
B. Jemmali
Department of Animal Production, School of Higher Education in Agriculture of Mateur, Tunisia
A. Ben Gara
Department of Animal Production, School of Higher Education in Agriculture of Mateur, Tunisia
H. Selmi
Regional Field crops Research Center Beja, Tunisia
Z. Ammari
Department of Animal Production, School of Higher Education in Agriculture of Mateur, Tunisia
C. Bouheni
Department of Animal Production, School of Higher Education in Agriculture of Mateur, Tunisia
M. Ben Larbi
National Agricultural Institute of Tunisia, Tunisia
M. Hammami
Department of Animal Production, School of Higher Education in Agriculture of Mateur, Tunisia
M. Amraoui
Department of Animal Production, School of Higher Education in Agriculture of Mateur, Tunisia
M. Kamoun
Department of Animal Production, School of Higher Education in Agriculture of Mateur, Tunisia
H. Rouissi
Department of Animal Production, School of Higher Education in Agriculture of Mateur, Tunisia
B. Rekik
Department of Animal Production, School of Higher Education in Agriculture of Mateur, Tunisia
Pakistan Journal of Biological Sciences
Year: 2013 | Volume: 16 | Issue: 24 | Page No.: 2031-2035







ABSTRACT
The genetic polymorphism of the goat Kappa casein was investigated in Tunisian goats. Blood samples were collected from local goat breeds. Samples of genomic DNA were obtained from leukocytes of 175 dairy goats and regions of interest in the gene were amplified by Polymerase Chain Reaction (PCR) and then evaluated in agarose gel. For a better characterization of the single nucleotide polymorphism, a PCR-Restriction Fragment Length Polymorphism was performed employing the endonuclease DNA amplification using 459 bp primers. The PCR products of primers (459 bp) digested by restriction enzyme Alw44I produced two fragments of 459 and 381 bp. The Kappa casein allelic variants in tested animals revealed different genotypes, two of them were homozygous: AA or BB, AC or BC and CC. Genotypic frequencies were 12.5, 60.5 and 27% for AA or BB, CC and AC or BC, respectively. Identification of different variants of the Kappa casein can be used for the improvement and conservation of Tunisian local goats.
PDF Abstract XML References Citation
Received: March 16, 2013;
Accepted: April 03, 2013;
Published: September 16, 2013
How to cite this article
B. Jemmali, A. Ben Gara, H. Selmi, Z. Ammari, C. Bouheni, M. Ben Larbi, M. Hammami, M. Amraoui, M. Kamoun, H. Rouissi and B. Rekik, 2013. Kappa Casein Gene Polymorphism in Local Tunisian Goats. Pakistan Journal of Biological Sciences, 16: 2031-2035.
DOI: 10.3923/pjbs.2013.2031.2035
URL: https://scialert.net/abstract/?doi=pjbs.2013.2031.2035
DOI: 10.3923/pjbs.2013.2031.2035
URL: https://scialert.net/abstract/?doi=pjbs.2013.2031.2035
INTRODUCTION
Goat population in Tunisia is around 1.3 million heads distributed over different regions all over the country. The distribution of these goats is uneven as shown in Fig. 1. Goat genetic resources are concentrated in the arid regions, mainly in the South. Farmers appreciate the rusticity, mothering ability and resistance to health disorders of the local goat and consider its kid meat as the main product (Ammar et al., 2011) in addition to milk for household consumption. Milk is an essential product not only in providing young needs but also in nutrition of humans because of its richness in versatile components from minerals to vitamins (Jann et al., 2004; Haenlein, 2004; Botaro et al., 2009; Rekik, 2012).
The composition of the milk of different species varies with respect to contents of these constituents. All milks contain relatively the same kinds of constituents, but in varying amounts. Within a given species, genetic factors and environmental conditions (climatic conditions, management and the stage of lactation) influence milk yield and composition.
Goat milk composition is comparable to other dairy animals. The most important component of this milk is proteins.
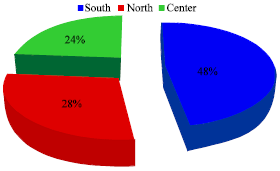 | |
| Fig. 1: | Geographic distribution of goats in Tunisia (2012) |
The four caseins expressed in goat milk, αS1-, β-, αS2- and κ-casein, are coded by the loci CSN1S1, CSN2, CSN1S2 and CSN3, respectively, located within a 250 kb segment of goat chromosome 6 (Martin et al., 2002; Feligini et al., 2005; D'Alessandro et al., 2007; Jemmali et al., 2012).
Nowadays, studies of the genetic polymorphism in goats and other domesticated farm species address the impact of genetic differences on production traits and health and, for this purpose, techniques capable of detecting small variations in the DNA molecule are used (Byun et al., 2008; Crisa et al., 2010; Jemmali et al., 2011b; Hirwa et al., 2011; Kawaguchi, 2013). Milk protein polymorphism, that is genetic variation, affect protein yield and quality and consequently milk products properties such as cheese yield and quality (Aleandri et al., 1990; Lodes et al., 1996; Falaki et al., 1997). Various studies suggest important relationships between casein polymorphisms and milk production traits (Bovethuts and Weller, 1994; Ikonen et al., 1999; Velmala et al., 1999). The Kappa casein plays an important role in the formation, stabilization and aggregation of the casein micelles thus altering the manufacturing properties and digestibility of milk. In the present study, 175 animals were analyzed to detect the genetic variants of Kappa casein in Tunisian local goats.
MATERIALS AND METHODS
Sample collection and DNA extraction: A total of 175 local goats were randomly sampled. Blood samples were collected in vacutainer tubes containing EDTA (1 mg mL-1). Genomic DNA was extracted using standard protocol (Easy-DNA™ Kit Invitrgen) and stored at -20°C until used in assay.
The concentration of DNA samples was estimated using UV-visible range spectrophotometer and diluted to 50 ng μL-1 before PCR amplification. All the DNA samples had 260/280 OD ratios in the range of 1.8 to 2, indicating high purity. DNA was also examined by loading samples on 0.8% agarose gel and visualizing the band under UV light with a Gel Doc 1000 system (BioRad) after ethidium bromide staining.
PCR amplification: The PCR was performed in a final volume of 25 μL containing 100 ng of template DNA, 50 pmole of each primer, 5 μL of 10X PCR buffer (20 mM Tris-HCL pH 8.4, 50 mM KCl), 1.5 mM MgCl2, 0.2 mM of dNTPs and 1 U of Taq DNA polymerase (Jemmali et al., 2011a). Kapa-casein gene polymorphisms were identified by PCR-RFLP (Baltrenaite et al., 2009). Primer sequences used were:
| • | Forward:5’-TGT-GCT-GAG-TAG-GTA-TCC-TAGTTA-TGG-3’ |
| • | Reverse:3’-GAT-TCC-TCT-GTA-GTT-TCT-CCT-GTT-GCG-5’ |
The digestion of 459 bp PCR product was performed using endonucleases Alw44I (SibEnzyme, Italy). This solution was initially denatured at 94°C for 5 min followed by 39 cycles of denaturation (95°C for 30 sec), annealing (60°C for 45 sec) and elongation (72°C for 1 min) and a final extension at 72°C for 10 min. The PCR products were electrophoresed on 2% agarose gels in order to check the quality and specificity of DNA fragment amplification.
PCR-RFLP condition: For PCR-RFLP analysis, the 459 bp PCR products were digested with Alw44I (SibEnzyme, Italy). Restriction fragments (459 and 381 bp) were separated by electrophoresis in a 2% agarose gel and their sizes were estimated using the molecular markers (Invitrogen, 1 kb DNA Ladder). A volume of 20 μL of PCR products was digested for 4 h at 37°C with 10 units of restriction enzyme. Digested products were separated by electrophoresis on a 2% agarose gel and visualized with Ethidium bromide under UV light with a Gel Doc 1000 system (BioRad) after ethidium bromide staining.
RESULTS
Early and precise identification of milk protein polymorphism have great impact on breeding strategies (Gouda et al., 2013). Using the PCR-RFLP technique for the identification of the genetic polymorphism in goat casein allowed both rapid and efficient determination of the genetic variation in this gene regardless of the age and sex of animals (Jemmali et al., 2012).
PCR reaction and DNA amplification: The primers described by Baltrenaite et al. (2009) amplified DNA fragments with sizes of 459 bp, corresponding to cited primers. After the reaction was completed, the samples were quantified to visualize the amplified fragments. Only one strand was detected (Fig. 2).
PCR-RFLP (PCR-restriction fragment length polymorphism) analysis: Digestion of fragment amplified region by restriction endonuclease Alw44I generated two fragments of 459 and 381 bp (Fig. 3). The results of the RFLP analysis for used endonucleases demonstrated the existence of two alleles. The digested fragment of 381 bp represents the CC genotype. Undigested fragments of 459 bp, represent the AA or BB genotype. The digested fragment of 381 bp and the undigested fragment of 459 bp indicate the CA or CB genotype. Present results revealed that the Kappa casein allelic variants in the tested breeds showed different genotypes, two of them were homozygous. Frequencies were 12.5 and 60.5% for AA or BB and CC, respectively. The remaining genotypes were heterozygous either AC or BC (27%).
DISCUSSION
Kappa-casein gene variation: Kappa casein gene exon 4 was analyzed for sequence variations. A total of nine SNPs were identified. Seven SNPs had been identified previously in goat populations. Yahyaoui et al. (2001) identified polymorphism in Spanish and French breeds in the positions 245 (nucleotide change from T to C), 309 (G to A), 471 (G to A) and in the position 591 (T to C).
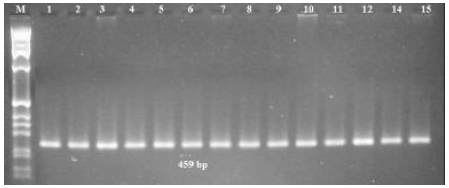 | |
| Fig. 2: | Results after PCR amplification of the Kappa casein gene, Line M-DNA Ladder, 1 Kb, Line 1-15: Analyzed animals of local goats |
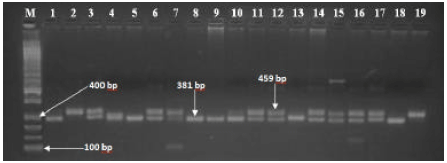 | |
| Fig. 3: | DNA electrophoretic pattern obtained after digestion of PCR amplified goat kappa-casein products with Alw44I, Lane M: 1 kb DNA ladder marker, Lines 1-4-5.8-9-10-13-18: Digested fragment of 381 bp (specific for the C allele and the CC genotype), Lines 2-7-19: Undigested fragment of 459 bp (specific for A or B allele) and Line 3-6-11-12-14-15-16-17: Digested fragment of 381 bp with indigested fragment of 459 bp (specific for C and A or B alleles) |
Polymorphism in the position 247 (A to G) was first identified by Caroli et al. (2001) in Italian goat breeds, while polymorphism in the positions 384 (G to A) and 550 (T to C) was previously identified by Prinzenberg et al. (1999) and Prinzenberg et al. (2005). These SNPs have also been described in various goat populations outside Eastern Africa (Angiolillo et al., 2002; Yahyaoui et al., 2003; Jann et al., 2004).
Blood collection and DNA extraction: A (5 mL) blood sample was collected from each animal and 300 mL aliquots were then used for DNA extraction according to the previously described methodology. After extraction, the DNA was then submitted to electrophoresis in (0.8%) agarose gel, in order to achieve concentration and quality in each all DNA samples. The concentration of DNA samples was estimated using UV visible range spectrophotometer and diluted to 50 ng μL-1 before PCR amplification. All the DNA samples had 260/280 OD ratios in the range of 1.8 to 2, indicating high purity.
PCR reaction and DNA amplification: PCR product (459 bp) specified the amplified band of Kappa casein region on exon 4. Our result is similar to those founded by Coll et al. (1993). The efficacy of PCR is measured by its specificity, efficiency and fidelity. A highly specific PCR will generate one and only one amplification product that is the intended target sequence showed in Fig. 2. More efficient amplification will generate more products with fewer cycles. A highly accurate PCR, will contain a negligible amount of DNA polymerase-induced errors in its product. An ideal PCR would be the one with high specificity, yield and fidelity. Studies indicate that each of these parameters is influenced by numerous components of PCR, including the buffer conditions, the PCR cycling regime and DNA polymerases. Unfortunately, adjusting conditions for maximum specificity may not be compatible with high yield; likewise, optimizing for the fidelity of PCR may result in reduced efficiency. The use of PCR-RFLP method can be a pathway to detected single nucleotide polymorphism.
PCR-restriction fragment length polymorphism analysis: The digested fragments by Alw44I produced two restriction patterns at 381 and 78 bp for allele C, while the PCR product form A or B remained intact (Fig. 3). A second digestion with HaiIII is necessary to detect single nucleotide polymorphism if exist. This is the first report of an analysis of the Kappa casein gene by the PCR-RFLP technique in Tunisian local goats. The variant Kappa-casein genotype found in these animals was CC and its allele was C. The variant genotype CC codes for high cheese yield.
Milk yield is a quantitative trait. Animals used in the study had milk production varying from high to low. The Kappa-casein gene could be a source of variation of yield of these animals, but, this influence could not be measured. In goats, the Kappa-casein gene can be used as a molecular marker to identify milk mixtures in cheese making processes, because in cattle and other species this same gene is polymorphic. The allele C is frequent in the majority of the goat populations.
CONCLUSION
The present study revealed that DNA amplification using primers produced fragments with sizes of 459 bp. The PCR products of primer (459 bp) digested by restriction enzyme Alw44I produced two fragments at 459 and 381 bp. Three genotypes were identified. Different variants of the Kappa casein can be a pathway to conserve local goat breeds.
ACKNOWLEDGMENT
This study was carried in Animal Biotechnology laboratory of INAT, we acknowledge the help of Pr. Haddad Brahim; Pr. Khaldi Gley and Pr. Romdhane Mohamed Salah.
REFERENCES
- Aleandri, R., L.G. Buttazzoni, J.C. Schneider, A. Caroli and R. Davoli, 1990. The effects of milk protein polymorphisms on milk components and cheese-producing ability. J. Dairy Sci., 73: 241-255.
CrossRefDirect Link - Ammar, H., R. Boda, M. Ben Younes and S. Lopez, 2011. Goat Breeding Systems in the South of Tunisia (Tataouine). In: Economic, Social and Environmental Sustainability in Sheep and Goat Production Systems, Bernues, A., J.P. Boutonnet, I. Casasus, M. Chentouf and D. Gabina et al. (Eds.). CIHEAM/FAO/CITA-DGA, Zaragoza, Spain, pp: 283-288.
Direct Link - Angiolillo, A., M.H. Yahyaoui, A. Sanchez, F. Pilla and J.M. Folch, 2002. Characterization of a new genetic variant in the caprine k-casein gene. J. Dairy Sci., 85: 2679-2680.
Direct Link - Baltrenaite, L., S. Kerziene, K. Morkuniene and I. Miceikiene, 2009. Genetic diversity in milk proteins among goats breed in Lithuania. Acta Univ. Latviensis, 753: 117-124.
Direct Link - Botaro, B.G., Y.V.R. de Lima, C.S. Cortinhas, L.F.P. e Silva, F.P. Renno and M.V. dos Santos, 2009. Effect of the kappa-casein gene polymorphism, breed and seasonality on physicochemical characteristics, composition and stability of bovine milk. R. Bras. Zootec., 38: 2447-2454.
CrossRefDirect Link - Bovenhuis, H. and J.I. Weller, 1994. Mapping and analysis of dairy cattle quantitative trait loci by maximum likelihood methodology using milk protein genes as genetic markers. Genetics, 136: 267-280.
PubMedDirect Link - Byun, S.O., H. Zhou, R.H.J. Forrest, C.M. Frampton and J.G.H. Hickford, 2008. Association of the ovine calpastatin gene with birth weight and growth rate to weaning. Anim. Genet., 39: 572-573.
CrossRef - Caroli, A., O. Jann, E. Budelli, P. Bolla, S. Jager and G. Erhardt, 2001. Genetic polymorphism of goat k-casein (CSN3) in different breeds and characterization at DNA level. Anim. Genet., 32: 226-230.
Direct Link - Crisa, A., C. Marchitelli, L. Pariset, G. Contarini and F. Signorelli et al., 2010. Exploring polymorphisms and effects of candidate genes on milk fat in dairy sheep. J. Dairy Sci., 93: 3834-3845.
PubMed - D'Alessandro, A.G., G. Martemucci and D. Casamassima, 2007. Behavioural profile during suckling period in Martina Franca breed jennies reared under semi-extensive conditions. Applied Anim. Behav. Sci., 106: 107-124.
CrossRef - Falaki, M., A. Prandi, C. Corradini, M. Sneyers and N. Gengler et al., 1997. Relationships of growth hormone gene and milk protein polymorphism to milk production traits in Simmental cattle. J. Dairy Res., 64: 47-56.
Direct Link - Feligini, M., S. Frati, V.C. Curik, A. Brambilla and P. Parma et al., 2005. Caprine αs1-casein polymorphism: Characterisation of A, B, E and F variants by means of various biochemical and molecular techniques. Food Technol. Biotechnol., 43: 123-132.
Direct Link - Gouda, E.M., M.K. Galal and A.S. Abdelaziz, 2013. Genetic variants and allele frequencies of kappa casein in Egyptian cattle and buffalo using PCR-RFLP. J. Agric. Sci., 5: 197-203.
Direct Link - Haenlein, G.F.W., 2004. Goat milk in human nutrition. Small Rumin. Res., 51: 155-163.
CrossRefDirect Link - Hirwa, C.A., P. Wallace, X. Shen, Q. Nie, G. Yang and X. Zhang, 2011. Genes related to economically important traits in beef cattle. Asian J. Animal Sci. 5: 34-45.
CrossRefDirect Link - Ikonen, T., M. Ojala and O. Ruottinen, 1999. Associations between milk protein polymorphism and first lactation milk production traits in Finnish Ayrshire cows. J. Dairy Sci., 82: 1026-1033.
CrossRefDirect Link - Jemmali, B., S. Bedhiaf and M. Djemali, 2011. Frequence de genes de prolificite en race ovine Barbarine en Tunisie [Frequency of genes in prolific sheep breed Barbarine Tunisia]. Rencontres Recherches Ruminants, 18: 422-422.
Direct Link - Jemmali, B., S. Bedhiaf and M. Djemali, 2011. PCR-RFLP of BMPR-IB gene in North African barbarine sheep. Res. Opin. Anim. Vet. Sci., 1: 200-203.
Direct Link - Jemmali, B., M. Kamoun, M. Haddar, A. Ben Gara and H. Selmi et al., 2012. Genetic polymorphism of casein α-S1 gene in Tunisian local goat. BIOMIRROR, 3: 1-4.
Direct Link - Kawaguchi, A., 2013. PCR-RFLP identifies differences in hrpZ sequences to distinguish two genetic groups of Pseudomonas syringae pv. syringae strains from barley and wheat with bacterial black nod. J. Gen. Plant Pathol., 79: 51-55.
CrossRef - Lodes, A., I. Krause, J. Buchberger, J. Aumann and H. Klostermeyer, 1996. The influence of genetic variants of milk proteins on the compositional and technological properties of milk. 1. Casein micelle size and the content of non-glycosylated κ-casein. Milchwissenschaft, 51: 368-373.
Direct Link - Martin, P., M. Szymanowsk, L. Zwierzchowsk and C. Leroux, 2002. The impact of genetic polymorphisms on the protein composition of ruminant milks. Reprod. Nutr. Dev., 42: 433-459.
CrossRefDirect Link - Prinzenberg, E.M., K. Gutscher, S. Chessa, A. Caroli and G. Erhardt, 2005. Caprine κ-Casein (CSN3) polymorphism: New developments in molecular knowledge. J. Dairy Sci., 88: 1490-1498.
CrossRefDirect Link - Velmala, R.J., H.J. Vilkki, K.T. EIo, D.J. de Koning and A.V. Maki-Tanila, 1999. A search for quantitative trait loci for milk production traits on chromosome 6 in Finnish Ayrshir cattle. Anim. Genet., 30: 136-143.
PubMed - Yahyaoui, M.H., A. Angiolillo, F. Pilla, A. Sanchez and J.M. Folch, 2003. Characterization and genotyping of the caprine kappa casein variants. J. Dairy Sci., 86: 2715-2720.
Direct Link - Yahyaoui, M.H., A. Coll, A. Sanchez and J.M. Folch, 2001. Genetic polymorphism of the caprine kappa casein gene. J. Dairy Res., 68: 209-216.
CrossRefDirect Link