
M. Ikhwanuddin
Institute of Tropical Aquaculture, Universiti Malaysia Terengganu, Kuala Terengganu, Malaysia
M.N. Azra
Institute of Tropical Aquaculture, Universiti Malaysia Terengganu, Kuala Terengganu, Malaysia
H. Siti-Aimuni
Faculty of Fisheries and Aqua-Industries, Universiti Malaysia Terengganu, Kuala Terengganu, Malaysia
A.B. Abol-Munafi
Faculty of Fisheries and Aqua-Industries, Universiti Malaysia Terengganu, Kuala Terengganu, Malaysia
Pakistan Journal of Biological Sciences
Year: 2012 | Volume: 15 | Issue: 15 | Page No.: 720-728







ABSTRACT
Blue swimming crab, Portunus pelagicus is widely study and research throughout the Indo-West Pacific, but little is known of its reproductive biology in Malaysia. The present study describes the fecundity, embryonic development and ovarian development stages of the P. pelagicus from Johor coastal water, Malaysia. Carapace width range of berried crabs sampled was from 9.64 to 13.32 cm, while the body weight range was from 75 to 235 g. The mean number of egg produced by females in different sizes ranged from 105443.333±35448.075 per eggs batch. Mean egg size during embryonic development at stage 1 was 0.307±0.037, while 0.386±0.039 and 0.396±0.033 for stage 2 and stage 3, respectively. Study showed that there was significant (p<0.05) relationship between the number of eggs and carapace width/body weight. Mean diameter oocyte during ovarian development at stage 1 was 97.732±12.391 while for stage 2 was 149.516±23.287. Stage 3 showed increasingly of size with mean diameter was 158.506±27.616 and 181.013±24.339 for stage 4.
PDF Abstract XML References Citation
Received: October 04, 2012;
Accepted: December 10, 2012;
Published: January 24, 2013
How to cite this article
M. Ikhwanuddin, M.N. Azra, H. Siti-Aimuni and A.B. Abol-Munafi, 2012. Fecundity, Embryonic and Ovarian Development of Blue Swimming Crab, Portunus pelagicus (Linnaeus, 1758) in Coastal Water of Johor, Malaysia. Pakistan Journal of Biological Sciences, 15: 720-728.
DOI: 10.3923/pjbs.2012.720.728
URL: https://scialert.net/abstract/?doi=pjbs.2012.720.728
DOI: 10.3923/pjbs.2012.720.728
URL: https://scialert.net/abstract/?doi=pjbs.2012.720.728
INTRODUCTION
Blue swimming crab, Portunus pelagicus is a commercially important species (Ikhwanuddin et al., 2011; Ikhwanuddin et al., 2012a, b) and its fishery occupies a significant place due to its high nutritive value, delicacy and good market price. The increasing demands have led to a growing interest in the aquaculture of this crab. Many countries in Asia like China (Lai et al., 2010), India (Soundarapandian et al., 2007; Soundarapandian and Singh, 2008; Bhat et al., 2011; Japan (Hamasaki et al., 2011), Thailand (Nitiratsuwan et al., 2010), Indonesia (Rejeki, 2007) and Malaysia (Ikhwanuddin et al., 2012c) are actively involved in P. pelagicus fisheries and research.
World total landings for P. pelagicus were approximately 158,688 tons in 2006 and increased to 172,144 tons in 2007 (FAO, 2007). In Malaysia, the total landings of P. pelagicus were 3,514 tons in 2007 and increased to 4,427 tons in 2008. State of Johor has been the major catching areas for P. pelagicus in Malaysia, contributing to about 4,195 tons (22%) of total production 18,349 tons in period 2005-2008 (Department of Fisheries, 2009). Though there is high market demand on the fisheries of P. pelagicus, information is very scarce in the case of reproductive biology.
Fecundity, embryonic and ovarian development are important aspects of reproductive biology of crab. Much work has been conducted on various aspects of the reproductive biology of many Portunus spp. such as in Australia (Sumpton, 2001; Kumar et al., 2003; De Lestang et al., 2003a, b; Xiao and Kumar, 2004; Stewart et al., 2010; Johnson et al., 2010), China (Wu et al., 2010), India (Samuel and Soundarapandian, 2009; Josileen, 2011; Sahoo et al., 2011; Ravi et al., 2011), Japan (Hamasaki et al., 2003, 2004, 2006), Pakistan (Rasheed and Mustaquim, 2010), Taiwan (Lee and Hsu, 2003) but few data available in tropical region especially from Malaysia except by Arshad et al. (2006) and Ikhwanuddin et al. (2009, 2011).
Thus, the information pertaining to the reproductive biology of P. pelagicus offer a unique and valuable tool for furthering our understanding of developmental processes and determine appropriate broodstock options for larval rearing of this species. The objective of this study are to determine the fecundity, embryonic and ovarian development of P. pelagicus from Johor coastal water, Malaysia.
MATERIALS AND METHODS
Crab samples: Samples were taken from the Gelang Patah (1°22’60N, 103°37’60E), coastal water of Johor, Malaysia. A total of 42 female crabs randomly sampled of various sizes from Aug. to Oct. 2011 where 30 berried females in various size were sampled to determine the crab fecundity and embryonic development stages while 12 non-berried females were sampled to determine the ovarian development stages through histological assessment.
Carapace width (CW) and body weight (BW) measurements: The CW of the crab was measured to the nearest 0.1 cm using vernier calipers. Crab size was measured as the external CW which is the distance between the 9th anterolateral spines of the carapace (Ikhwanuddin et al., 2009). The BW of the crab was measured to the nearest gram by using a digital electronic balance of 0.1 g sensitivity.
Fecundity and embryonic examination: The berried females were killed and the fresh eggs were stripped off from the broad abdomen of the crab pleopod. The egg mass was washed with fresh water before been preserved in Modified Gilson’s fluid (Ikhwanuddin et al., 2011). The chemical amount used to prepare this preservative were as follows; 100 mL of 60% alcohol, 880 mL of water, 15 mL of 80% nitric acid, 18 mL of glacial acetic acid and 20 g of mercuric chloride. After that, the preserved egg mass was shaken vigorously and left for 24 h. After 24 h, the shaken activities repeated to help in separation of the eggs from the eggs tissues and assisted the penetration of the preservative. The egg mass was preserved for three months (Ikhwanuddin et al., 2011). After three months, the preserved egg mass was washed with distilled water before measuring eggs diameter and counting using volumetric method.
Eggs colouration and eggs size: Matured eggs externally were captured using Cannon EOS DSLR 1100D. The eggs were described by different egg colourations. The eggs were measured using Nikon Measuring Microscope; total amount of the eggs measured was 100 eggs. The eggs also were counted by using the Sedge-Wick Rafter cell after transferred into the 10 mL measuring cylinder in which the eggs are left to settle down. Five replicate subsamples from each ovary were taken. Then the number of eggs in each subsample was counted using a profile projector (Nikon). Lastly, the egg was counted and the fecundity was taken.
Ovarian examination: Crabs were dissected in order to determine the ovarian development stages. The external morphology and the gonad were determined. The external morphological characteristics of the ovary were determined before fixation. Small portions of the anterior, middle and posterior ovarian lobes were fixed in Bouin’s solution about less than 12 h. Then, the gonad was removed into the 70% alcohol solution before proceed with tissue processing. Tissue processing was done in the Automatic Tissue Processor Machine. After processing and hydration of tissues, wax impregnation was done through embedding processed in a paraffin wax to form a solid block.
After that, solid block was cut into 5 μm sections and put in the water bath for a floating process in order to stretch up the sample. The samples were stained with haematoxylin and eosin (Mohajeri et al., 2011) and mounted on slides with DPX. The diameter of at least 90 oocytes for each ovarian lobe was measured with an image analyser. A small portion of ovary was fixed in Davidson’s fixation for further histological study. Tissue from the middle of the left ovarian lobe of female P. pelagicus was dissected for histological examination. The tissue were dehydrated in ascending ethanol concentrations from 70-100%, transferred to lemosol and embedded in paraffin. The tissue were sectioned to 5 μm and stained with Mayer’s hematoxylin-eosin. The development stage of the oocyte was determined histologically by making reference to those of other crabs (Islam et al., 2010).
Data analysis: All results were presented as Means±Standard Deviation (SD). The difference was displayed as statistically significant when p<0.05. The means of BW, CW and oocyte diameter were compared using Analysis of Variance (ANOVA), followed by Duncan’s multiple range test.
RESULTS
Crab samples: Carapace width range of berried crabs sampled was from 9.64 to 13.32 cm, while the BW range was from 75 to 235 g.
Fecundity: The mean number of egg produced by females in different sizes ranged from 105443.333±35448.075 per eggs batch. Minimum fecundity was found in a berried female crab having CW of 9.74 cm and BW of 75 g while the maximum fecundity was found in a berried female crab having CW of 13.32 cm and BW of 235 g (Table 1).
| Table 1: | Analysis of P. pelagicus female crab fecundity and its related characteristics |
 | |
| n = 30 | |
Eggs colouration and eggs size during embryonic development: Table 2 showed egg coloration and mean egg size diameter during embryonic development of P. pelagicus. Eggs of P. pelagicus were spherical and bright orange in color when first deposited. It became yellow, brown and black before reaching the hatching stage. Eggs diameter varies from stage to stage of the embryonic development. Mean egg size during embryonic development at stage 1 was 0.307±0.037), while 0.386±0.039 and 0.396±0.033 for stage 2 (Fig. 1) and stage 3 (Fig. 2), respectively.
 | |
| Fig. 1: | Stage 1 of embryonic development with yellow-orange colour eggs |
 | |
| Fig. 2: | Stage 2 of embryonic development with brown colour eggs |
 | |
| Fig. 3: | Stage 3 of embryonic development with black colour eggs |
Figure 3 represents stage 3 of embryonic development stage with black egg colour.
Relationship between carapace width and fecundity: Carapace width range was measured for determining CW-fecundity relationship. Analysis regression showed that there was significant (p<0.05) relationship between the number of berried eggs and CW. For stage 1 of embryonic development, linear regression was F = 8715 CW-5617 (R2 = 0.065) (Fig. 4). Linear regression for stage 2 was F = 33956 CW-28892 (R2 = 0.502) (Fig. 5) while stage 3 was F = 32916 CW-25269 (R2 = 0.574) (Fig. 6). All the relationship between CW and fecundity of stage 1, stage 2 and stage 3 of embryonic development was found to be linear as shown in Fig. 4-6.
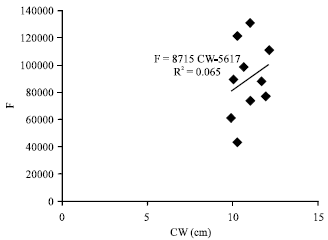 | |
| Fig. 4: | Relationship between carapace width (CW) and fecundity (F) of female P. pelagicus at stage 1 of embryonic development |
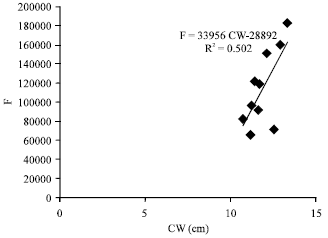 | |
| Fig. 5: | Relationship between carapace width (CW) and fecundity (F) of female P. pelagicus at stage 2 of embryonic development |
| Table 2: | Egg coloration and mean egg size diameter during embryonic development of P. pelagicus |
 | |
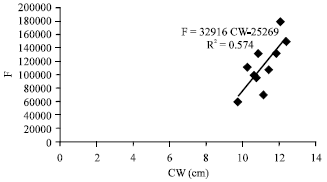 | |
| Fig. 6: | Relationship between carapace width (CW) and fecundity (F) of female P. pelagicus for stage 3 of embryonic development |
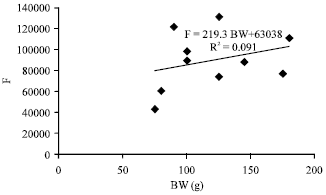 | |
| Fig. 7: | Relationship between body weight (BW) and fecundity (F) of female P. pelagicus for stage 1 of embryonic development |
Relationship between body weight and fecundity: For stage 1 of embryonic development, linear regression was F = 219.3 BW+63038 (R2 = 0.091) (Fig. 7). Linear regression for stage 2 was F = 735.7 BW+2184 (R2 = 0.709) (Fig. 8) while stage 3 was F = 814.0 BW+11758 (R2 = 0.621) (Fig. 9). The result showed that fecundity increasing as BW increased.
Ovarian development stage: A total of 12 female crabs were dissected to observe the ovarian development through histological assessment. CW range from 9.64 to 13.05 cm and BW range from 83 to 200 g of crab samples were used to determine the ovarian development. The progress of ovarian development was classified into four stages based on the presence of the most advanced oocytes with the histological observation (Table 3, Fig. 10).
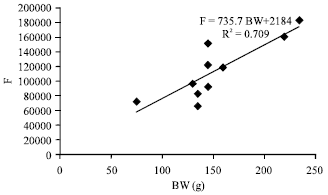 | |
| Fig. 8: | Relationship between body weight (BW) and fecundity (F) of female P. pelagicus for stage 2 of embryonic development |
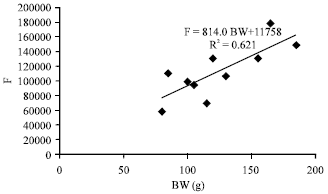 | |
| Fig. 9: | Relationship between body (BW) weight and fecundity (F) of female P. pelagicus for stage 3 of embryonic development |
For stage 1 also known as immature stage, the color of the oocytes was sometimes difficult to recognize because the oocytes showed transparent and translucent in color. Furthermore, stage 1 ovary was built from oogonia that concentrated mostly at the periphery of the ovary. Each oogonium that formed in ovary was spherical in shape and the cytoplasm is hardly visible.
Stage 2 (early maturing), ovary was yellow in color and small egg globules started to appear inside the larger oocytes as soon as vacuolated globules disappeared from the cytoplasm. But, the egg globules did not occupy the entire cytoplasm. Stage 3 or late-maturing stage showed that ovary was light orange in color. In this stage, the oocyte grows rapidly and the nucleus reached maximum size.
| Table 3: | Morphological and histological characteristics of the ovarian development of P. pelagicus |
 | |
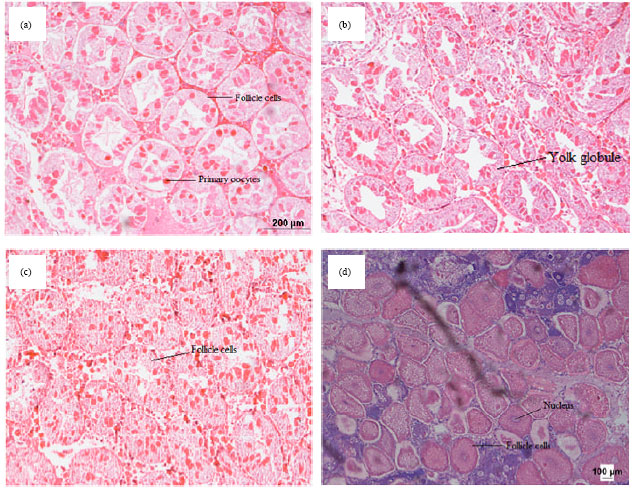 | |
| Fig. 10(a-d): | Stages (a) Immature (stage 1), (b) Early maturing (stage 2), (c) Late maturing (stage 3) and (d) Fully mature (stage 4) of ovarian development of P. pelagicus based on the histological assessment using 10x magnification |
Yolk globules formed in the cytoplasm with larger globular inclusions, while follicle cells were hardly recognizable. In the mature stage (stage 4), the color of the ovary become deep orange or reddish and started to enlarge to the maximum and eventually covered the hepatopancreas and the majority of stomach.
Size of the oocytes varies from stage to stage of the ovarian development. Mean diameter oocyte for stage 1 was 97.732±12.391 while for stage 2 was 149.516±23.287. Stage 3 showed increasingly of size with mean diameter was 158.506±27.616 and 181.013±24.339 for stage 4.
DISCUSSION
The color and size of the crab eggs may well indicate its ‘age’ or maturity. Generally there were three obvious colors of the crab eggs which is yellowish-orange, brown and black. The color of the eggs will change as the embryos develop. At the early stage of newly laid eggs, crab eggs present yellowish to orange in color. The changes continues to brown and from brown to black as the egg yolk was used up. According to Batoy et al. (1987), the eggs swell as they develop and nearly double their newly laid volume by the time they are ready to hatch. Through previous study, result of mean diameter of the egg support this observation which is the older eggs are significantly larger than the newly laid eggs. From this study, mean diameter of the eggs at stage 1 was much smaller which is 0.307 mm compare to stage 2 and stage 3 which was 0.386 and 0.396 mm, respectively. P. pelagicus eggs were translucent and spherical in shape. The shape of P. pelagicus eggs were the same as other Portunid crab such as Callinectes sapidus (Johnson, 1980) and Scylla serrata (Robertson and Kruger, 1994). The results of the present study also shows that there was a significant relationship between the number of the eggs produced and CW. The number of eggs per female increased with increasing CW.
Fecundity refers to reproductive output which indicates the number of eggs produced in every batch. In previous studies by Kumar (2000), P. pelagicus produces more than one batch of egg in one season. Egg production is directly related to BW and growth of individual. The fecundity of P. pelagicus varied from 43, 100 to 183, 100 at the CW range from 9.74 to 13.32 mm and 75-235 g BW. While the mean number of eggs was 105443. 333±35448. 075 for a female with an mean CW of 11.288±0.901 mm and 132±41 g. Previous studies by Kumar (2000) showed that the fecundity of P. pelagicus varied between 651, 505 (> 95-105 mm CW and 0.169 kg BW) to 761, 432 (>125 mm CW and 0.421 kg BW). This difference may be attributing to the loss of eggs during incubation period or during handling as the crab were obtained from the commercial landing.
The number of eggs produced per ovulation in the previous study was large and varies with the size of the crab and between females. Previous study showed that larger female with larger CW produce more eggs than smaller ones with smaller CW (Hamasaki et al., 2006). Other researchers such as De Lestang et al. (2003a) and Arshad et al. (2006) also described about this trend of studies. Furthermore, Kumar et al. (2003) has revealed that fecundity will increased 83.9% with an increase of CW from 10.5 to 12.5 cm. The present study also showed that fecundity of female P. pelagicus initially increased with CW, reaches the maximum point and decreased thereafter. Through this study, it has been found that the number of eggs increased linearly with the increase of CW, BW and weight of eggs oviposition. These positive significant correlations between fecundity and CW and BW were the same as the result by Kumar (2000) on P. pelagicus. The variation in fecundity is very common in Portunid crab and has been reported by other researchers such as De Lestang et al. (2003a) and Johnson et al. (2010).
Rasheed and Mustaquim (2010) described that the size of the berried females varied from 63 to 130 mm CW. Minimum fecundity (272,000) was found in a crab having 63 mm CW whereas the maximum fecundity total 1,395,000 was found in crab having CW of 120 mm. The mean fecundity was 663,000±283,000 for berried female crabs with a mean of CW of 90±15 mm. While the study from west coast of Australia, these data, together with the batch fecundities of different size crabs, indicate that the estimated number of eggs produced by P. pelagicus during the spawning season ranges from about 78,000 in small crabs which was had a 80 mm of CW to about 1,000,000 in large crabs had CW 180 mm. This study showed that the number of first zoeas of Portunidae increased with female body size and the relationship between these two variables was described using a power function.
In Australia, the total fecundity of the year in this species is dependent on batch fecundity, number of ovipositions during an intermolt period, growth rate at molt and number of molts per year. According to De Lestang et al. (2003a), the number of batches estimated in the field during the breeding season correlated with female body size in P. pelagicus. Nakaoka (2003) explained that the females invest the same amount of energy in the reproductive output throughout the breeding season, so that they increase egg number with decreasing of egg size dependent on the trade-off between egg size and the number. Study on cancrid crab, Cancer setosus by Brante et al. (2003) along the Chilean coast showed that the egg number and size was no significant difference with the latitudinal (temperature) gradient, but egg size and reproductive output decreased in Northern Chile (high temperature).
The ovarian development classified based on external characteristics correlated closely with the oocyte diameter and ovarian histology. The general histological features regarding information of oogenesis and ovarian development stages of S. paramamosain are similar to those of S. serrata, S. olivacea and other brachyuran crabs such as P. pelagicus (De Lestang et al., 2003a). Cheng et al. (2002) stated that the major phases of oogenesis in crabs were the previtellogenic phase which contains primary oocytes and the vitellogenic phase where size oocytes increased as yolk was developed into the cytoplasm. Through, Robertson and Kruger (1994) studies, six stages were identified in ovarian development of S. serrata based on external and histological features of the ovary excluding the spent stage.
The developing oocytes and the follicle cells are the major cell types within the ovarian lobes of Scylla sp. Johnson (1980) described the follicle cells are larger and more obvious in immature gonads which is similar to the blue crab. The follicle cells surround each oocytes will flattened when the maturation occurs. The follicle cells also will form the chorionic membrane of the mature ovum (Ryan, 1967). Vacuolated globules were noticed in the cytoplasm of oocytes before initiation of vitellogenesis which were shown to be oil globules in other crustaceans. Oil globules are functioning to supply protein in prawns and crabs. A general feature of crustacean maturation is the production of yolk-laden eggs. The yolk serves as the source of nutrition for the developing embryo because of the yolks contains proteins, lipids, sugars and some steroid hormones (Couch and Hagino, 1983; Adiyodi, 1985). In several mud crab species such as S. serrata and S. olivacea, the color of the immature ovaries is translucent to yellowish and becomes darker yellow to dark orange in mature ovaries due to accumulation of yolk in the oocytes (Quinitio and Parado-Estepa, 2003). In the present study, the color of the ovaries were just the same with the previous studies of the ovaries development in S. serrata and S. olivacea, but differ with one species of mud crab which is S. paramamosain. The phenomena of changing coloration in ovaries due to accumulation of yolk in the oocytes which was used for the developing of the embryo.
From the previous study by Quinitio and Parado-Estepa (2003) of ovarian development of S. serrata as the benchmark for this present study. Oocyte diameter for stage 1 of was from 45-100 μm. For stage 2, stage 3 and stage 4 was 80-150, 120-200 and 150-250 μm, respectively. Oocyte diameter of female of P. pelagicus was much bigger compare to oocyte of the S. serrata. This difference may be due to the diet intake of the crab, habitat and the different of the species. The result of the present study showed that ovarian maturation cannot be determined precisely by morphological observation of the abdomen. However, the initiation of vitellogenesis could be determined through the external observation of the ovary without examining ovarian stage histologically. Therefore, external observation of the ovarian and gonad color would enables crab growers, aquaculturist, fishermen and fisheries managers to reduce time and cost for determining whether female P. pelagicus reaches ovarian maturity.
CONCLUSION
From the study, the fecundity of P. pelagicus increases with the increasing of CW. Female P. pelagicus with bigger CW produced more eggs compare to female with smaller BW. The results showed that the fecundity estimates range from 105443.333±35448.075 eggs with brood size highly correlated to CW and BW. The present study also determined the crab maturity through ovarian histological assessment which is easy to observe the maturity. There are four color of gonad in every stages ranging from milky white in color for stage 1, yellowish during stage 2, orange in stage 3 and finally the ovary turned to dark orange when the ovary are well developed. The size of the oocytes in the ovaries increase with the increasing of maturity stage.
REFERENCES
- Arshad, A., Efrizal, M.S. Kamarudin and C.R. Saad, 2006. Study on fecundity, embryology and larval development of blue swimming crab Portunus pelagicus (Linnaeus, 1758) under laboratory conditions. Res. J. Fish. Hydrobiol., 1: 35-44.
Direct Link - Batoy, C.B., J.F. Sarmago and B.C. Pilapil, 1987. Breeding season, sexual maturity and fecundity of the blue crab, Portunus pelagicus (L.) in selected coastal waters in leyte and vicinity, Philippines. Ann. Trop. Res., 9: 157-177.
Direct Link - Bhat, B.A., S. Ravichandran and S.A. Allayie, 2011. Influence of the eyestalk hormones on the metabolism and ionic regulation of the Crab Portunus pelagicus (Lineaus, 1857). J. Biol. Sci., 11: 203-209.
CrossRef - Brante, A., M. Fernandez, L. Eckerle, F. Mark, H.O. Portner and W. Arntz, 2003. Reproductive investment in the crab Cancer setosus along a latitudinal cline: Egg production, embryo losses and embryo ventilation. Mar. Ecol. Prog. Ser., 251: 221-232.
Direct Link - Cheng, Y., S. Li, G. Wang, X. Chen and Q. Lin, 2002. Structural modulation of the area between oocytes and follicular cells during vitellogenesis of the mud crab (Scylla serrata). Acta Zool. Sin., 48: 80-92.
Direct Link - De Lestang, S., N.G. Hall and I.C. Potter, 2003. Reproductive biology of the blue swimmer crab (Portunus pelagicus, Decapoda: Portunidae) in five bodies of water on the West coast of Australia. Fish. Bull., 101: 745-757.
Direct Link - De Lestang, S., N. Hall and I.C. Potter, 2003. Influence of a deep artificial entrance channel on the biological characteristics of the blue swimmer crab Portunus pelagicus in a large microtidal estuary. J. Exp. Mar. Biol. Ecol., 295: 41-61.
CrossRef - Hamasaki, K., K. Fukunaga and K. Maruyama, 2003. Egg development and incubation period of swimming crab Portunus trituberculatus (Decapoda: Portunidae) reared in the laboratory. Crustacean Res., 32: 45-54.
Direct Link - Hamasaki, K., H. Imai, N. Akiyama and K. Fukunaga, 2004. Ovarian development and induced ovipositon of the overwintering swimming crab Portunus trituberculatus (Brachyura: Portunidae) reared in the laboratory. Fish. Sci., 70: 988-995.
CrossRef - Hamasaki, K., K. Fukunaga and S. Kitada, 2006. Batch fecundity of the swimming crab Portunus trituberculatus (Brachyura: Portunidae). Aquaculture, 253: 359-365.
CrossRefDirect Link - Hamasaki, K., Y. Obata, S. Dan and S. Kitada, 2011. A review of seed production and stock enhancement for commercially important portunid crabs in Japan. Aquacult. Int., 19: 217-235.
CrossRefDirect Link - Ikhwanuddin, M., M.L. Shabdin and A.B. Abol-Munafi, 2009. Size at maturity of blue swimming crab (Portunus pelagicus) found in Sarawak coastal water. J. Sustainability Sci. Manage., 4: 56-65.
Direct Link - Ikhwanuddin, M., M.N. Azra, M.A.D. Talpur, A.B. Abol-Munafi and M.L. Shabdin, 2012. Optimal water temperature and salinity for production of blue swimming crab, Portunus pelagicus 1st day juvenile crab. AACL Bioflux, 5: 4-8.
Direct Link - Ikhwanuddin, M., M.N. Azra, Y.Y. Sung, A.B. Abol-Munafi and M.L. Shabdin, 2012. Live foods for Juveniles' production of blue swimming crab, Portunus pelagicus (Linnaeus, 1766). J. Fish. Aquat. Sci., 7: 266-278.
CrossRefDirect Link - Ikhwanuddin, M., A.H. Nurfaseha, A.B. Abol-Munafi and M.L. Shabdin, 2012. Movement patterns of Blue Swimming Crab, Portunus Pelagicus, the Sarawak Coastal Water, South China Sea. J. Sustainability Sci. Manage., 7: 8-15.
Direct Link - Islam, M.S., K. Kodama and H. Kurokura, 2010. Ovarian development of the mud crab Scylla paramamosain in a tropical mangrove swamps, Thailand. J. Sci. Res., 2: 380-389.
Direct Link - Johnson, D.D., C.A. Gray and W.G. Macbeth, 2010. Reproductive biology of Portunus pelagicus in a South-East Australian estuary. J. Crustacean Biol., 30: 200-205.
CrossRefDirect Link - Samuel, N.J. and P. Soundarapandian, 2009. Embryonic development of commercially important portunid crab Portuns sanguinolentus (Herbst). Int. J. Anim. Vet. Adv., 1: 32-38.
Direct Link - Josileen, J., 2011. Morphometrics and length-weight relationship in the blue swimmer crab, Portunus pelagicus (Linnaeus, 1758) (Decapoda, Brachyura) from the Mandapam coast, India. Crustaceana, 84: 1665-1681.
Direct Link - Kumar, M.S., Y. Xiao, S. Venema and G. Hooper, 2003. Reproductive cycle of the blue swimmer crab, Portunus pelagicus, off Southern Australia. J. Mar. Biol. Assoc. UK., 83: 983-994.
CrossRefDirect Link - Lai, J.C.Y., P.K.L. Ng, and P.J.F. Davie, 2010. A revision of the Portunus pelagicus (Linnaeus, 1758) species complex (Crustacea: Brachyura: Portunidae), with the recognition of four species. Raffles Bull. Zool., 58: 199-237.
Direct Link - Lee, H.H. and C.C. Hsu, 2003. Population biology of the swimming crab Portunus sanguinolentus in the waters off Northen Taiwan. J. Crustacean Biol., 23: 691-699.
Direct Link - Mohajeri, J., M. Afsharnasab, B. Jalali, S. Kakoolaki, M. Sharifrohani and A. Haghighi, 2011. Immunological and histopathological changes in Penaeus semisulcatus challenged with Vibrio harveyi. Iran. J. Fish. Sci., 10: 254-265.
Direct Link - Nitiratsuwan, T., C. Nitithamyong, S. Chiayvareesajja and B. Somboonsuke, 2010. Distribution of blue swimming crab (Portunus pelagicus Linnaeus, 1758) in Trang province. Songklanakarin J. Sci. Technol., 32: 207-212.
Direct Link - Rasheed, S. and J. Mustaquim, 2010. Size at sexual maturity, breeding season and fecundity of three-spot swimming crab Portunus sanguinolentus (Herbst, 1783) (Decapoda, Brachyura, Portunidae) occurring in the coastal waters of Karachi, Pakistan. Fish. Res., 103: 56-62.
CrossRefDirect Link - Ravi, R., M.K. Manisseri and N.K. Sanil, 2011. Ovarian maturation and oogenesis in the blue swimmer crab, Portunus pelagicus (Decapoda: Portunidae). Acta Zool.
CrossRef - Rejeki, S., 2007. The effects of different water flow rates on the survival rate of blue crab (Portunus pelagicus) zoea IV-megalopa stages. J. Coastal Dev., 10: 197-203.
Direct Link - Sahoo, D., S. Panda and B.C. Guru, 2011. Studies on reproductive biology and ecology of blue swimming crab Portunus pelagicus from Chilika Lagoon, Orissa, India. J. Mar. Biol. Assoc. UK., 91: 257-264.
CrossRefDirect Link - Soundarapandian, P., E. Thamizhazhagan and N.J. Samuel, 2007. Seed production of commercially important blue swimming crab Portunus pelagicus (Linnaeus). J. Fish. Aquatic Sci., 2: 302-309.
CrossRefDirect Link - Soundarapandian, P. and R.K. Singh, 2008. Biochemical composition of the eggs of commercially important crab Portunus pelagicus (Linnaeus). Int. J. Zool. Res., 4: 53-58.
CrossRefDirect Link - Stewart, M.J., P. Stewart, N. Soonklang, V. Linthong, P.J. Hanna, W. Duan and P. Sobhon, 2010. Spermatogenesis in the blue swimmer crab, Portunus pelagicus, and evidence for histones in the mature sperm nuclei. Tissue Cell., 42: 137-150.
Direct Link - Wu, X., B. Zhou, Y. Cheng, C. Zeng, C. Wang and L. Feng, 2010. Comparison of gender differences in biochemical composition and nutritional value of various edible parts of the blue swimmer crab. J. Food Composition Anal., 23: 154-159.
CrossRefDirect Link - Xiao, Y. and M. Kumar, 2004. Sex ratio and probability of sexual maturity of females at size, of the blue swimmer crab, Portunus pelagicus Linneaus, off Southern Australia. Fish. Res., 68: 271-282.
CrossRefDirect Link - Robertson, W.D. and A. Kruger, 1994. Size at maturity, mating and spawning in the Portunid crab Scylla serrata (Forskal) in Natal, South Africa. Estuarine Coastal Shelf Sci., 39: 185-200.
CrossRefDirect Link