
Javaid Ahmad Shah
Aquatic Ecology Laboratory, Centre of Research for Development, University of Kashmir 190006, J and K, India
Ashok K. Pandit
Aquatic Ecology Laboratory, Centre of Research for Development, University of Kashmir 190006, J and K, India
Pakistan Journal of Biological Sciences
Year: 2013 | Volume: 16 | Issue: 19 | Page No.: 976-983







ABSTRACT
The present study scrutinizes the seasonal variation in the distribution of crustacean zooplankton in relation to physico-chemical liminology of Wular lake. Water samples were collected from five study sites during September 2010-August 2011. Remarkable spatial and temporal variations were observed among the different study sites. Among the parameters recorded water temperature showed positive correlation with pH (p<0.01) and orthophosphate (p<0.05).Transparency showed a significant negative correlation with total alkalinity (p<0.01), free carbon dioxide (p<0.01) and ammonical nitrogen (p<0.01). Crustacean zooplankton were collected by filtering 100 L lake water through a 30 μm nytal mesh and preserved in 4% formalin. A total of 42 taxa of crustacean were recorded, among them 23 species belonged to Cladocera, 16 to Copepoda and only 3 to Ostracoda. Species diversity and density depicted an abrupt up surge on the advent of warmer period (extending from March). Among the various sites a maximum of 40 species were recorded from a single biotope (site III) against the minimum of only 17 species being recorded from site I. Bray-Curtis cluster analysis showed close similarity (0.928-0.944%) between summer and autumn in terms of species diversity. Pearson correlation coefficient of the various physico-chemical parameters of water with crustacean zooplankton depicted that not only a single but multiple factors govern over the distribution and diversity of crustacean in the lake.
PDF Abstract XML References Citation
Received: December 07, 2012;
Accepted: February 26, 2013;
Published: April 16, 2013
How to cite this article
Javaid Ahmad Shah and Ashok K. Pandit, 2013. Relation Between Physico-chemical Limnology and Crustacean Community in Wular Lake of Kashmir Himalaya. Pakistan Journal of Biological Sciences, 16: 976-983.
DOI: 10.3923/pjbs.2013.976.983
URL: https://scialert.net/abstract/?doi=pjbs.2013.976.983
DOI: 10.3923/pjbs.2013.976.983
URL: https://scialert.net/abstract/?doi=pjbs.2013.976.983
INTRODUCTION
Aquatic environments are dynamic systems with cycles and processes operating at a range of spatial and temporal scales. The active natures of the events collectively with natural series of disorderness, anthropogenic pressures, act to allocate the aquatic biotic communities into spatial and temporal diversity (Furnas, 1995; Keough and Butler, 1995; Jeppesen et al., 2002). Further, temporal unpredictability of environmental conditions are known as a prerequisite factor for the structure of communities (Rutherford et al., 1999; Ruokolainen et al., 2009) besides, resources availability in the surrounding environment have a challenging impact on plankton dynamics and are strongly related (Beisner, 2001). The composition and diversity of zooplankton provide information on the characteristics and quality of the water body (Okayi et al., 2001). Crustaceans have worldwide-distribution, establish across all habitats. They are important for evaluation of the impacts of climatic change and anthropogenic pressures on non-model systems (Buhay, 2011). Crustaceans an important constituent of zooplankton and play fundamental role in aquatic food chains. Besides being an important food item of fishes, the animalcules also find use as potential indicators of the trophic status of a water body since their structure and composition are highly affected by eutrophication (Patalas, 1972). Therefore, this study was designed to assess crustacean community in terms of species composition and diversity between various sites, under the operative influence of varying physico- chemical environment of Wular lake, valued as a wetland (a Ramsar Site) of Kashmir Himalaya on which no published work is available till date.
MATERIALS AND METHODS
Study area: Wular lake, a rural lake in the north-west of Kashmir about 55 km from Srinagar city, is situated at an altitude of 1,580 m (a.m.s.l) and lies between 34°16´-34°20´N and 74°33´-74°44´E geographical co-ordinates. It is the largest freshwater lake of the Indian sub-continent. The area of lake as per topo-sheets of 1911 was 217.8 km which includes 58 km of associated marshes.
 | |
| Fig. 1: | Map of Wular lake showing five sampling sites |
The area was reduced from 157.74 to 86.71 km during 1911 to 2007. Overall there was reduction the lake area by 45% mainly due to conversion for agriculture (28%) and Salix plantation (17%). Further associated marshes were reduced by 70% again due to conversions for agriculture and settlements. GIS pictures recently taken reveal the total area of the lake to be 45 km2 (Fig. 1). The shrinkage in lake area was mainly due to continuous siltation brought about by various tributaries (Erin, Madhumati, Ashtung nalla, Hartal nalla, etc.), besides River Jhelum being the main feeding channel. The lake is mono-basined having an elliptical shape and plays a significant role in hydrographic system of Kashmir valley by acting as a huge reservoir and absorbs high annual flood of River Jhelum.
Methods: Water samples were taken from five different sites in both limnetic and littoral zones. Crustacean zooplankton were collected by filtering 100 litre sub-surface water through a 30 ìm nytal mesh and fixed in 4% formalin to which 4-5 drops of glycerine and 5% sucrose were added. Identification of the crustaceans was done with the help of standard works of Pennak (1978), Adoni (1985); Battish (1992) and Edmondson (1992). The quantitative analysis was done with the help of Sedgewick Rafter plankton counter cell and the results were expressed as individual per litre (ind./L). At least three tubes were taken at different sites and integrated to get one composite sample. In addition, crustacean zooplankton was collected in horizontal transects by towing a net. Water samples were also taken in the open water for biological and chemical analyses and some limnological variables were determined in situ (temperature, pH, depth and transparency). The remaining parameters were determined following the standard methods of APHA (1998).
Statistical analysis: Statistical analyses were performed using SPSS (statistical Version 11.5 for Windows 7, SPSS and Chicago, IL, USA). The Pearson correlation (r) was made for the statistical interpretation of the physico-chemical parameters of water and crustacean species density. The relationship between crustacean species abundance with different seasons was determined using a similarity percentage program PAST (statistical Version 1.93 for Windows 7).
RESULTS AND DISCUSSION
Physico-chemical characteristics of water: Mean with minimum and maximum values in the physico-chemical parameters of water is presented in Table 1. During the study period, water temperature ranged between the maximum of 29.13°C and minimum of 3.6°C with average of 15.02±0.77. Transparency of water fluctuated both spatially and temporally. In general, highest transparency values were recorded in winter registering its highest of 1.7 m against the lowest of 0.57 m in summer. Devi and Sharma (2008) also obtained similar results in Oksoipat lake, Manipur. The pH range during present study did not fluctuate much and remained between 7.17-8.27, reflecting alkaline nature of the lake. Water depth ranged from 1 m in winter to 4.27 m in spring. Dissolved Oxygen (DO) revealed a definite seasonal trend registering higher content in winter (11.1 mg L-1) and lower content in summer (4.63 mg L-1). Highest dissolved oxygen during winter could be attributed to the fact that in lower temperature oxygen carrying capacity of water increases (Wetzel, 1983; Desai et al., 1995). The maximum alkalinity value (197.6 mg L-1) was noted in winter, followed by steep fall in the summer till it touched a minimum value (56. 67 mg L-1). Free carbon dioxide registered a minimum value of 11 mg L-1 at in summer and a maximum of 23.3 mg L-1 in winter (average = 15.7±1.28 mg L-1).
| Table 1: | Physico-chemical parameters of water in Wular lake during Sep. 2010 to Aug. 2011 |
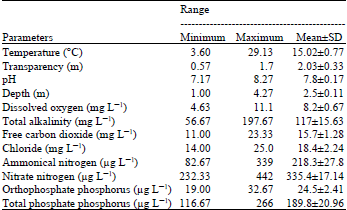 | |
Chloride content within the lake fluctuated in a narrow range 14.0-22.67 mg L-1, being recorded in winter and summer respectively. These results corroborates with findings of Govindan and Sundaresan (1979) and Jana (1973). NH4-N concentrations fluctuated between 82.7 in summer to 339 μg L-1 in winter, the average being 218.3±27.8 mg L-1. The relatively higher levels of ammonical nitrogen in winter may be due to low microbial activity which converts ammonia into nitrites and nitrates. Nitrate nitrogen concentrations fluctuated between 232.3-442 μg L-1, being observed in summer and in winter respectively (average = 335.4±17.14). Phosphorous, is generally recognized as one of the key nutrients in the productivity of freshwaters as it is essential element determining fertility of lakes. According to Cole (1975), the high concentration of phosphate in water is due to decay and subsequent mineralization or decomposition of dead organic matter and surface run-off.
On the basis of Pearson inter-correlation matrix positive correlation was observed between water temperature and pH (r = 0.944, p<0.01) and orthophosphate (r = 0.536; p<0.05) which almost corroborate with the findings of Essien-Ibok et al. (2010). Depth showed negative correlation with transparency (r = 0.680; p<0.01), pH (r = 0.652; p<0.01) and orthophosphate (r = 0.45; p<0.05). Transparency in the present investigation showed a significant negative correlation with total alkalinity(r = 0.918; p<0.01), free carbon dioxide (r = 0.713; p<0.01), ammonical nitrogen (r = 0.890; p<0.01), nitrate-nitrogen (r = 0.780; p<0.01) and total phosphate (r = 0.789, p<0.01). Ammonical nitrogen in the present investigation showed positive significant correlation with nitrate nitrogen (r = 0.711, p<0.01) and total phosphate phosphorus (r = 0.743, p<0.01), whileas, nitrate nitrogen showed positive correlation with total phosphate phosphorus (r = 0.711, p<0.01).
Crustacean community
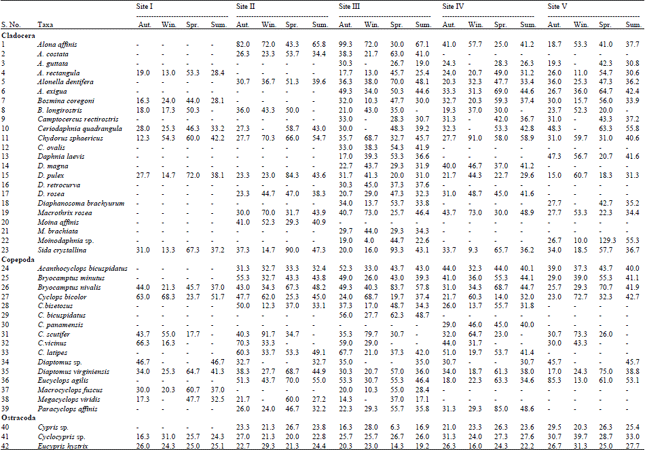
Species composition: The diversity of crustacean groups and its seasonal variation at five study sites are summarized in Table 2. Total number of recorded species were 42 belonging to three different groups namely Cladocera (23), Copepoda (16) and Ostracoda (3). Highest number of crustacean species (40) was registered at site III, followed by 31, 28, 27 and 17 at sites IV, V, II and site I, respectively. Further, group-wise distribution of crustacean zooplankton indicates discernible differences at different studied sites. Cladocera, thus recorded 22, 16, 15, 11 and 07 species at sites III, V, IV, II and I in a decreasing order, followed by Copepoda recording 15 at site III, 13 each at sites IV and II, 9 at site V and 8 at site I respectively. Ostracoda was the least represented group registering only 03 species each at sites II, III, IV and only 2 species at site I. The most dominant cladocerans reported in the lake were Chydorus sphaericus, Alona affinis, Macrothrix rosea and Moinodaphnia sp. Among Copepoda dominant species included Bryocamptus nivalis, Cyclops bicolor and Eucyclops agilis. Among Ostracoda, Cyclocypris sp. and Eucypris hystrix were the main representatives of the group. Other species of crustaceans were found either sporadically or were of relatively low abundance. Rich diversity and abundance of crustaceans in lake is due to shallow nature of the lake associated with macrophytes that play an important role in the structure and dynamics of the microcrustacean assemblages (Scheffer et al., 2006).
Seasonal variation in population density: In general, Cladocera showed unimodal growth peak in spring season (Table 2). The spring maximum is attributed to accumulation of organic matter in the lake due to rise of primary productivity on account of rise in temperature (Sharma and Kotwal, 2011). Cladocerans have high seasonal variability due to the undulation of parthenogenetic and gamogenetic reproduction modes (Egloff et al., 1997; Rivier, 1998). A rapid increase in densities accomplished as a result of high rates of embryonic and postembryonic growth is a distinguishing feature of cladocerans, besides some abiotic factors such as temperature that modulates egg development (Pandit, 1998, 1980; Saunders et al., 1999; Mergeay et al., 2006; Hansson et al., 2007). Cladocerans attain high population abundances very rapidly, but persists over a very short time only. Reasons for a sudden decline in abundance in colder periods (winter and autumn) is not immediately known, but we believe decrease is typically accompanied by an intensive gamogenetic reproduction (Onbe, 1978) and low food availability, that result in the formation of resting eggs at the end of the reproductive season.
| Table 2: | Seasonal variations in the population density of crustacean zooplankton at five study sites of Wular lake* |
 | |
| *: Average results based on three analyses, Aut.: Autumn, Win.: Winter, Spr.: Spring, Sum.: Summer and -: Absent | |
| Table 3: | Correlation of crustacean groups with abiotic parameters |
 | |
| *: Correlation at 0.05(2-tailed) **: Correlation at 0.01(2-tailed), WT: Water temperature, Transp: Transparency, DO: Dissolved oxygen, TA: Total alkalinity, CO2: Free carbon dioxide, Cl: Chloride, NH4-N: Ammonical nitrogen, NO3-N: Nitrate nitrogen, OP: Orthophosphate phosphorus and TP: Total phosphate phosphorus | |
This has been suggested to be triggered by varying environmental conditions and aimed at ensuring survival of the species over the winter (Egloff et al., 1997; Rivier, 1998). Among the cladocerans species like Bosmina longirostris, being thermophobic, was completely absent in the summer season at all the sites while as species like Alona guttata, Camptocercus rectirostris, Ceriodaphnia quadrangula and Diaphanosoma brachyurum were not encountered throughout winter, an observation in consonance with the earlier findings of Hakkari (1978) for Finland lakes and Yousuf and Qadri (1981), Yousuf et al. (1983) and Balkhi and Yousuf (1996) for Kashmir lakes.
Copepoda was the second dominant group, after Cladocera, in terms of both species number and density. During the present study peak populations of Copepoda vary among the study sites autumn being the peak growth season for sites I (345 ind. L-1) and II (568.3 ind. L-1) and spring for sites III, IV and V respectively registering 629, 569 and 364 ind. L-1 (Table 2). In general, copepods were dominant in warmer period. Habitat preferences and environmental conditions including food availability may be the reasons for peak populations (Santer and Lampert, 1995; Alekseev et al., 2007). Studies revealed that species like Megacyclops viridis and Diaptomus sp. were completely absent in winter though the latter one was also absent in spring. Likewise, Cyclops scutifer and C. vicinus were absent in summer and the latter was again missing in spring season. These findings are inconsonance with the earlier studies of Yousuf (1988) and Balkhi and Yousuf (1990, 1992).
Ostracoda was represented by Cypris sp., Cyclocypris sp. and Eucypris hystrix. In general, no regular growth pattern was followed by Ostracoda as majority of them were found to live in the bottom of the lake. The population dynamics of this group is not clearly known though some species exhibit distinct seasonal periodicity (Wetzel, 1983). According to Pennak (1978) ostracods can tolerate wide range of ecological stresses.
Within a particular lake, the species composition of aquatic communities are closely linked with seasonal and hydrologic cycles. Many species of zooplankton, can, however, tolerate changing conditions by both physiological and life cycles adaptations (Gauthier, 1928; Margaritora, 1983; Ramdani, 1988). For example, the resistant eggs of Ostracoda, Cladocera and Copepoda (Diaptomidae) can tolerate dry periods. Some adult species of Copepoda (Cyclopoidae) can transform into a dormant stage (Dussart, 1969). However, when environmental changes exceed species tolerance limits, community changes at species level usually occurs. During the present study crustaceans depicted significant positive correlation with temperature (p<0.05), thereby indicating that temperature has a strong influence on the crustaceans abundance, a fact well supported by Bhuiyan and Gupta (2007); Park and Shin (2007) and Rajagopal et al. (2010). Temperature is said to have the major influences on the distribution of crustaceans and has been reported to affect zooplankton (crustaceans) abundance in two ways. First it acts directly to hasten growth rates, thereby increasing population densities and secondly, it stimulates the growth of phytoplankton population which provides nutrients.
It is known that there is a positive correlation between temperature and species richness of zooplankton in aquatic environments (Matsubara, 1993; Castro et al., 2005; Hessen et al., 2007). Moreover, other water quality parameters such as nitrate and phosphate also affected, but negatively in deciding the abundance of copepods (Table 3). Thus, it is noteworthy to note that when the concentration of nitrate (r = 0.804, p<0.01) and phosphate were more, the abundance of copepods was less (Joseph and Yamakanamardi, 2011). Regarding the depth, somewhat direct relationship exists between the two (p<0.01). Further significant negative correlation was observed with free CO2 (Table 3; p<0.01 for Cladocera; p<0.05 for Copepoda) as proposed by Welch (1952) and Moshood (2009). Crustaceans preferring less alkaline waters (Qadri and Yousuf, 1980) infers that there exists inverse relation between the crustaceans (particularly Copepoda) and the total alkalinity (r = 0-.642, p<0.01) (Paulose and Meheswari, 2008). In the present study a direct relation was observed between the pH and the crustaceans which corroborates with the findings of Basu et al. (2010).
Bray-curtis cluster analysis showed great similarity between summer and autumn (0.928-0.944%).
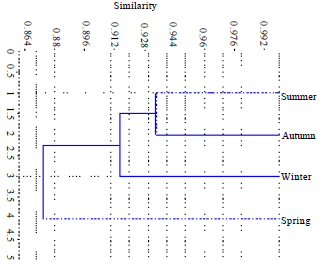 | |
| Fig. 2: | Bray-curtis cluster analysis of crustaceans at five study sites |
Contrary to these two seasons, spring season showed maximum dissimilarity during the entire study period, representing the most favourable period for crustacean density and diversity (Fig. 2).
CONCLUSION
From the present study, it can be inferred that the diversity and distribution of crustaceans in the Wular lake is controlled by a combination of abiotic (temperature, depth, pH and alkalinity) as well as biotic factors including food availability, predation, alternation of parthenogenetic and gamogenetic reproduction modes. Low abundance and diversity was observed at site I due to anthropogenic stresses and low macrophytic growth. Temporal variability in a multitude of environmental conditions is considered to be the main regulatory factor for crustacean zooplankton distribution, abundance and diversity, therefore, rebuffing the single factor concept controlling the entire plankton community.
ACKNOWLEDGMENT
Thanks are due to the Director, Centre of Research for Development and Head, Environmental Science, University of Kashmir for providing necessary laboratory facilities.
REFERENCES
- APHA., 1998. Standard Methods for Examination of Water and Wastewater. 20th Edn., American Public Health Association, Washington, DC., ISBN-13: 978-0875532356, Pages: 1220.
Direct Link - Beisner, B.E., 2001. Plankton community structure in fluctuating environments and the role of productivity. Oikos, 95: 496-510.
CrossRefDirect Link - Buhay, J.E., 2011. Population dynamics of crustaceans: Introduction to the symposium. Integr. Comp. Biol., 51: 577-579.
CrossRef - Castro, B.B., S.C. Antunes, R. Pereira, A.M.V.M. Soares and F. Goncalves, 2005. Rotifer community structure in three shallow lakes: Seasonal fluctuations and explanatory factors. Hydrobiologia, 543: 221-232.
CrossRefDirect Link - Essien-Ibok, M.A., A.W. Akpan, M.T. Udo, L.A. Chude, I.A. Umoh and I.E. Asuquo, 2010. Seasonality in the physical and chemical characteristics of Mbo river, Akwa Ibom State, Nigeria. Niger. J. Agric. Food Environ., 6: 60-72.
Direct Link - Furnas, M.J., 1995. Land-Sea Interactions and Oceanographic Processes Affecting the Nutrient Dynamics and Productivity of Australian Marine Ecosystems. In: The State of the Marine Environment Report for Australia, Zann, L.P. and P. Kaiola (Eds.). Common Wealth Government Printer, Canberra, Australia, ISBN-13: 9780642174062, pp: 61-74.
- Hansson, L., S. Gustafsson, K. Rengefors and L. Bomark, 2007. Cyanobacterial chemical warfare affects zooplankton community composition. Freshwater Biol., 52: 1290-1301.
CrossRef - Hessen, D.O., V. Bakkestuen and B. Walseng, 2007. Energy input and zooplankton species richness. Ecography, 30: 749-758.
CrossRef - Jana, B.B., 1973. Seasonal periodicity of plankton in a freshwater pond in West Bengal, India. Int. Rev. Hydrobiol., 58: 127-143.
CrossRefDirect Link - Jeppesen, E., J.P. Jensen and M. Sondergaard, 2002. Response of phytoplankton, zooplankton and fish to reoligotrophication: An 11-year study of 23 Danish lakes. Aquat. Ecosyst. Health Manage., 5: 31-43.
Direct Link - Joseph, B. and S.M. Yamakanamardi, 2011. Monthly changes in the abundance and biomass of zooplankton and water quality parameters in Kukkarahalli lake of Mysore, India. J. Environ. Biol., 32: 551-557.
PubMed - Matsubara, T., 1993. Rotifer community structure in the South Basin of Lake Biwa. Hydrobiologia, 271: 1-10.
CrossRefDirect Link - Mergeay, J., S. Declerck, D. Verschuren and L. De Meester, 2006. Daphnia community analysis in shallow Kenyan lakes and ponds using dormant eggs in surface sediments. Freshwater Biol., 51: 399-411.
CrossRef - Moshood, K.M., 2009. Zooplankton assemblage of Oyun Reservoir, Offa, Nigeria. Revista de Biologia Tropical, 4: 1027-1047.
Direct Link - Park, K.S. and H.W. Shin, 2007. Studies on phyto-and-zooplankton composition and its relation to fish productivity in a West coast fish pond ecosystem. J. Environ. Biol., 28: 415-422.
PubMedDirect Link - Patalas, K., 1972. Crustacean plankton and the eutrophication of St. Lawrence Great Lakes. J. Fish. Res. Board Can., 29: 1451-1462.
CrossRefDirect Link - Paulose, P.V. and K. Meheswari, 2008. Seasonal variation in zooplankton community structure of Ramgarah lake, Jaipur, Rajasthan. Proceeding of the 12th World Lake Conference, October 29-November 2, 2007, Jaipur, India, pp: 82-87.
Direct Link - Rajagopal, T., A. Thangamani, S.P. Sevarkodiyone, M. Sekar and G. Archunan, 2010. Zooplankton diversity and physico-chemical conditions in three perennial ponds of Virudhunagar district, Tamilnadu. J. Environ. Biol., 31: 265-272.
PubMedDirect Link - Ramdani, M., 1988. Stagnant water ecosystems in Morocco: Biotypological and biogeographical studies of the zooplankton. Trav. Inst. Sci. Rabat Zool., 43: 1-40.
Direct Link - Ruokolainen, L., E. Ranta, V. Kaitala and M.S. Fowler, 2009. Community stability under different correlation structures of species environmental responses. J. Theor. Biol., 261: 379-387.
CrossRef - Rutherford, S., S. D'Hondt and W. Prell, 1999. Environmental controls on the geographic distribution of zooplankton diversity. Nature, 400: 749-753.
CrossRef - Santer, B. and W. Lampert, 1995. Summer diapause in cyclopoid copepods: Adaptive response to a food bottleneck? J. Anim. Ecol., 64: 600-613.
Direct Link - Saunders, P.A., K.G. Porter and B.E. Taylor, 1999. Population dynamics of Daphnia spp. and implications for trophic interactions in small, monomictic lake. J. Plankton Res., 21: 1823-1845.
CrossRef - Scheffer, M., G.J. Van Geest, K. Zimmer, E. Jeppesen and M. Sondergaard et al., 2006. Small habitat size and isolation can promote species richness: Second-order effects on biodiversity in shallow lakes and ponds. Oikos, 112: 227-231.
CrossRef