
R.K. Kar
Plant Physiology and Biochemistry Laboratory, Department of Botany, Visva-Bharati University, Santiniketan-731 235, West Bengal, India
N. Parvin
Department of Botany, Biology-1, Biocentre, Ludwig Maximillians, University of Munich, 82152 Martinsried, Germany
D. Laha
Centre for Plant Molecular Biology, Eberhard Karls, University of Tuebingen, Auf der Morgenstelle 1, 72076 Tuebingen, Germany
Pakistan Journal of Biological Sciences
Year: 2013 | Volume: 16 | Issue: 24 | Page No.: 1991-1996







ABSTRACT
Regulation of stomatal aperture is crucial in terrestrial plants for controlling water loss and gaseous exchange with environment. While much is known of signaling for stomatal opening induced by blue light and the role of hormones, little is known about the regulation of stomatal closing in darkness. The present study was aimed to verify their role in stomatal regulation in darkness. Epidermal peelings from the leaves of Commelina benghalensis were incubated in a defined medium in darkness for 1 h followed by a 1 h incubation in different test solutions [H2O2, propyl gallate, ethrel (ethylene), AgNO3, sodium orthovanadate, tetraethyl ammonium chloride, CaCl2, LaCl3, separately and in combination] before stomatal apertures were measured under the microscope. In the dark stomata remained closed under treatments with ethylene and propyl gallate but opened widely in the presence of H2O2 and AgNO3. The opening effect was largely unaffected by supplementing the treatment with Na-vanadate (PM H+ ATPase inhibitor) and tetraethyl ammonium chloride (K+-channel inhibitor) except that opening was significantly inhibited by the latter in presence of H2O2. On the other hand, H2O2 could not override the closing effect of ethylene at any concentrations while a marginal opening of stomata was found when Ag NO3 treatment was given together with propyl gallate. CaCl2 treatment opened stomata in the darkness while LaCl3 maintained stomata closed. A combination of LaCl3 and propyl gallate strongly promoted stomatal opening. A probable action of ethylene in closing stomata of Commelina benghalensis in dark has been proposed.
PDF Abstract XML References Citation
Received: March 15, 2013;
Accepted: March 27, 2013;
Published: September 16, 2013
How to cite this article
R.K. Kar, N. Parvin and D. Laha, 2013. Differential Role of Ethylene and Hydrogen Peroxide in Dark-induced Stomatal Closure. Pakistan Journal of Biological Sciences, 16: 1991-1996.
DOI: 10.3923/pjbs.2013.1991.1996
URL: https://scialert.net/abstract/?doi=pjbs.2013.1991.1996
DOI: 10.3923/pjbs.2013.1991.1996
URL: https://scialert.net/abstract/?doi=pjbs.2013.1991.1996
INTRODUCTION
Stomata are microscopic pores on the surface of plants which are surrounded by a pair of guard cells, thus becoming under the control of movement of such cells. These are of crucial importance for the control of water loss and gas exchange simultaneously and as such they are major regulators of global carbon and water cycles. Stomatal opening depends on the increased turgidity of guard cells which is brought about by accumulation of solutes such as K+, Cl¯, malate2-and sucrose (Talbott and Zeiger, 1996). Such accumulation is triggered by light via different overlapping ways. These include an accumulation of K+ in response to hyperpolarization caused by a light activated plasma membrane located H+-ATPase (Zeiger, 1983). Stomatal closing, on the other hand, occurs when such solutes diffuse out due to possible membrane depolarization (Schroeder et al., 2001) thereby leading to turgor loss. The hormone abscisic acid (ABA) has been implicated for stomatal closure by activating specific signalling cascades involving H2O2 and Ca2+ as intermediary components (Schroeder et al., 2001; Fan et al., 2004; Zhang et al., 2004). Ethylene has also been demonstrated to induce stomatal closure acting through H2O2 (Desikan et al., 2006) though its role is conflicting. Typically, these experiments on stomatal regulation by ABA or ethylene treatment were carried out in light to demonstrate their counteracting ability. No reports are there so far demonstrating the effect of such agents on stomata in darkness.
In our earlier study on epidermal peeling of Commelina benghalensis leaves we demonstrated the effect of ethylene and H2O2 on stomatal behaviour during light-induced opening (Laha et al., 2009). Interestingly ethylene and Ca2+ was found to be associated with opening while H2O2 induced closure. In the present investigation the role of ethylene, Ca2+ and H2O2 and their interaction during stomatal closure in darkness was verified using again the epidermal strips from leaves of C. benghalensis.
MATERIALS AND METHODS
Twigs of C. benghalensis were collected from the Departmental garden and washed with water before use. Mature leaves from the apical part of the plants were used for peeling epidermal strips from the lower surface of leaves.
Epidermal peels of about 5x10 mm were floated on a media (50 mM KCl and 10 mM MES buffer adjusted to pH 6.15 with KOH) kept in Petri dishes in darkness at a constant temperature (25±1°C). After 1 h strips were transferred to the same media now added with test compounds in specified concentrations while sets were still in dark. Stomatal peelings were treated with H2O2 (1 mM, unless otherwise specified), propyl gallate (0.1 mM, unless otherwise specified), ethrel (commercial preparation of ethylene, 0.1 mM, unless otherwise specified), AgNO3 (1 mM, unless otherwise specified), sodium orthovanadate (1 mM), tetraethyl ammonium chloride (1 mM), CaCl2 (0.1 mM), LaCl3 (1 mM) or combinations of such treatments. After 1 h dark incubation strips were examined under microscope. Stomatal apertures were measured using an ocular micrometer standardized against stage micrometer and the area of the aperture was calculated from length and breadth. Data presented were average of at least ten measurements taken from different microscopic fields.
RESULTS
Figure 1 shows the photo plate of microscopic views of stomata on epidermal peelings of C. benghalensis leaves incubated in darkness in the specified medium added with H2O2 (1 mM) or AgNO3 (1 mM) along with a control set (distilled water). While the stomatal aperture was totally closed under control condition (Fig. 1a), it was widely opened in case of both the treatments (Fig. 1b-c), more prominently with AgNO3. Effect of propyl gallate (0.1 mM), H2O2 (1 mM), AgNO3 (1 mM) and ethylene (ethrel, 0.1 mM) on stomatal aperture of Commelina species kept in darkness has been depicted as bar graph in Fig. 2. Again treatment with H2O2 and AgNO3 opened stomata while stomata remained almost closed in propyl gallate and fully closed in ethylene.
Table 1 includes data on the effect of combined treatment of H2O2 (1 mM) and AgNO3 (1 mM) with either Na-vanadate (1 mM) (inhibitor of plasmalemma H+ ATPase) or tetraethyl ammonium chloride (1 mM) (a potassium channel inhibitor) along with individual treatments for comparison. Individual treatments with H2O2 and AgNO3 opened stomata but apertures remained at control values in Na-vanadate or tetraethyl ammonium chloride. However, stomatal opening caused by AgNO3 was slightly inhibited by both Na-vanadate and tetraethyl ammonium chloride. In contrast, stomatal opening induced by H2O2 could not be decreased by Na-vanadate but significantly inhibited by tetraethyl ammonium chloride treatment.
Table 2 describes the effects of combination of ethylene (0.05, 0.1 mM) with H2O2 (0.5, 1 mM) of different concentrations and combinations of AgNO3 (0.1, 1 mM) with propyl gallate (0.05, 0.1 mM). Stomata remained closed in any combination of ethylene with H2O2. On the other hand, a marginal opening of stomata was found in case of any combination of AgNO3 and propyl gallate.
| Table 1: | Effect of different treatments and their combinations on stomatal opening (area of aperture, μm2) of Commelina benghalensis incubated in darkness |
 | |
| Treatments were: H2O2 (1 mM), AgNO3 (Ag+, 1 mM), Na-Vanadate (1 mM) and tetraethyl ammonium chloride (TEAC, 1 mM) and combinations of these as shown below. Data presented were average of ten replications (Mean±SE) | |
| Table 2: | Effect of different combination of treatments on stomatal opening (area of aperture, μm2) of Commelina benghalensis incubated in darkness |
 | |
| Treatments were either combination of ethylene (0.05 mM or 0.1 mM) and H2O2 (0.5 mM or 1 mM) or combination of AgNO3 (Ag+, 0.1 mM or 1 mM) and propyl gallate (PG, 0.05 mM or 0.1 mM). Data presented were average of ten replications (Mean±SE) | |
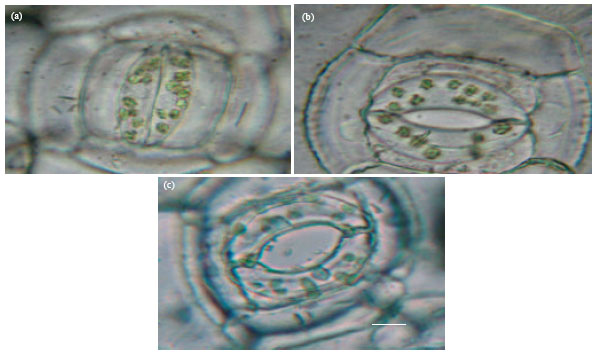 | |
| Fig. 1(a-c): | Photographic plates showing microscopic view of representative stomata of Commelina benghalensis incubated in darkness under treatment with either (b) 1 mM H2O2 or (c) 1 mM AgNO3 (a) Along with control distilled water. Scale bar shown in the figure measures 10 μm |
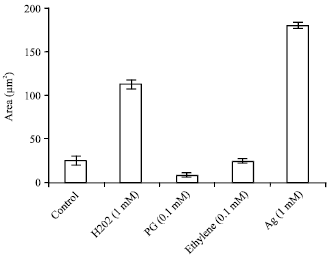 | |
| Fig. 2: | Effect of treatments with 1mM H2O2, 0.1 mM Propyl Gallate (PG), 0.1 mM ethylene and 1 mM AgNO3 along with control (distilled water) on stomatal opening (area of aperture, μm2) of Commelina benghalensis incubated in darkness. SE shown as vertical bars |
Figure 3 depicts the changes in the stomatal aperture during treatment with CaCl2 (0.1 mM), LaCl3 (1 mM) (a plasma membrane located calcium channel blocker) and combinations of LaCl3 with ethylene (0.1 mM) and with propyl gallate (0.1 mM) in darkness. CaCl2 treatment alone caused significant opening of stomata while stomata remained unaffected by LaCl3 treatment.
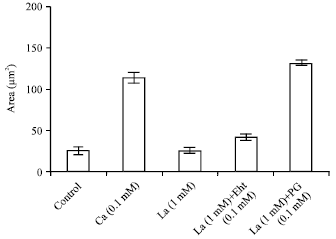 | |
| Fig. 3: | Effect of treatments with CaCl2 (Ca), 0.1 mM, LaCl3 (La), 1 mM and combinations of LaCl3 (La), 1 mM with Propyl Gallate (PG), 0.1 mM and with Ethylene (Eth), 0.1 mM on stomatal opening (area of aperture, μm2) of Commelina benghalensis incubated in darkness. SE shown as vertical bars |
Combining LaCl3 had only a small promoting effect on opening but when combined with propyl gallate, LaCl3 opened stomata widely.
DISCUSSION
Stomatal opening induced by light is known to depend on osmotic accumulation of K+ ions and other solutes like chloride, malate anions and sugars. There is a well-defined promoting role of blue light that acts via induction of plasma membrane H+ ATPase in facilitating K+ uptake (Roelfsema and Hedrich, 2005). However, the regulation of such turgor development in guard cells is controversial, particularly in respect of the exact role of ethylene and H2O2 and Ca2+ as signalling component. in our earlier work on stomatal regulation in leaf epidermal peelings of C. benghalensis in light (Laha et al., 2009) we showed that though H2O2 treatment closed stomata, ethylene and Ca2+ played a positive role. In the present investigation with a similar test system we attempted to clarify the roles of ethylene and H2O2 in stomatal regulation in darkness. H2O2 was found to open stomata in darkness and treatment with propyl gallate, a potent reactive oxygen species scavenger, resulted in closed stomata (Fig. 1 and 2). This is in contrast to the finding of Desikan et al. (2004) who demonstrated inhibition of dark-induced stomatal closure by H2O2 scavenging enzyme catalase or diphenylene iodonium, an inhibitor of reactive oxygen species generating enzyme NADPH oxidase. Also, ABA has been implicated in dark-induced stomatal closure, because closure is impaired in an ABA-deficient mutant (Lee et al., 2006). The action of ABA occurs via signalling components such as Ca2+ and H2O2 (Pei et al., 2000; Kwak et al., 2003). H2O2 is reported to be generated through NADPH oxidase (Kwak et al., 2003) or copper amine oxidase (An et al., 2008). A recent finding showed that elevation of intracellular H2O2 induced by ABA, not constitutive increase in H2O2 level, functions in ABA-induced stomatal closure (Jannat et al., 2011). Exogenous treatment with H2O2 also caused stomatal closure in Arabidopsis thaliana and treatment with fusicoccin suppresses such closure by removing H2O2 (She et al., 2010).
In contrast to the role of ABA, involvement of ethylene in stomatal regulation is much less clearly defined and there are contradictory reports. On the one hand it is found responsible for stomatal closing (Desikan et al., 2006), on the other hand ethylene inhibits ABA-induced stomatal closure (Tanaka et al., 2005, 2006). In the former case ethylene-induced stomatal closure was seemingly mediated by H2O2 produced by NADPH oxidase (Desikan et al., 2006). In one recent report, ethylene was found to inhibit darkness-induced stomatal closure by scavenging Nitric Oxide (NO) (Song et al., 2011). Surprisingly, in epidermal peelings of C. benghalensis leaves ethylene treatment in the dark was without effect but AgNO3 (1 mM), an inhibitor of ethylene perception, increased stomatal aperture markedly (Fig. 1, 2), apparently indicating a possible role of ethylene in dark-induced closing of stomata. This is in contrast to its probable role in light where treatment opened stomata in the same species (Laha et al., 2009).
Opening of stomata, either in darkness or light must involve osmoregulation (Talbot and Zeiger, 1998). Likely candidates involved in such regulation are plasma membrane H+ ATPase and K+ channels along with other transporters (Pandey et al., 2007). Thus possible involvement of plasma membrane H+ ATPase and K+ channels in case of stomata opening induced by Ag+ and H2O2 in darkness in the present system was tested by combination treatments of these agents with the inhibitors, Na-orthovanadate (an inhibitor of plasma membrane H+ ATPase) and tetraethyl ammonium chloride (an inhibitor of K+ channels) (Table 1). It appeared that opening of stomata in darkness resulting from inhibition of ethylene action by AgNO3 was not fully dependent on the action of plasma membrane H+ ATPase and K+ channels, as the inhibitors could marginally inhibit stomatal opening. However, H2O2-induced stomatal opening was only sensitive to tetraethyl ammonium chloride indicating a role of K+ channels in such opening in darkness. In an early work on dark opening of stomata under CO2 free air, area of stomatal aperture was correlated with the K+ content of the guard cells (Rogers et al., 1980). It may be assumed that in case of H2O2-induced stomatal opening K+ uptake occurred via K+-channels, although it is not clear how H2O2 can drive such K+ uptake.
Ethylene and H2O2 have counteracting roles in stomatal regulation instead of a collinear action at least in case of C. benghalensis. This was also found in case of light incubation of the epidermal peels of the same species by Laha et al. (2009). However, in case of dark experiments ethylene could override the opening effect of H2O2 (Table 2) apparently through different pathway. On the other hand, in absence of ethylene action (AgNO3 treatment) reactive oxygen species scavenger (propyl gallate treatment) could not close stomata completely. This may be either due to residual amount of H2O2 that acted towards opening or resulted from complex interaction of signalling pathways.
Calcium is well known for its role in signalling for a number of plant processes and cytosolic Ca2+ plays a pivotal role in stomatal regulation (Allen et al., 2001). Level of cytosolic Ca2+ may be controlled either by the entry of apoplastic Ca2+ through plasma membrane located Ca2+ channels or release from endosomal stores. Increased cytosolic Ca2+ level, as downstream effect, may have impact on anion channels and plasma membrane H+ ATPase. As such Ca2+ has been reported to be involved in stomatal closing as an intermediate in the cascade induced by ABA or H2O2. On the contrary, our earlier finding in C. benghalensis leaves was that Ca2+ is involved in stomatal opening in light as La3+ (inhibits the entry of apoplastic Ca2+ in the cytosol) inhibited opening and also Ca2+ treatment prevented H2O2-or Ag+-induced stomatal closing (Laha et al., 2009). In the present investigation with the same species, Ca2+ treatment was again found to trigger stomatal opening even in darkness. Although the sensitivity of stomata to extracellular Ca2+ may vary (Roelfsema and Hedrich, 2005), no report is there so far about the opening effect of Ca2+ except one report on C. benghalensis leaves showing Ca2+ requirement for blue light-induced stomatal opening (Parvathi and Raghavendra, 1997) and our earlier report (Laha et al., 2009). LaCl3-treatment showed no effect when given individually or marginal opening when supplemented with ethylene. Ethylene-induced stomatal opening in light in C. benghalensis was explained by possible release of endosomal Ca2+ release (Laha et al., 2009), but in darkness ethylene is associated with stomatal closing through some different mechanism. Interestingly, a combined treatment of LaCl3 with propyl gallate resulted in wide opening of stomata, which was also observed in light in the same species (Laha et al., 2009), but in the latter case individual treatment with propyl gallate caused stomatal opening. It is suggested that a complex interplay of Ca2+ influx from apoplast and Ca2+ release from endosomal stores through coordinated regulation of respective channels or transporters maintaining Ca2+ homeostasis (Roelfsema and Hedrich, 2005) and accordingly stomatal aperture is regulated by a defined window of guard cell cytosolic Ca2+ oscillation parameters (Allen et al., 2001).
Apparently ethylene operates in darkness to maintain the stomatal closure and probably it acts through maintaining a critical cytosolic Ca2+ balance by controlling Ca2+ influx from apoplast and Ca2+ release from endosomal stores. H2O2 induces stomatal opening in darkness probably by controlling K+ channel activity which may lie downstream of Ca2+ signalling. Further in-depth research and analysis may provide support to this contention, which is apparently in contrast to other findings.
REFERENCES
- Allen, G.J., P.C. Sarah, C.L. Harrington, K. Schumacher and T. Hoffmann et al., 2001. A defined range of guard cell calcium oscillation parameters encodes stomatal movements. Nature, 411: 1053-1057.
CrossRefDirect Link - An, Z., W. Jing, Y. Liu and W. Zhang, 2008. Hydrogen peroxide generated by copper amine oxidase is involved in abscisic acid-induced stomatal closure in Vicia faba. J. Exp. Bot., 59: 815-825.
Direct Link - Desikan, R., M.K. Cheung, A. Clarke, S. Golding and M. Sagi et al., 2004. Hydrogen peroxide is a common signal for darkness- and ABA-induced stomatal closure in Pisum sativum. Funct. Plant Biol., 31: 913-920.
Direct Link - Desikan, R., K. Last, R. Harrett-Williams, C. Tagliavia and K. Harter et al., 2006. Ethylene-induced stomatal closure in Arabidopsis occurs via AtrbohF-mediated hydrogen peroxide synthesis. Plant J., 47: 907-916.
CrossRefPubMedDirect Link - Fan, L.M., Z.X. Zhao and S.M. Assmann, 2004. Guard cells: A dynamic signaling model. Curr. Opin. Plant Biol., 7: 537-546.
CrossRefPubMedDirect Link - Kwak, J., I.C. Mori, Z.M. Pei, N. Leonhardt and M.A. Torres et al., 2003. NADPH oxidase AtrbohD and AtrbohF genes function in ROS-dependent ABA signaling in Arabidopsis. EMBO J., 22: 2623-2633.
CrossRef - Laha, D., N. Parvin and R.K. Kar, 2009. Role of ethylene and its interaction with hydrogen peroxide and calcium in light-induced stomatal opening of Commelina benghalensis. J. Theor. Exp. Biol., Vol. 5, No. 3.
Direct Link - Lee, K.H., H.L. Piao, H.Y. Kim, S.M. Choi and F. Jian et al., 2006. Activation of glucosidase via stress-induced polymerization rapidly increased active pools of abscisic acid. Cell, 126: 1109-1120.
CrossRefDirect Link - Pandey, S., W. Zhang and S.M. Assmann, 2007. Roles of ion channels and transporters in guard cell signal transduction. FEBS Lett., 581: 2325-2336.
CrossRef - Parvathi, K. and A.S. Raghavendra, 1997. Blue light-promoted stomatal opening in abaxial epidermis of Commelina benghalensis is maximal at low calcium. Physiol. Plant., 101: 861-864.
CrossRef - Pei, Z.M., Y. Murata, G. Benning, S. Thomine and B. Klusener et al., 2000. Calcium channels activated by hydrogen peroxide mediate abscisic acid signalling in guard cells. Nature, 406: 731-734.
CrossRefDirect Link - Roelfsema, M.R.G. and R. Hedrich, 2005. In the light of stomatal opening: New insights into the Watergate. New Phytol., 167: 665-691.
CrossRef - Rogers, C., P.J.H. Sharpe and R.D. Powell, 1980. Dark opening of stomates of Vicia faba in CO2-free air: Effect of temperature on stomatal aperture and potassium accumulation. Plant Physiol., 65: 1036-1038.
CrossRef - Schroeder, J.I., G.J. Allen, V. Hugouvieux, J.M. Kwak and D. Waner, 2001. Guard cell signal transduction. Ann. Rev. Plant Physiol. Plant Mol. Biol., 52: 627-658.
Direct Link - She, X.P., A.X. Huang, J. Li and X.Z. Han, 2010. Inhibition of dark-induced stomatal closure by fusicoccin involves a removal of hydrogen peroxide in guard cells of Vicia faba. Physiol. Plant., 140: 258-268.
CrossRef - Song, X.G., X.P. She, J. Wang and Y.C. Sun, 2011. Ethylene inhibits darkness-induced stomatal closure by scavenging nitric oxide in guard cells of Vicia faba. Funct. Plant Biol., 38: 767-777.
CrossRef - Talbott, L.D. and E. Zeiger, 1996. Central roles for potassium and sucrose in guard-cell osmoregulation. Plant Physiol., 111: 1051-1057.
Direct Link - Talbott, L.D. and E. Zeiger, 1998. The role of sucrose in guard cell osmoregulation. J. Exp. Bot., 49: 329-337.
CrossRefDirect Link - Tanaka, Y., T. Sano, M. Tamaoki, N. Nakajima, N. Kondo and S. Hasezawa, 2005. Ethylene inhibits abscisic acid-induced stomatal closure in Arabidopsis. Plant Physiol., 138: 2337-2343.
Direct Link - Tanaka, Y., T. Sano, M. Tamaoki, N. Nakajima, N. Kondo and S. Hasezawa, 2006. Cytokinin and auxin inhibit abscisic acid-induced stomatal closure by enhancing ethylene production in Arabidopsis. J. Exp. Bot., 57: 2259-2266.
CrossRef - Zeiger, E., 1983. The biology of stomatal guard cells. Annu. Rev. Plant Physiol., 34: 441-475.
CrossRef - Zhang, X., H. Wang, A. Takemiya, C.P. Song, T. Kinoshita and K. Shimazaki, 2004. Inhibition of blue light-dependent H+ pumping by abscisic acid through hydrogen peroxide-induced dephosphorylation of the plasma membrane H+-ATPase in guard cell protoplasts. Plant Physiol., 136: 4150-4158.
CrossRefDirect Link