
M.M. Hossain
Department of Fisheries and Marine Bioscience, Jessore Science and Technology University, Jessore, Bangladesh
K. Kawai
Department of Aquaculture, Faculty of Agriculture, Kochi University, Nankoku Shi, Kochi 783-8502, Japan
S. Oshima
Department of Aquaculture, Faculty of Agriculture, Kochi University, Nankoku Shi, Kochi 783-8502, Japan
Pakistan Journal of Biological Sciences
Year: 2011 | Volume: 14 | Issue: 15 | Page No.: 755-767







ABSTRACT
Japanese eel Anguilla japonica were immunized with inactivated Edwardsiella tarda bacterin preparations {formalin-killed cells, FKC (0.4%), formalin with heat-killed cells, FHKC (0.1% and 70°C for 10 min), heat-killed cells, HKC (70°C for 15 min), potassium chloride-killed cells, KKC (0.6%), tannic acid-killed cells, TKC (0.9%), citric acid-killed cells, CAKC (0.9%), pressure-killed cells, PKC (600 psi for 5 min) and electric current-killed cells, ECKC (100 mA at 12 v DC for 5 sec) via intraperitoneal injection in order to develop adequate inactivating method. Immune parameters in the immunized eel were measured to compare responses to different bacterins. Generally, eel rose agglutinating antibody titer in the serum within 2 week and the maximum titer occurred at 6 weeks post immunization. Elevated and significantly higher titer was produced with the PKC of E. tarda than other bacterin preparations. An Enzyme Linked Immunosorbent Assay (ELISA), to determine specific anti-E. tarda antibody in the serum, also showed significantly higher antibody titer with PKC than the other antigen preparations. Bacteriostatic assay with serum and live E. tarda indicated significantly higher activity in the PKC-immunized fish. Immunization with PKC also showed the increased level of phagocytosis. PKC-inactivated vaccine at an immunization dose of 106 cells/fish induced high protection against experimental infection. Coincident with higher immune parameters and protection in the fish immunized with the PKC bacterin strongly suggested that pressure-killing is an effective inactivating method to develop an effective vaccine against edwardsiellosis.
PDF Abstract XML References Citation
Received: July 05, 2011;
Accepted: October 15, 2011;
Published: November 24, 2011
How to cite this article
M.M. Hossain, K. Kawai and S. Oshima, 2011. Immunogenicity of Pressure Inactivated Edwardsiella tarda Bacterin to Anguilla japonica (Japanese Eel). Pakistan Journal of Biological Sciences, 14: 755-767.
DOI: 10.3923/pjbs.2011.755.767
URL: https://scialert.net/abstract/?doi=pjbs.2011.755.767
DOI: 10.3923/pjbs.2011.755.767
URL: https://scialert.net/abstract/?doi=pjbs.2011.755.767
INTRODUCTION
Edwardsiella tarda, a Gram-negative, motile, flagellated and rod-shaped bacterium, is the causative agent of edwardsiellosis in many commercially important freshwater and marine fish such as channel catfish, eels, mullet, Chinook salmon, flounder, carp, tilapia and striped bass (Kwon et al., 2005). It causes septicemia with extensive skin lesions, affecting internal organs and leads to extensive losses in both freshwater and marine aquaculture (Lan et al., 2008).
E. tarda, antibiotic resistance has been reported widely in the world (Aoki and Takahashi, 1987; Aoki et al., 1989; DePaola et al., 1995). To limit the use of chemicals and antibiotics, vaccination is highly recommended (Chinabut and Puttinaowarat, 2005).
Vaccines consisting of various E. tarda antigen preparations have been tested in eels on a number of occasions (Salati et al., 1983; Salati and Kusuda, 1985, 1986). Although several attempts have been made to induce protection against E. tarda (Gutierrez and Miyazaki, 1994; Mekuchi et al., 1995) the protection efficiency was variable among the studies and the majority deals with practical questions of vaccine efficacy. To improve the effectiveness of vaccines, many alternatives including the use of formalin killed whole cell bacterin (Salati et al., 1983), cellular lipid (Salati and Kusuda, 1986) and lipopolysaccharides (Salati et al., 1987a) have been reported.
To examine primary immune response of eel against E. tarda infection, fish were immunized with formalin-killed E. tarda cells (Salati et al., 1983) are established and effective. It has been reported that, most of the available fish vaccines are inactivated with formalin (Huang et al., 2008; Dumrongphol et al., 2008), which induced early immunoglobulin M response and lower the antigenicity of the bacterium (Bachmann et al., 1993) also exerts little side effects (Inaba et al., 1973). Other than formalin killed vaccine is not yet likely to be established, though result recent social situation desired new and safe effective inactivating method for vaccine preparation.
This study attempted to compare immune responses in eel to an inactivated E. tarda bacterin administered by intraperitoneal (IP) injection. The efficacies of the vaccines prepared by different methods were also compared and the optimal conditions for i.p. immunization were established.
MATERIALS AND METHODS
Bacterial strain and growth conditions: E. tarda V-1 strain, originally isolated from kidney of diseased Japanese eel was used in this study as a representative antigen. The bacterial strain was pre-cultured for 24 h at 30°C in brain heart infusion (BHI, Difco) broth and was inoculated into 1000 mL brain heart infusion broth, cultured with shaking at 30°C for 18 h. The cells were harvested by centrifugation at 4000xg for 15 min at 4°C.
Selected bacterins: Bacterial inactivation activity determined by viable counts (Reed and Reed, 1948) to confirm the bacterial inactivation, agglutination (using rabbit-antiserum), sodium dodecyl sulfate polyacrylamide gel electrophoresis (SDS-PAGE) and Western blotting (data were not shown) were performed to considered final conditions in Table 1a and b.
Fish and vaccination: Japanese eels Anguilla japonica weighing 101.2±3.8 g (Mean±SD) were obtained successively from an eel farm in Yoshikawa at Kochi Prefecture, Japan, which had no previous occurrence of infection with E. tarda in this farm. For each group, before immunization, 60 (20x3) fish were allowed to adapt for 2 weeks in three individual tank (100 L-tank) separately, supplied with well-aerated flowing water at 25°C and fed with 0.5 mm commercial dried pellets (Nissui) corresponding to 3% of the fish body weight per day for the entire experiment.
To compare the efficacies, the fish were immunized by intraperitoneal injection with 100 μL volume containing 100 μg of FKC (formalin at 0.4%), FHKC (0.1% formalin with 70°C for 10 min), HKC (Heating at 70°C for 15 min), KKC (0.6% KCl), TKC (0.9% tannic acid), CAKC (0.9% citric acid), PKC (pressure at 600 psi for 5 min) and ECKC (electric current at 100 mA at 12v DC for 5 sec) adjusted to a density of 5x106 cell/fish. Control fish were similarly injected with the PBS equal to the volume of vaccine.
After immunization, fish were maintained at 25°C. An identical booster immunization was administered 4 weeks post-primary immunization. After 5 weeks post-primary immunization 6 eel from each group were bled from Vena caudalis using a syringe and immediately transferred to heparinised containers were assayed for agglutinating antibody titers. Then sera from 10 fish of each group were bled to assayed peripheral blood or phagocytosis individually. The other fish (20-21 fish/each group) were used in ELISA or bacteriostatic assay and 25 fish/each group of FKC, FHKC, CAKC, PKC and ECKC in challenge tests.
Agglutinating antibodies in fish sera: Agglutinating antibodies against E. tarda were evaluated in sera from fish at 2, 4, 6, 8 and 12 weeks post immunization. Fish were bled from the caudal vessels and the sera was heat treated (44°C, 20 min) to inactivate complement activity (Sakai, 1981). The blood was allowed to clot at 4°C for 24 h and serum collected and analyzed for agglutinating antibodies against E. tarda were determined by the microtitre method (Nowotny, 1969). A two-fold dilution series of 25 μL of each serum sample was made in PBS in wells of a 96 well microtitre plate. Then 25 μL of differently inactivated E. tarda killed-suspension containing 108 CFU mL-1 was added to each well. After 2 h at 25°C and overnight at 4°C incubation, titers were scored as the highest serum dilution showing agglutination.
Preparation of peripheral blood, head kidney and spleen leucocytes: Peripheral blood samples were taken from the caudal vein using sterile syringes containing 0.5 mL of 5,000 I.U. mL-1 sodium heparin. The sample was then diluted by adding 2 mL of salted phosphate buffer saline (sPBS). The sPBS was prepared by adding a 20% solution of sodium chloride (NaCl) to Phosphate Buffer Saline (PBS) in the ratio of 1:25. The final osmolarity of the sPBS was adjusted to 372 mOsm, the osmolarity of eel plasma. The sample was centrifuged at 220xg for 10 min, the supernatant discarded and the leucocyte rich upper layer of cells was added to 1 mL of new born calf serum (NBCS, Sigma-Aldrich). Both the spleen and the anterior kidney of each fish were aseptically removed and placed in plastic Petri dishes containing salted RPMI (sRPMI). The sRPMI was also prepared by adding 20% NaCl solution to RPMI in the ratio of 1:25. The cell suspensions were prepared by pressing and disruption of the tissues using a sterile syringe piston and then transferring the homogenates to sterile 15 mL conical tubes. After the crude sediment had settled, the supernatant containing the cells was centrifuged at 220xg for 10 min at 4°C and the cells were resuspended in 1 mL NBCS.
| Table 1a: | Summary for effective inactivator and kinetic parameters for inactivation of E. tarda (~5 min post inactivation) with different methods, initial bacterial count (log CFU mL-1 = 8.9) |
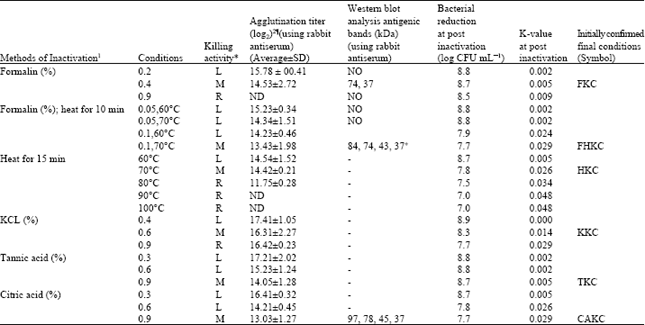 | |
| *L, low killing where bacteria still remain alive after 24 h, M, moderate killing in 24 h, where almost all bacteria were killed within 24 h, R, rapid killing in 6 or 12 h. Killing activity: Confirmation of bacterial inactivation was performed by the number of cells surviving for all treatments, immediately after the treatment ended (approximately 0, 6, 12, 18 and 24 h; incubation at 15°C) serial dilutions in 0.067 M phosphate buffer were prepared. The "drop plate" counting method was employed for colony counts (Reed and Reed, 1948). ¶Figures indicate the highest dilution times of the serum with positive agglutination. 2Geometric mean reciprocal log2 value of the highest dilution of the serum that showed positive agglutination. ND, agglutination value was not determined by autoagglutination. kDa, kilo Dalton. (-), indicates, no clear bands (remaining high antigenicity) were observed in the bacterial cells inactivated with above treatment. +37 kDa is the major protective antigen of this bacterium (Kawai et al., 2004). 1Initial concentrations of chemicals below 1% (from 0.1 to 0.9%) and chemicals combined with heat; initial heating 1, 5, 10, 15, 20 and 25 min for each condition were performed (data were not shown) **** | |
| Table 1b: | Summary for effective inactivator and kinetic parameters for inactivation of E. tarda (~5 min post inactivation) with different methods, initial bacterial count (log CFU mL-1 = 8.9) |
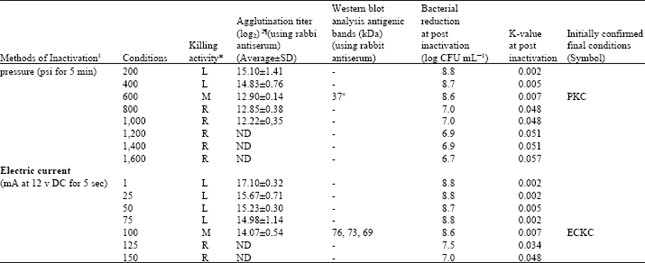 | |
| *L, low killing where bacteria still remain alive after 24 h, M, moderate killing in 24 h, where almost all bacteria were killed within 24 h, R, rapid killing in 6 or 12 h. Killing activity: Confirmation of bacterial inactivation was performed by the number of cells surviving for all treatments, immediately after the treatment ended (approximately 0, 6, 12, 18 and 24 h, incubation at 15°C) serial dilutions in 0.067 M phosphate buffer were prepared. The "drop plate" counting method was employed for colony counts (Reed and Reed, 1948). ¶Figures indicate the highest dilution times of the serum with positive agglutination. 2Geometric mean reciprocal log2 value of the highest dilution of the serum that showed positive agglutination. ND, agglutination value was not determined by autoagglutination. kDa, kilo Dalton. (-), indicates, no clear bands (remaining high antigenicity) were observed in the bacterial cells inactivated with above treatment. + 37 kDa is the major protective antigen of this bacterium (Kawai et al., 2004). 1 Initially 1, 2, 3, 4, 5 to 10 min were used to achieve the treatment pressure by the French pressure cell press, or French press (No 5501, OHTAKE Mfg, 941, Tokyo, Japan), decompression time was approximately 30-60 sec and current 1, 5, 10, 15, 20, 25 and 30 sec (Kevin et al., 2003) were applied using an electrolytic vessel with two electrodes was connected with electrophoresis machine for each condition) | |
The head kidney, spleen and Peripheral Blood Leucocytes (PBL) were separated on a Histopaque gradient composed of 1 mL of Histopaque 1.077 and 1 mL of Histopaque 1.119 (Sigma-Aldrich). The gradients were then centrifuged at 450xg for 25 min at 4°C, as described by Jeney et al. (1994). The leucocytes rich layer was collected, washed twice with sPBS and placed in a solution of sRPMI at a concentration of 5x106 cells mL-1. To the sRPMI were added 100 I.U. mL-1 penicillin, 100 μg mL-1 streptomycin, 0.3 mg mL-1 L (+) glutamine and 10% NBCS. Cell viability was analyzed by trypan blue (0.1%) with a haemocytometer (Law et al., 2001).
Macrophages isolation and phagocytosis assay: After homogenization of the head kidney and spleen, leucocytes were harvested by centrifugation then cell counts and viability were determined using a 0.5% trypan blue solution which was mixed 1:1 with an aliquot of the cell suspension. After cell concentration was adjusted a using haemocytometer. To determine percentages of macrophages cell suspensions were smeared onto glass slides, air dried and stained (Quick Dip(R); Histo Labs, River stone, NSW, Australia), then differential cell counts was performed as followed by previous study (Crosbie and Nowak, 2004).
Phagocytosis of the macrophage suspension was determined using a technique based on the previous study (Gravningen et al., 2008; Seeley et al., 1990). The number of head kidney macrophages and spleen macrophages were adjusted to 105 cells mL-1 in RPMI 1640 medium containing 10% fresh Eel Serum (ES). The cells were allowed to adhere to a coverslip for 1 h and loose cells were removed by washing with the medium. E. tarda bacterins (1x107 cells mL-1) were suspended in RPMI 1640 medium (10% ES) and were added to the coverslip and incubated for 2 h at 25°C. The cells on the coverslip were fixed with methyl alcohol and stained with Giemsa. The percent of phagocytic cells per 300 (phagocytes) cells was counted microscopically. Student’s t-test was used to evaluate the difference. The Phagocytic Index (PI) calculated by the following formula:
Enzyme-linked immunosorbent assay: A standard indirect ELISA was used to quantify specific serum antibody in eel after immunizations according to Crosbie and Nowak (2004). Microtitre plate wells (Iwaki, Scitech Div. Japan) were coated with formalin inactivated E. tarda sonicate (5.5 μg mL-1 protein) suspended in a coating solution (Sterile Carbonate Buffer, pH 9.5). Following overnight incubation at 4-5°C, washed twice with 300 μL wash solution PBST {0.1 M phosphate-buffered saline or Tris-buffered saline (pH 7.4) with Tween 20 (0.02-0.05% v/v)} and residual binding sites blocked with blocking buffer (1% bovine serum albumin and 0.05% Tween-20 in PBS) during incubation for 1 h at 37°C. Plates were washed twice with PBST, before the addition of eel serum samples, diluted in blocking buffer (1% bovine serum albumin and 0.05% Tween-20 in PBS), in duplicate wells across rows in a twofold serial dilution beginning at 1:50 through to 1:1600, incubated for 1 h at 37°C then washed again as above. A rabbit anti-eel serum, diluted 1:400 in blocking buffer, was then added to all wells and incubated for 1 h at 37°C. After repeating the washing step a goat anti-rabbit IgG (Gaithersburg MD 20878-USA) horseradish peroxidase (HRP) conjugate, diluted to 1:2000 in blocking buffer, was then added to all wells and plates were again incubated for 1 h at 37°C. Following a final wash step, the horseradish peroxidase was activated with 0.05% enzyme substrate p-nitrophenyl phosphate (Wako Chemicals USA, Inc). The reaction was stopped with 3 m HCl after 30 min and OD read with a microplate reader (Thermo Electron Corporation) at 492 nm. All reagents were added to wells in volumes of 50 μL and non-immune and immune standard sera were titrated in duplicate on each plate. The non-immune standard was a pool of sera from non-immunized fish and a standard anti-E. tarda antiserum (immune standard) was pooled antisera from several fish immunized by IP injection with 8 mg of the bacterin, then re-immunized after 4 weeks. All fish supplying standard sera had a mean weight of 101.2 g (SD 3.8) at the time of first immunization.
Anti-E. tarda activities of eel serum samples were determined according to Arkoosh and Kaattari (1990); Crosbie and Nowak (2004) with a method that equates an arbitrary unit of antibody activity to a volume of serum. The specific antibody activities of samples were expressed as units of antibody activity per volume of serum and were determined relative to the immune standard included on all plates and calculated as follows:
where, activity of the standard is the reciprocal of the volume of the immune standard which gives 40% maximum OD, as determined from a titration curve and is expressed as units of activity per μL of serum, 40% volume of the standard is the volume of the immune standard giving 40% maximum OD and 40% volume of the sample is the sample volume giving 40% maximum OD.
Bacteriostatic assay: A modified bacteriostatic assay was employed in which E. tarda suspension was grown to a density of approximately 5x108 cells mL-1. Bacterial culture was initiated by inoculating 120 mL of BHI with a cryopreservation bead (stored at - 80°C) of the bacterium, grown overnight and then inoculated into fresh media (3 mL). After incubation at 30°C in a shaking water bath for 2-2.5 h the cultures were in logarithmic growth phase. Cell densities were approximated from a standard curve constructed from a plot of log10 of cell densities vs. absorbance at 550 nm measured during the growth phase of the bacteria. Seventy-five microlitres of this culture were incubated with 25 μL of sample sera in 96-well flat-bottom microtitre plates at 24°C for 1 h after which 100 μL of 3-(4,5-dimethylthiozol-2-yl)-2, 5-diphenyltetrazolium bromide; thiazol blue (MTT, 5 mg mL-1 H2O) diluted 1:10 in BHI was added to each well; OD was immediately read at 550 nm and again after 15 min in the dark at room temperature with shaking. Differences in OD of samples and pooled non-immune standard eel serum (ΔOD) were calculated and antibacterial activity was determined by the formula:
Results are expressed as a percentage inhibition of E. tarda relative to the non-immune pooled standard sera. Samples were assayed in triplicate and all plates had pooled immune and non-immune serum standards prepared as described for the ELISA.
Challenge test: Injection (i.p.) challenge tests were done 6 weeks post immunization and the control fish were challenged accompanied with five identical vaccine groups at the respective time points. The E. tarda dose for i.p. challenge test was 5x106 cells/fish and 5x103 cells/fish. Control fish that had been injected with PBS were challenged as well. The cumulated mortality was recorded 12 days post challenge. E. tarda was confirmed by analysis of kidney isolates cultured on Salmonella Shigella agar (SS agar, Nissui) and the infection of E. tarda was confirmed by observation of black pigments. Slide agglutination titer was also performed to confirmed E. tarda using eel antiserum mixed with bacterin on PBS; visible granular clumps (agglutination) were observed under the microscope.
Relative percentage survival, RPS (Croy and Amend, 1977) was calculated from the end cumulative mortalities as:
Inactivation kinetics of the E. tarda: Rahn suggested that microbial destruction was due to inactivation of a single critical molecule in the cell and, therefore, could be assumed to follow first-order kinetics (Rahn, 1945). Therefore, to evaluate the effects of different treatment on the rates of inactivation of bacterial spores, a simplified mathematical model based on first-order kinetics was used (Joslyn, 2001; Pflug et al., 2001).
where, N = microbial population at any time, t; N0 = initial microbial population (the average viable spore counts measured prior to the different treatment i.e., control); and at time t (t = time immediately after inactivation), respectively and k is the specific death rate constant or death rate coefficient which is a function of inactivation’s.
Statistical analysis: The data of antibody and immune responses between treatment groups were represented as Means±SD and analyzed by one-way Analysis of Variance (ANOVA) and Tukey’s test was used to compare the mean values between individual treatments (Zar, 1984) using Stat plus 2007 professional. The significance of the data from phagocytosis experiments was assessed using a Student's t-test; differences in mortality were tested for statistical significance by the Chi-square contingency table test with the Yates’ correction (Pagano and Gauvereau, 2000). Mean values were also considered significantly different at p<0.05 or p<0.001.
RESULTS
Serum antibody titer: Agglutinating antibodies were detected in all vaccinated groups at post immunization. Average antibody titer in those group elevated at 2 weeks post-immunization with a mean titer of PKC group was (14.64 log2) which was significantly greater than other vaccinated groups or controls (p<0.05). By 6 weeks post-immunization, mean titers remained significantly (p<0.05) elevated to peak with the highest titers reaching 18.64 log2 but decreased to 13.64 log2 with PKC group at 12 weeks post-immunization. The control fish had a low level of antibodies but on average, the control group was 8.83 log2 (Fig. 1). Fish from groups PKC and FKC were followed up to over the post vaccination and fish in both groups had an increased level of agglutinating antibodies at week 6 compared to earlier time points.
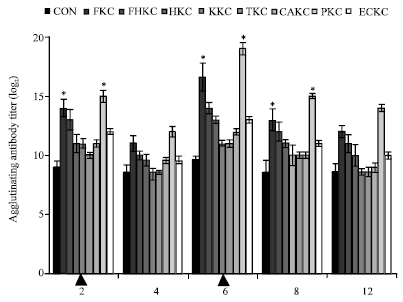 | |
| Fig. 1: | Changes in the agglutinating antibody titer of immunized and non-immunized eel sera, p: pre- immunization; Arrows (▲) mark primary and booster immunizations. Bars represent the agglutination antibody titer (Mean±SE; n = 6). Symbols denote statistically significant differences (*, p <0.05) between groups |
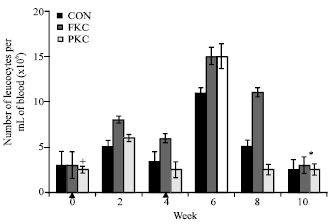 | |
| Fig. 2: | Total number of isolated peripheral blood leukocytes (PBLs) from eel immunized with PKC and FKC. Bars represent the Mean±SE (n = 4) of leucocytes per liter of blood (x106). Symbols denote statistically significant differences (*, p<0.05; +, p<0.05 and p>0.001) between groups. Arrows (▲) mark primary and booster immunizations |
Fish immunized with other treatments did not exhibit significant antibodies at any time.
Peripheral blood leukocytes: Firstly, we evaluated the changes in the number of PBLs resulting from the intraperitoneally injection of different E. tarda bacterins to eel. The studies demonstrated that, a significantly lower number of leucocytes were seen in the control group. The highest numbers of the cells were detecting in KKC group (Table 2). Mean number of cells with all treatment initially peaked at 2 weeks post-immunization and reached a maximum at 6 weeks post-immunization (Fig. 2). The studies showed that, the pools of leucocytes were subsided by 4 weeks and least abundant at 10 weeks post-immunization for all treatments. The intra-group differences, in content of leucocytes number with PKC group was significant (p<0.05).
Leukocyte populations: As shown Fig. 3, HKLp and SLLp were observed as the highest populations, while HKMo and SLMo showed low values over the experimental period. Status about general differences between (profiles, comparison) groups throughout period in Fig. 3.
Phagocytosis of macrophages: A significant increase in phagocytosis of head kidney macrophages was observed in the PKC vaccinated group (39.0±2.5) compared to controls (14.4 ±1.9) at 10 days (p<0.01); and (17.4±2.9) in vaccinated versus (6.6±1.4) in controls (p<0.05) at 21 days post vaccination (Table 3). Phagocytosis of spleen macrophages were also significantly (p<0.05) increase with PKC vaccinated group at 10 and 21 days post vaccination. As shown in Table 3, phagocytic index of head kidney and spleen macrophages suspensions with PKC group indicated higher macrophage activation value than other vaccinated group.
Specific antibody responses: Antibody responses were noted in all bacterin immunized groups after both primary and secondary immunizations (Fig. 4), however serum was not collected from four individuals after the primary immunization due to difficulties in sampling the fish.
| Table 2: | Changes the No. of total leucocytes (Mean±SD) in peripheral blood |
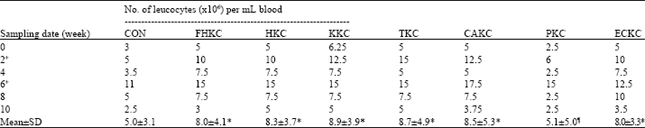 | |
| +Symbol denoted primary and booster immunizations; ¶Value is significantly different (p<0.05) from the corresponding all other values. *Data are not significantly different (p<0.05) from control group | |
| Table 3: | Head kidney and spleen macrophage phagocytic indices 10 and 21 days post-secondary immunizations with E. tarda bacterin via intraperitoneal (IP) injection |
 | |
| *Mean±SD, n = 3. †Data arc significantly different (p<0.05) from control group | |
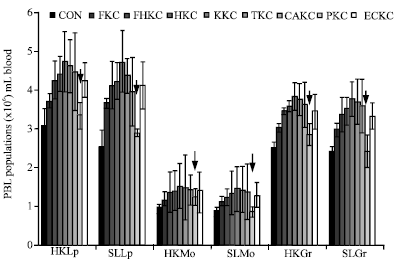 | |
| Fig. 3: | Leukocytes (PBLs) populations from eel with immunized with differently prepared bacterins; FKC (formalin at 0.4%), FHKC (0.1% formalin with 70°C for 10 min), HKC (Heating at 70°C for 15 min), KKC (0.6% KCl), TKC (0.9% tannic acid), CAKC (0.9% citric acid), PKC (pressure at 600 psi for 5 min) and ECKC (electric current at 100 mA at 12v DC for 5 sec). Symbol denotes (↓) statistically significant (p<0.05) between groups. Bar represents the mean number (x106) of leucocytes populations (Mean±SE; n = 6) in head kidney (HK) and spleen (SL) of eel. Lymphocytes (HKLp, SLLp), monocytes (HKMo, SLMo), granulocytes (HKGr, SLGr) at 10 weeks post immunization |
In some instances anti-E. tarda activity was noted in control (saline-immunized) fish, however this activity was only found in one individual fish after both primary and secondary immunizations. Analyses of antibody activity within groups 21 days after primary immunization showed that the bacterin-immunized groups were significantly higher than saline-treated controls.
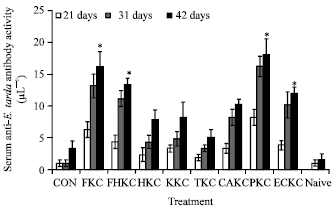 | |
| Fig. 4: | Specific anti-E. tarda antibody activity of eel sera after immunization with an experimental E. tarda bacterin or saline as a control (con) via intraperitoneal (IP) injection or naive to immunization and handling. Twenty-one days post-primary immunization antibody activities were determined for all groups (n = 21), then groups were reimmunized as before. Serum antibody activities were then determined for half the fish in all groups after 10 days (n = 10) and the other half after another 11 days (n = 10), i.e., at 31 and 42 days post-primary immunization. Statistical comparisons were: (1) each bacterin-treated group to appropriate control at 21, 31 and 42 days post-primary immunization; (2) bacterin treated groups to the naive groups treatment groups at 31 and 42 days post-primary immunization; ( (3) responses within groups at 21 and 31 days and then at 21 and 42 days post-primary immunization. Significant differences were PKC, FKC, FHKC and ECKC-treated groups to controls at 21, 31 and 42 days. (values are Mean±SD and *, p<0.05) |
Similar patterns of significantly higher antibody activity in bacterin-treated groups compared with saline controls were seen at 31 and 42 days post-primary immunization. Antigen-specific antibody activity was more consistent in the bacterin-treated groups with the higher percentages of fish responding whereas control groups had fewer responders (Table 4). However, this experiments indicates, the PKC group showed significantly (p<0.05) higher antibody response than other treated groups.
Bacteriostatic assay: Bacteriostatic activity of eel serum, expressed as percentage inhibition of E. tarda relative to pooled non-immune standard eel sera, was observed in all bacterin-immunized groups as well as in sera of some individuals in control groups (Fig. 5). This activity was, however, significantly higher in the PKC bacterin group with all individuals displaying substantial inhibition of E. tarda and 18 of 20 fish recording greater than 60%.
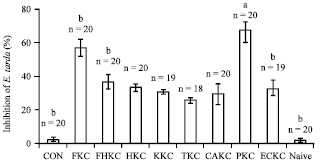 | |
| Fig. 5: | E. tarda inhibitory activity of eel serum (relative to pooled non-immune standard sera) after secondary immunization with E. tarda bacterin via intraperitoneal (IP) injection after secondary immunizations, N signifies groups naive to all treatments and handling. Data are Mean (±SD) at 10 and 21 days post-booster. Bars with asterisks are significantly different from all other groups (p<0.001) |
| Table 4: | Number of individual eel displaying anti-E. tarda antibody activity, as determined by ELISA and bacterial inhibitory activity in serum after immunizations with E. tarda bacterin or saline via intraperitoneal (IP) injection |
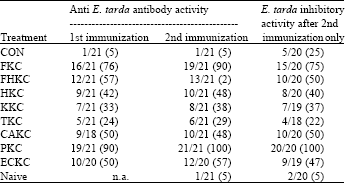 | |
| Values in parentheses are in percentage. n.a., Not applicable as naive group was only sampled at the end of the experiment | |
Inhibitory activity was generally much lower and more varied in other groups with 50% or less (Table 4) of individuals showing E. tarda inhibitory activity as assessed by this assay.
Efficacy of vaccines: To compare the efficacy of differently-inactivated vaccines, the final concentrations of the vaccines during injection (i.p.) immunization were adjusted to 106 cells/fish. There were no abnormalities in any of the vaccinated or control fish before challenge. The fish in each group were then injection (i.p.)-challenged 6 weeks post immunization. The cumulative mortality of the challenged fish was 56-72% for the control group, 16-28% for FKC group, 28-48% for FHKC group, 48-68% for CAKC group, 4-8% for PKC group and 40-64% ECKC group (Fig. 6 and 7).
| Table 5: | Efficacy of vaccines against Edwardseilla tarda in Japanese eel (Anguilla japonica) |
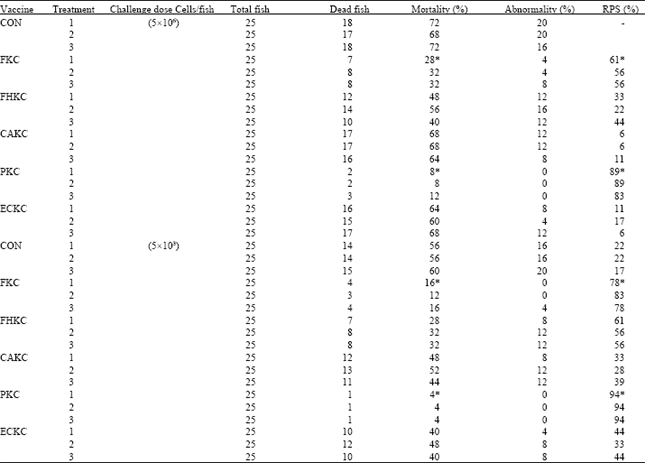 | |
| Asterisk: Significantly (p<0.05) lower than control | |
The RPS value of the fish immunized by PKC was 89-93, i.e. much higher than that of the fish immunized by FKC (RPS = 61-71) or other vaccinated group (Table 5). In addition, all efficacy trials the dead fish showed clinical signs typical of Edwardsiellosis disease. No pathogen other than E. tarda was isolated from dead fish.
Inactivation kinetics: The kinetic parameters for inactivation of microbial populations due to different inactivation methods appear to have a significant influence on inactivation rates for populations of E. tarda, are presented in Table 1a and b.
DISCUSSION
Present new approach to produce an inactivated E. tarda vaccine is conducted with considering the awareness of the residual effect of inactivation to human is based on the low concentration of formalin combined with heat, different chemicals, some fatty acids, heat, pressure and low amperage electric current other than high concentration of formalin inactivation of bacterial cells.
In eels, it has been demonstrated that high agglutinating antibodies titer or humoral immune response was obtained with intramuscular injection of the FKCs or crude lipopolysaccharide (LPS) of E. tarda (Salati and Kusuda, 1985; Salati et al., 1987a) respectively. In the present study, the bacterins of E. tarda, generated a strong serum antibody response in eel via intraperitoneal injection. Significantly higher serum agglutination titers detected in PKC groups shows that PKC remain antigen in a good state.
The apparent anti-E. tarda antibody activity noted in some individuals in control groups may be due to previous exposure of controls to the bacterium, cross-reactivity of existing antibodies to E. tarda antigens or inaccuracies and lack of specificity of the ELISA. Serum anti-E. tarda antibodies measured by ELISA was useful in evaluating the immunigenecity of the bacterins have been reported against Gram-negative bacteria in fish as well as in mammals (Salati et al., 1987b; Bricknell et al., 1997; Gudmundsdottir et al., 2003; Bridle et al., 2002). However, ELISA data in our study also indicated that the immune response in vaccinated eel correlated well with the protection efficacy.
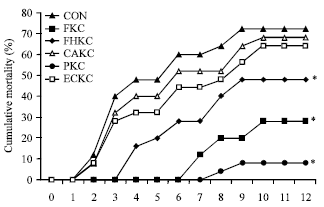 | |
| Fig. 6: | Cumulative mortality of Japanese eel (Anguilla japonica) after intraperitoneal injection challenge of E. tarda. Eels were separately injection (i.p.)-immunized with formalin-killed cell(0.4% formalin, FKC), formalin and heat-killed cells (0.1% formalin and 70°C for 10 min, FHKC), citric acid-killed cell (0.9% citric acid, CAKC), pressure-killed cell (600 psi/5 min, PKC), low amperage electric current-killed cell (100 mA/5 sec/12v DC, ECKC) with bacterial concentration of 106 cells/fish, or control group (CON) with PBS only and then intraperitoneal injection-challenged with E. tarda (5x106 cells/fish) 6 weeks post immunization. The mortalities reached 72% by day 12 post-challenge for the groups injected with a high dose. *p<0.001 (Chi-square) |
An interesting finding of this study was that all the bacterin-treated fish showed a serum antibody response and these sera showed substantial E. tarda inhibitory activity regardless of the magnitude of specific antibody activity. This above finding may indicate that the bacterin elicited a potentially protective response, at least in the specific humoral arm of the immune system (Yano, 1996).
The effects of other micoorganisms or lymphocyte disorders can often be estimated by counting the number of leucocytes present in the blood. Experimentally in infected fish, or the presence of particulate antigens, such us bacteria, parasites or tumor cells it has been observed the fluctuation of leukocytes from peripheral blood, head kidney and spleen tissues (Pressley et al., 2005; Chin and Woo, 2005).
To differentiate some of the leukocyte populations and their variations, morpho-functional studies (Sepulcre et al., 2002; Cuesta et al., 2003) has been carried out. Variations in the experimental designs, including temperature, photoperiod regime and fish size, all factors known to affect the immune system (Slater and Schreck 1998; Harrahy et al., 2001).
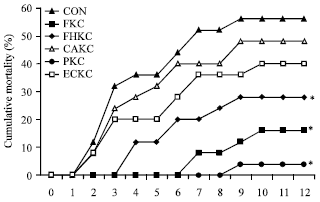 | |
| Fig. 7: | Cumulative mortality of Japanese eel (Anguilla japonica) after intraperitoneal injection challenge of E. tarda. Eels were separately injection (i.p.)-immunized with formalin-killed cell(0.4% formalin, FKC), formalin and heat-killed cells (0.1% formalin and 70°C for 10 min, FHKC), citric acid-killed cell (0.9% citric acid, CAKC), pressure-killed cell (600 psi/5 min, PKC), low amperage electric current-killed cell (100 mA/5 sec/12v DC, ECKC) with bacterial concentration of 106 cells/fish, or control group (CON) with PBS only and then intraperitoneal injection-challenged with E. tarda (5x103 cells/fish) 6 weeks post immunization. The mortalities reached 56% by day 12 post-challenge for the groups injected with a high dose. *p<0.001 (Chi-square) |
In the present study, the notable changes in numbers of leucocytes in peripheral blood take place during at post immunization and that marked changes are associated with booster immunization, which was not influenced by fish growth (data were not shown). It was also stated that, fluctuations of leucocytes populations, maturation status and presence of particulate antigens elicited activation of adaptive or innate immune system to produce specific immune responses to the antigen (Evans et al., 2000; Ellis, 2001; Cuesta et al., 2003, 2007).
Observed considerable variation in this study within groups and individual eel tested on the same and different days, which interfered with the interpretation of results in immune responses. It was interesting that increased number of leucocytes were detecting in KKC group until the end of the experimental period. Although number of cells reached a maximum at 6 weeks post-immunization with all treatment, the distribution of leukocyte populations of peripheral blood, only gradual changes was observed until the end of the experimental period with in PKC and FKC group. This finding indicates that leucocytes fluctuations may not directly correlated fish defense mechanisms, sometimes increased numbers create affect to the normal immune defense mechanisms or lymphocyte disorders (Pettersen et al., 2003). The notable changes observed in the KKC group, indicates some properties of leukocytes may change due to activation with the presence of immature cells.
Evidence of protective efficacy in eel by IP vaccination, antigenicity superiority was found in PKC group (RPS = 89-93), whereas the RPS values of FKC were 61-71 which happily avoids morbidity of eels encourages further investigation of this vaccine in several species. Protective efficacy in fish with E. tarda bacterin against edwardsiellosis has been reported by Ashida et al. (1999), Kwon et al. (2005, 2007), Swain et al. (2007) and Castro et al. (2008), respectively.
The results presented here show that eel respond systemically to E. tarda bacterin when it is delivered via IP injection exposure to leucocytes and its population rapidly recruited to the injection site and increased phagocytosis or bacteriostatic activity and that the response was highest in PKC group. PKC demonstrated that a single dose of the vaccine was sufficient to induce an immune response and prevent disease in eel.
ACKNOWLEDGMENTS
We would like to thank the staff and students of Fish Disease Laboratory, Faculty of agriculture, Kochi University. Japan.
REFERENCES
- Ashida, T., E. Okimasu, M. Ui, M. Heguri, Y. Oyama and A. Amemura, 1999. Protection of Japanese flounder Paralichthys olivaceus against experimental edwardsiellosis by formalin-killed Edwardsiella tarda in combination with oral administration of immunostimulants. Fish. Sci., 65: 527-530.
CrossRefDirect Link - Bridle, A.R., P.B.B. Crosbie, R.N. Morrison, J. Kwang and B.F. Nowak, 2002. The immuno-adjuvant effect of carp interleukin-1 β on the humoral immune response of barramundi Lates calcarifer (Bloch). J. Fish Dis., 25: 429-432.
CrossRef - Castro, N., A.E. Toranzo, S. Nunez and B. Magarinos, 2008. Development of an effective Edwardsiella tarda vaccine for cultured turbot (Scophthalmus maximus). Fish Shellfish Immunol., 25: 208-212.
Direct Link - Chin, A. and P.T. Woo, 2005. Innate cell-mediated immune response and peripheral leukocyte populations in Atlantic salmon, Salmo salar L., to a live Cryptobia salmositica vaccine. Parasitol. Res., 95: 299-304.
PubMed - Chinabut, S. and S. Puttinaowarat, 2005. The choice of disease control strategies to secure international market access for aquaculture products. Dev. Biol., 121: 255-261.
Direct Link - Crosbie, P.B.B. and B.F. Nowak, 2004. Immune responses of barramundi, Lates calcarifer (Bloch), after administration of an experimental Vibrio harveyi bacterin by intraperitoneal injection, anal intubation and immersion. J. Fish Dis., 27: 623-632.
Direct Link - Cuesta, A., A. Rodry guez, I. Salinas, J. Meseguer and M. N. Esteban, 2007. Early local and systemic innate immune responses in the teleost gilthead seabream after intraperitoneal injection of whole yeast cells. Fish Shellfish Immunol., 22: 242-251.
PubMed - Cuesta, A., M.A. Esteban and J. Meseguer, 2003. Identification of a FasL-like molecule in leukocytes of the teleost fish gilthead seabream (Sparus aurata L.). Dev. Comp. Immunol., 27: 21-27.
PubMed - DePaola, A., J.T. Peeler and G.E. Rodrick, 1995. Effect of oxytetracycline-medicated feed on antibiotic resistance of gram-negative bacteria in catfish ponds. Applied Environ. Microbiol., 61: 2335-2340.
PubMedDirect Link - Dumrongphol, Y., T. Hirota, H. Kondo, T. Aoki and I. Hirono, 2008. Identification of novel genes in Japanese flounder (Paralichthys olivaceus) head kidney up-regulated after vaccination with Streptococcus iniae formalin-killed cells. Fish Shellfish Immunol., 26: 197-200.
CrossRef - Ellis, A.E., 2001. Innate host defense mechanisms of fish against viruses and bacteria. Dev. Comp. Immunol., 25: 827-839.
CrossRefDirect Link - Evans, D.L., S.L. Taylor, J.H. Leary, G.R. Bishop, A. Eldar and L. Jaso-Friedmann, 2000. In vivo activation of tilapia nonspecific cytotoxic cells by Streptococcus iniae and amplification with apoptosis regulatory factor(s). Fish Shellfish Immunol., 10: 419-434.
PubMed - Gravningen, K., M. Sakai, T. Mishiba and T. Fujimoto, 2008. The efficacy and safety of an oil-based vaccine against Photobacterium damsela subsp. Piscicida in yellowtail (Seriola quinqueradiata): A field study. Fish Shellfish Immunol., 24: 523-529.
PubMed - Gudmundsdottir, S., S. Lange, B. Magnadottir and B.K. Gudmundsdottir, 2003. Protection against atypical furunculosis in Atlantic halibut, Hippoglossus hippoglossus (L.); comparison of a commercial furunculosis vaccine and an autogenous vaccine. J. Fish Dis., 26: 331-338.
PubMed - Gutierrez, M.A. and T. Miyazaki, 1994. Responses of Japanese eels to oral challenge with Edwardsiella tarda after vaccination with formalin-killed cells or lipopolysaccharide of the bacterium. J. Aquat. Anim. Health, 6: 110-117.
CrossRefDirect Link - Harrahy, L.N.M., C.B. Schreck and A.G. Maule, 2001. Antibody-producing cells correlated to body weight in juvenile chinook salmon (Oncorhynchus tshawytscha) acclimated to optimal and elevated temperatures. Fish Shellfish Immunol., 11: 653-659.
CrossRef - Huang, Y., H. Lou, X. Wu and Y. Chen, 2008. Characterization of the BPI-like gene from a subtracted cDNA library of large yellow croaker (Pseudosciaena crocea) and induced expression by formalin-inactivated Vibrio alginolyticus and Nocardia seriolae vaccine challenges. Fish Shellfish Immunol., 25: 740-750.
PubMed - Inaba, Y., H. Kurogi, K. Sato, Y. Goto, T. Omori and M. Matumoto, 1973. Formalin-inactivated, aluminum phosphate gel-adsorbed vaccine of bovine ephemeral fever virus. Arch. Virol., 42: 42-53.
CrossRefDirect Link - Kawai, K., Y. Liu, K. Ohnishi and S.I. Oshima, 2004. A conserved 37 kDa outer membrane protein of Edwardsiella tarda is an effective vaccine candidate. Vaccine, 22: 3411-3418.
CrossRefDirect Link - Kevin, P.D., A. Morteza and M.M. Raina, 2003. Comparative electrochemical inactivation of bacteriaand bacteriophage. Water Res., 37: 2291-2300.
Direct Link - Kwon, S.R., E.H. Lee, Y.K. Nam, S.K. Kim and K.H. Kim, 2007. Efficacy of oral immunization with Edwardsiella tarda ghosts against edwardsiellosis in olive flounder (Paralichthys olivaceus). Aquaculture, 269: 84-88.
CrossRefDirect Link - Kwon, S.R., Y.K. Nam, S.K. Kim, D.S. Kim and K.H. Kim, 2005. Generation of Edwardsiella tarda ghosts by bacteriophage PhiX174 lysis gene E. Aquaculture, 250: 16-21.
Direct Link - Law, W.Y., W.H. Chen, Y.L. Song, S. Sufour and C.F. Chang, 2001. Differential in vitro suppressive effects of steroids on leukocyte phagocytosis in two teleosts, tilapia and common carp. Gen. Comp. Endocr., 121: 163-172.
CrossRef - Pettersen, E.F., M. Ulvenes, G.O. Melingen and H.I. Wergeland, 2003. Peripheral blood and head kidney leucocyte populations during out-of-season (0+) parr-smolt transformation and seawater transfer of Atlantic salmon (Salmo salar L.). Fish Shellfish Immunol., 15: 373-385.
PubMed - Pressley, M.E., P.E. Phelan, P.E. Witten, M.T. Mellon and C.H. Kim, 2005. Pathogenesis and inflammatory response to Edwardsiella tarda infection in the zebrafish. Dev. Comp. Immunol., 29: 501-513.
PubMed - Reed, R.W. and G.B. Reed, 1948. Drop plate method of counting viable bacteria. Can. J. Res., 26: 317-326.
CrossRefDirect Link - Salati, F., M. Hamaguchi and R. Kusuda, 1987. Immune response of red sea bream to Edwardsiella tarda antigens. Fish Pathol., 22: 93-98.
Direct Link - Seeley, P.D., D. Gillespie and B.A. Weeks, 1990. A simple technique for the rapid spectrophotometric determination of phagocytosis by fish macrophages. Mar. Environ. Res., 30: 37-41.
CrossRef - Sepulcre, P., P. Pelegryn, V. Mulero and J. Meseguer, 2002. Characterisation of gilthead seabream acidophilic granulocytes by a monoclonal antibody unequivocally points to their involvement in fish phagocytic response. Cell Tissue Res., 308: 97-102.
PubMed - Slater, C.H. and C.B. Schreck, 1998. Season and physiological parameters modulate salmonid leucocyte androgen receptor affinity and abundance. Fish Shellfish Immunol., 8: 379-391.
Direct Link - Swain, P., A. Behura, S. Dash and S.K. Nayak, 2007. Serum antibody response of Indian major carp, Labeo rohita to three species of pathogenic bacteria; Aeromonas hydrophila, Edwardsiella tarda and Pseudomonas fluorescens. Vet. Immunol. Immunopathol., 117: 137-141.
Direct Link - Lan, J., X.H. Zhang, Y. Wang, J. Chen and Y. Han, 2008. Isolation of an unusual strain of Edwardsiella tarda from turbot and establish a PCR detection technique with the gyrB gene. J. Applied Microbiol., 105: 644-651.
PubMedDirect Link - Croy, T.R. and D.F. Amend, 1977. Immunization of sockeye salmon (Oncorhynchus nerka) against vibriosis using the hyperosmotic infiltration technique. Aquaculture, 12: 317-325.
CrossRef