusion. Eur. J. Pharmacol., 674: 132-142.
CrossRefDirect Link - Karadottir, R., P. Cavelier, L.H. Bergersen and D. Attwell, 2005. NMDA receptors are expressed in oligodendrocytes and activated in ischaemia. Nature, 438: 1162-1166.
CrossRefDirect Link - Kaur, C., V. Sivakumar, L.S. Ang and A. Sundaresan, 2006. Hypoxic damage to the periventricular white matter in neonatal brain: Role of vascular endothelial growth factor, nitric oxide and excitotoxicity. J. Neurochem., 98: 1200-1216.
CrossRefDirect Link - Kew, J.N.C. and J.A. Kemp, 2005. Ionotropic and metabotropic glutamate receptor structure and pharmacology. Psychopharmacology, 179: 4-29.
CrossRefDirect Link - Matute, C., 2006. Oligodendrocyte NMDA receptors: A novel therapeutic target. Trends Mol. Med., 12: 289-292.
CrossRefDirect Link - Matute, C., 2010. Calcium dyshomeostasis in white matter pathology. Cell Calcium, 47: 150-157.
CrossRef - Matute, C., E. Alberdi, M. Domercq, F. Perez-Cerda and A. Perez-Samartin et al., 2001. The link between excitotoxic oligodendroglial death and demyelinating diseases. Trends Neurosci., 24: 224-230.
CrossRefDirect Link - Matute, C., E. Alberdi, M. Domercq, M.V. Sanchez-Gomez and A. Perez-Samartin et al., 2007. Excitotoxic damage to white matter. J. Anatomy, 210: 693-702.
CrossRefDirect Link - Nadjafi, S., S.A. Ebrahimi and N. Rahbar-Roshandel, 2014. Protective effects of berberine on oxygen-glucose deprivation/reperfusion on oligodendrocyte cell line (OLN-93). Int. J. Prev. Med.
Direct Link - Rahbar-Roshandel, N., L. Razavi, B. Tavakoli-Far and M. Mahmoudian, 2008. Mebudipine and dibudipine protect PC12 cells against oxygen-glucose deprivation and glutamate-induced cell death. Pathophysiology, 15: 227-231.
CrossRefDirect Link - Richter-Landsberg, C. and M. Heinrich, 1996. OLN-93: A new permanent oligodendroglia cell line derived from primary rat brain glial cultures. J. Neurosci. Res., 45: 161-173.
CrossRefDirect Link - Salter, M.G. and R. Fern, 2005. NMDA receptors are expressed in developing oligodendrocyte processes and mediate injury. Nature, 438: 1167-1171.
CrossRefDirect Link - Szydlowska, K. and M. Tymianski, 2010. Calcium, ischemia and excitotoxicity. Cell Calcium, 47: 122-129.
CrossRefDirect Link - Tavakoli-Far, B., N. Rahbar-Roshandel, P. Rahimi-Moghaddam and M. Mahmoudian, 2009. Neuroprotective effects of mebudipine and dibudipine on cerebral oxygen-glucose deprivation/reperfusion injury. Eur. J. Pharmacol., 610: 12-17.
CrossRefDirect Link - Wang, Y. and Z.H. Qin, 2010. Molecular and cellular mechanisms of excitotoxic neuronal death. Apoptosis, 15: 1382-1402.
CrossRefDirect Link - Wang, X., R. Wang, D. Xing, H. Su and C. Ma et al., 2005. Kinetic difference of berberine between hippocampus and plasma in rat after intravenous administration of Coptidis rhizoma extract. Life Sci., 77: 3058-3067.
CrossRefDirect Link - Wilkins, A., H. Majed, R. Layfield, A. Compston and S. Chandran, 2003. Oligodendrocytes promote neuronal survival and axonal length by distinct intracellular mechanisms: A novel role for oligodendrocyte-derived glial cell line-derived neurotrophic factor. J. Neurosci., 23: 4967-4974.
Direct Link - Yao, S., P. Pandey, A. Ljunggren-Rose and S. Sriram, 2010. LPS mediated injury to oligodendrocytes is mediated by the activation of nNOS: Relevance to human demyelinating disease. Nitric Oxide, 22: 197-204.
CrossRefDirect Link - Zhang, Q., Z. Qian, L. Pan, H. Li and H. Zhu, 2012. Hypoxia-inducible factor 1 mediates the anti-apoptosis of berberine in neurons during hypoxia/ischemia. Acta Physiologica Hungarica, 99: 311-323.
CrossRefDirect Link