
Jigita Padhiar
Department of Microbiology, Genohelix Biolabs A Division of Centre for Advanced Studies in Biosciences, Jain University, India
Arijit Das
Department of Microbiology, Genohelix Biolabs A Division of Centre for Advanced Studies in Biosciences, Jain University, India
Sourav Bhattacharya
Department of Microbiology, Genohelix Biolabs A Division of Centre for Advanced Studies in Biosciences, Jain University, India
Pakistan Journal of Biological Sciences
Year: 2011 | Volume: 14 | Issue: 22 | Page No.: 1011-1018







ABSTRACT
The present study was aimed at optimization, production and partial purification of lipases from Pseudomonas aeruginosa, Candida albicans and Aspergillus flavus. Various nutritional and physical parameters affecting lipase production such as carbon and nitrogen supplements, pH, temperature, agitation speed and incubation time were studied. Refined sunflower oil (1% v/v) and tryptone at a pH of 6.2 favored maximum lipase production in Pseudomonas at 30°C and 150 rpm, when incubated for 5 days. In C. albicans refined sunflower oil (3% v/v) and peptone resulted in maximum lipase production at pH 5.2, 30°C and 150 rpm, when incubated for 5 days. In A. flavus coconut oil (3% v/v) and peptone yielded maximum lipase at pH 6.2, 37°C, 200 rpm after an incubation period of 5 days. The lipases were partially purified by ammonium sulphate precipitation and dialysis. In P. aeruginosa enzyme activity of the dialyzed fraction was found to be 400 U mL-1 and for C. albicans 410 U mL-1. The dialysed lipase fraction from A. flavus demonstrated an activity of 460 U mL-1. The apparent molecular weights of the dialyzed lipases were determined by sodium dodecyl sulphate polyacrylamide gel electrophoresis (SDS-PAGE). The dialyzed lipase fraction obtained from P. aeruginosa revealed molecular weights of 47, 49 and 51 kDa, whereas, lipases from C. albicans and A. flavus demonstrated 3 bands (16.5, 27 and 51 kDa) and one band (47 kDa), respectively. These extracellular lipases may find wide industrial applications.
PDF Abstract XML References Citation
Received: August 04, 2011;
Accepted: October 14, 2011;
Published: December 01, 2011
How to cite this article
Jigita Padhiar, Arijit Das and Sourav Bhattacharya, 2011. Optimization of Process Parameters Influencing the Submerged Fermentation of Extracellular Lipases from Pseudomonas aeruginosa, Candida albicans and Aspergillus flavus. Pakistan Journal of Biological Sciences, 14: 1011-1018.
DOI: 10.3923/pjbs.2011.1011.1018
URL: https://scialert.net/abstract/?doi=pjbs.2011.1011.1018
DOI: 10.3923/pjbs.2011.1011.1018
URL: https://scialert.net/abstract/?doi=pjbs.2011.1011.1018
INTRODUCTION
Lipases (triacylglycerol acyl hydrolases EC 3.1.1.3) constitute a class of enzymes which catalyze the hydrolysis of lipids (triacylglycerols) yielding free fatty acids, diacylglycerols, monoacylglycerols and glycerol (Aravindan et al., 2007). A variety of chemical reactions such as hydrolysis, alcholysis, acidolysis, esterification and aminolysis can be catalyzed by lipases (Liu and Zhang, 2011). Lipases are synthesized by many biological sources such as plants, animals and microorganisms. Recently, microbial lipases have gained significant attention due to their wide applicability in various industrial processes (Anbu et al., 2011). Lipases are being industrially exploited due to the availability of inexpensive extraction methods, their stability over a broad range of temperature and pH, substrate specificity and activity even in the presence of organic solvents (Aravindan et al., 2007). Lipases are widely used in organic industries for synthesis and biotransformations (Gitlesen et al., 1997). They are also extensively used in the formulation of detergents, leather industry, environmental management, cosmetics and perfume industry (Pandey et al., 1999). The role of lipases also extends to food and pharmaceutical industries (Gulati et al., 2005).
Most of the industrial microbial lipases are derived from fungal and bacterial sources (Sharma et al., 2001). Among various Gram-positive and Gram-negative lipolytic bacterial species, lipases from the genus Bacillus and Pseudomonas are quite significant (Jaeger et al., 1994). Fungi are also important producers of extracellular lipases. Among the filamentous fungi, species of Aspergillus are among the most widely known prolific lipase producers and their enzymes are suitable for various industrial applications (Fu et al., 1995; Rani and Panneerselvam, 2009; Jayaprakash and Ebenezer, 2010). Interestingly, lipase production from some yeasts such as Candida sp. has also been documented (Rajendran and Thangavelu, 2007).
The ever-increasing industrial demand for lipolytic enzymes has triggered renewed interest and a constant search for new enzyme sources. Although many lipases have already been reported, novel lipases with better substrate specificity, greater thermal and pH stability and higher activity are being thoroughly investigated (Mateos Diaz et al., 2006). Inspite of the presence of well documented reports on the screening and selection of lipase producers, very less information is available on the fermentation process (Fadilo-Lu and Erkmen, 2002).
The rate of lipase production by various microorganisms may be enhanced with the incorporation of specific lipase inducers such as olive oil and organic nitrogen supplements into the production media (Nesbit and Gunasekaran, 1993). It has also been found that lipase activity is dependent on the source and type of substrate used (Vakhlu and Kour, 2006). This comparative study was aimed at investigation of the process parameters influencing the production of extracellular lipases from Pseudomonas aeruginosa, Candida albicans and Aspergillus flavus.
MATERIALS AND METHODS
The present study was conducted during the period from 18.05.2011 to 27.09.2011 at Genohelix Biolabs, A Division of Centre for Advanced Studies in Biosciences, Jain University, Chamarajpet, Bangalore, Karnataka, India.
Chemicals and reagents: All the media used during the course of the study were obtained from Himedia Laboratories Pvt. Limited (Mumbai, India). The analytical grade chemicals and reagents were purchased from Loba Chemie (Mumbai, India), Qualigens Fine Chemicals (Mumbai, India) and s d Fine-Chem Ltd. (Mumbai, India). All the other routine chemicals were procured from Nice Chemicals (Kochi, India). Pure distilled water was obtained with a Milli-Q system (Millipore, Tokyo, Japan).
Microorganisms and taxonomic study: Various bacterial and fungal isolates were obtained from the soil collected near a retail dairy outlet and oil cakes collected from a local oil mill in Bangalore city using serial dilution and spread plate techniques. The bacterial isolates were pure cultured, Gram’s stained and biochemically characterized using standard protocols. The sporulating fungi were identified based on the colony morphology and microscopical features. Clinical isolate of Candida albicans was obtained from the Department of Microbiology, Genohelix Biolabs, Bangalore. Pure cultures of the bacterial and fungal isolates were maintained on nutrient agar and Sabouraud dextrose agar slants respectively, at 4°C until use.
Screening for lipolytic activity: Selection of lipase producers was made by inoculating the bacterial and fungal cultures on tributyrin agar supplemented with 0.5% (v/v) Triton-X 100. The bacterial plates were incubated at 37°C for 2 days and the fungal plates were incubated at 27°C for 3-5 days. The lipolytic activity was indicated by the formation of a transparent zone of lipolysis surrounding the colonies against a turbid culture medium.
Source of lipid substrates: Different oil sources such as olive, coconut, groundnut and refined sunflower oil were procured from the local market in Bangalore city, India and used for the optimization of carbon sources for lipase production.
Inoculum preparation and production of lipase: Lipase production under submerged fermentation was studied in a medium containing g L-1: lipid substrate, 2; yeast extract, 5; NH4Cl, 35; K2HPO4, 3; KH2PO4, 1; MgCl2, 0.12; MgSO4.7H2O, 0.1 and distilled water, supplemented with 0.5% (v/v) Triton-X 100 at pH 7.0±0.2. Saline suspensions of the selected bacterial, yeast and mold cultures were aseptically added to the sterile broth at 1% (v/v). The inoculated media were incubated at 30°C for 2 days for the bacterium, 3 days for the yeast and 5 days for the mold under aerobic condition at 120 rpm in an orbital shaker.
Following incubation, the bacterial and yeast broths were centrifuged at 5000 rpm for 30 min at 4°C using a Hettich Universal 32R (Germany) and the supernatants were collected. The mold broth was filtered through normal filter paper and then through Whatman No. 1 filter paper (Whatman International Ltd., Maidstone, England) and the filtrate was collected. The clear supernatants and filtrates were subjected to lipase assay and further purification.
Lipase assay: Enzymatic assay of lipase was performed by a titrimetric method as per the protocol from Sigma-Aldrich Inc. The assay mixture consisted of 200 mM Tris HCl buffer, pH 7.7 at 37°C, olive oil substrate, 95% ethanol (nondenatured), 0.9% (w/v) thymolphthalein indicator solution (TPH Indic), 50 mM NaOH solution and lipase enzyme solution. Two clean conical flasks were taken, each containing 2.5 mL of deionized water, 1 mL of Tris HCl buffer and 3 mL of olive oil (substrate). The contents were mixed well by swirling. One milli liter of enzyme extract (cultured broth supernatant/filtrate) was added to the test flask and 1 mL deionized water was added to the blank. Both the flasks were incubated for 30 min at 37°C. Following incubation, 3 mL of 95% ethanol and 4 drops of thymolphthalein indicator was added to each of the flasks. The contents of the bottles were titrated against 50 mM NaOH. The end point was determined by the colour of the mixture changing from colourless to pale blue. Enzyme activity was calculated and expressed in Units mL-1. One Unit of lipase activity (U) was defined as μmoles of free fatty acids released due to enzyme action per hour of incubation per mL of enzyme extract under standard assay conditions.
The following formula was used to determine the lipase activity:
Units mL-1 of enzyme activity = (NaOH)*(Molarity of NaOH)*(1000)*(2)
| NaOH | = | Difference in the volume (in mL) of NaOH run down for the test and blank samples. Molarity of NaOH is 50 mM |
| 1000 | = | Conversion factor from milli-equivalent to micro-equivalent |
| 2 | = | Time conversion factor from 30 min to 1 h (unit definition) |
Optimization of nutritional supplements: The effect of carbon supplement on lipase synthesis was examined by incorporating different oils into the production media at varying concentrations (0.5, 1, 3 and 5% v/v). The effect of different organic nitrogen sources was investigated by incorporating tryptone, peptone, beef extract and yeast extract (1% w/v) into the production medium.
Optimization of physical parameters: The influence of physical factors such as pH, temperature, agitation speed and incubation time were also determined. The effect of pH was studied by adjusting the media pH from 4.2 to 9.2 with a gradual increment of one pH unit. Production of lipase was noted at 25, 30, 37 and 42°C. The role of agitation speed in submerged fermentation of microbial lipases was evaluated by incubating the broth cultures at 50, 100, 150 and 200 rpm in an orbital shaker. The effect of incubation time on microbial lipase production was determined after every 24 h for 5 days.
Scale-up studies and partial purification of lipases: Mass production of lipases was carried out using 500 mL of the optimized media in 1000 mL Erlenmeyer flasks, microbial cultures and all the optimized conditions. The crude enzyme extracts obtained after fermentation were subjected to 80% (w/v) ammonium sulphate precipitation. Precipitated enzyme proteins were resuspended in 250 mM Tris-HCl buffer (pH 7.0) and dialyzed with 10 mM Tris-HCl buffer (pH 7.0) overnight at 4°C.
Determination of molecular weights: The molecular weights of the partially purified lipases were determined by sodium dodecyl sulphate polyacrylamide gel electrophoresis (SDS-PAGE) using a Bio-Rad Mini-Protean Tetra Cell vertical electrophoresis unit. Electrophoresis was carried out on 10% polyacrylamide slab using the buffer system of Laemmli (1970). Samples were diluted twice with sample loading buffer consisting of 50 mM Tris-HCl pH 6.8, 10% (v/v) glycerol, 2% (w/v) SDS, 100 mM 2-mercaptoethanol and 0.1% (w/v) bromophenol blue. Gel proteins were stained with 0.2% (w/v) Coomassie brilliant blue solution. Apparent molecular weights of the microbial lipases were evaluated using broad range pre-stained protein marker (New England BioLabs, UK).
Statistical analysis: All the optimization studies were conducted in triplicates and the data were analyzed using single factor analysis of variance (ANOVA). All the data are graphically presented as the Mean±SD of triplicates (n = 3). ANOVA was performed using Microsoft Excel 2007. p values<0.05 were considered significant with a confidence limit of 95%.
RESULTS
Fifteen bacterial isolates were obtained from the soil near the retail dairy outlet located in Bangalore city through dilution and plating on nutrient agar. Pure culturing and Gram’s staining of the isolates revealed majority of them belonged to gram positive rods in chains with endospores and few gram negative rods in scattered appearance. On Sabouraud dextrose agar, nine fungal isolates comprising of six molds and three yeast strains were recovered from the oil cakes sampled from a local oil mill. Selection of the potent bacterial and fungal isolates was performed by observing the clear lipolytic zones on tributyrin triton X-100 agar. The best lipolytic bacterial strain was identified as Pseudomonas aeruginosa, which was isolated from the soil near the dairy outlet. Among the fungal strains, the clinical isolate of Candida albicans showed promising results for lipid hydrolysis, whereas, the mold isolated from the oil cakes and demonstrating maximum lipolytic activity was identified as Aspergillus flavus.
A comparative study was conducted on the optimization of process parameters affecting lipase production from P. aeruginosa, C. albicans and A. flavus. The effect of various nutritional supplements and physical factors were studied. Among the different lipid substrates, refined sunflower oil supported maximum lipase production from P. aeruginosa and C. albicans with an activity of 1840 U mL-1 and 2060 U mL-1, respectively. For the mold A. flavus, coconut oil yielded highest lipolytic activity of 1920 U mL-1 (Fig. 1).
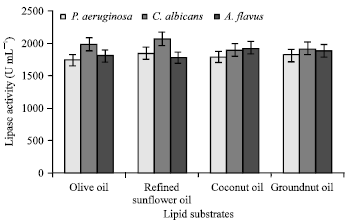 | |
| Fig. 1: | Different lipid substrates influencing extracellular lipase production from P. aeruginosa, C. albicans and A. flavus |
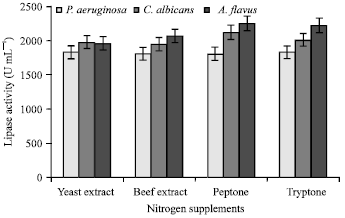 | |
| Fig. 2: | Influence of different nitrogen supplements on lipase production |
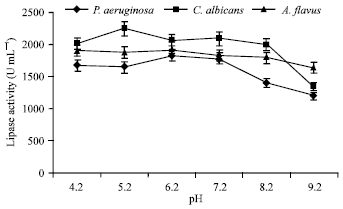 | |
| Fig. 3: | Effect of pH on lipase production |
Optimization of nitrogen supplements revealed maximum lipase production from P. aeruginosa with tryptone (1830 U mL-1). On the other hand, organic nitrogen supplement peptone resulted in optimum enzyme production from C. albicans (2110 U mL-1) and A. flavus (2240 U mL-1). The significant results have been graphically presented in Fig. 2.
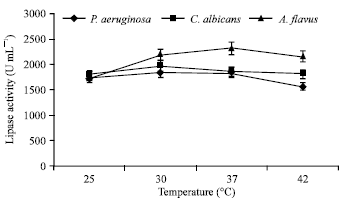 | |
| Fig. 4: | Effect of different temperatures on lipase production |
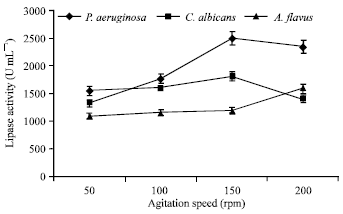 | |
| Fig. 5: | Effect of agitation speed on lipase production |
Optimization of physical parameters was also carried out. pH of the fermentation media plays a vital role in determining the level of enzyme production. pH 6.2 yielded maximum lipase production from P. aeruginosa (1840 U mL-1) and from A. flavus (1920 U mL-1). Comparatively, a lower pH of 5.2 favoured maximum lipase production from C. albicans (2250 U mL-1) (Fig. 3). A temperature of 30°C showed maximum lipase synthesis from P. aeruginosa (1830 U mL-1) and from C. albicans (1960 U mL-1), whereas 37°C proved to be most suitable for A. flavus (2310 U mL-1) (Fig. 4). In submerged fermentation, the production rate of a microbial metabolite is also influenced by the agitation speed. 150 rpm supported highest enzyme production from P. aeruginosa (2500 U mL-1) and from C. albicans (1810 U mL-1), whereas, shaker speed of 200 rpm was most favourable for A. flavus (1590 U mL-1) (Fig. 5). Lipase production has also been influenced by the time course of the fermentation process. Interestingly, the highest yield of lipase was recorded after 5 days for P. aeruginosa (2140 U mL-1), C. albicans (2330 U mL-1) and A. flavus (1940 U mL-1). The significant results have been illustrated in Fig. 6.
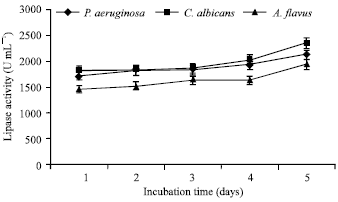 | |
| Fig. 6: | Effect of incubation time on lipase production |
The bacterial and fungal lipases were partially purified by ammonium sulphate precipitation and dialysis. The activity of the dialyzed fraction of extracellular lipase from P. aeruginosa, C. albicans and A. flavus were recorded as 400, 410 and 460 U mL-1, respectively. The dialyzed lipase fraction obtained from P. aeruginosa revealed molecular weights of 47, 49 and 51 kDa, whereas, lipases from C. albicans and A. flavus demonstrated 3 bands (16.5, 27 and 51 kDa) and one band (47 kDa), respectively.
DISCUSSION
A diverse range of microbial enzymes are finding enormous applications in food, beverage, dairy, pharmaceutical, biomedical, detergent, textile, leather, paper-pulp and cosmetic industries (Pandey et al., 1999; Saxena et al., 1999; Jaeger and Eggert, 2002; Houde et al., 2004). They act as valuable alternatives to laborious and expensive chemical methods. Lipases are among the most sought after industrial enzymes. Lipases obtained from different sources have variable activities and specificities. Some lipases exhibit affinity for short-chain fatty acids, some are specific for unsaturated fatty acids, while many others are nonspecific and randomly hydrolyse the triglycerides (Ghosh et al., 1996).
Industrial lipases are either obtained from bacteria, yeasts or molds (Hasan et al., 2010). Though a lot of reports are available on bacterial lipases, fungi are generally preferred as lipase producers because they secrete extracellular enzymes, thus facilitating the easy recovery from the fermentation broth (Jaeger et al., 1999; Pera et al., 2006). Among various bacterial members, lipase production by Pseudomonas aeruginosa has been extensively investigated (Stuer et al., 1986; Saeed et al., 2006; Syed et al., 2010). Lipase synthesis from yeasts has also been notably studied in species of Candida such as C. albicans, C. rugosa and C. antarctica (Benjamin and Pandey, 2001; Lopez et al., 2004; Vakhlu and Kour, 2006; Liu and Zhang, 2011). Among the filamentous fungi, lipase production has been documented in species of Aspergillus, Trichoderma viride, etc. (Pera et al., 2006; Kshmiri et al., 2006).
The present study deals with a comparative assessment of the various process parameters that influence extracellular lipase production from Pseudomonas aeruginosa, Candida albicans and Aspergillus flavus. A bacterium, yeast and mold have been simultaneously chosen to get a vivid understanding of their lipolytic abilities relative to each other. P. aeruginosa was isolated from the soil near the dairy outlet. The presence of this organism in that chosen environment may be correlated to the occasional spillage of milk and dairy products, thereby, adding some milk fats into the surrounding soil, favouring the proliferation of lipolytic bacterial strains. The recovery of A. flavus from the oil cakes may be attributed to the presence of fungal spores naturally occurring on oil seeds. C. albicans, the yeast form used in this study, was a clinical isolate. All these test organisms demonstrated promising potential when screened for the lipolytic activity on tributyrin agar. In P. aeruginosa the lipolytic activity was first detected, compared to C. albicans and A. flavus, indicating it to be a prolific lipase producer. Bhumibhamon et al. (2002) had described that the clear zone of lipid hydrolysis was observed around KLB1, a Pseudomonas sp., on tributyrin agar at 30°C just after 24 h of incubation.
Optimization of media and assessment of growth conditions are vital to the successful development of a fermentation process. Formulation of a cost effective medium is required to reduce the final cost of the enzyme (Das et al., 2010). Reduction of cost, enhancement of enzyme productivity and improvement of enzyme stability will contribute to a more viable fermentation process. Submerged fermentation has been preferred over the last two decades for the production of microbial enzymes because it allows better monitoring of various process parameters such as temperature, pH, nitrogen composition, carbon, lipid and inorganic source and dissolved oxygen tension in the medium (Long, 2010). Lipases are generally produced by microbes on lipid substrates such as oils, fatty acids, glycerol or tweens in the presence of an organic nitrogen source (Gupta et al., 2004). Studies on the effect of various oils as carbon sources for lipase production by P. aeruginosa, C. albicans and A. flavus showed that C. albicans was able to utilize a wide range of oils with comparatively greater yields than the bacterial and mold isolates. The highest enzyme activity of 2060 U mL-1 was detected when refined sunflower oil was used as the substrate (Fig. 1). For P. aeruginosa also refined oil proved to be the best substrate yielding an activity of 1840 U mL-1, whereas, for A. flavus coconut oil showed the highest enzyme activity of 1920 U mL-1. In contrast, Gilbert et al. (1991) used olive oil as a sole source of carbon for lipase production by P. aeruginosa. On the other hand, Amara and Salem (2009) used castor oil for lipase production from two strains of P. aeruginosa. In another study, olive oil was found to support the highest lipase production by a Penicillium species (Freire et al., 1997). Effects of varying percentages of carbon sources were also examined. Highest enzyme activity was observed in A. flavus with 3% coconut oil as the carbon source (2060 U mL-1). For P. aeruginosa 1% refined oil and for C. albicans 3% refined oil yielded maximum lipase activity of 1870 U mL-1 and 1990 U mL-1, respectively.
Nitrogen supplement plays an important role in the synthesis of enzymes, proteins and nucleotides. For P. aeruginosa tryptone was found to be the best nitrogen source for lipase production (1830 U mL-1), whereas, peptone was the preferred nitrogen source for both C. albicans (2110 U mL-1) and A. flavus (2240 U mL-1) (Fig. 2). The results of the nitrogen source optimization is in correlation with the previous study conducted by Gupta et al. (2004) wherein, organic nitrogen sources such as peptone and yeast extract were found to be preferred by bacteria. This may be due to the presence of some vitamins and accessory growth factors available in the organic nitrogen supplements.
The initial pH of the growth medium is important for the fermentation process. Enzymes of microbial origin remain metabolically active at a favourable pH range. Any drastic alteration of the media pH may result in the loss of enzyme activity by disrupting microbial membrane transport mechanisms. It was noticed that P. aeruginosa and A. flavus produced the maximum amount of enzyme at pH 6.2 and C. albicans at pH 5.2 (Fig. 3). Present result of pH optimization for lipase synthesis from P. aeruginosa is in close proximity with the optimum pH value of 6.5 reported by Gilbert et al. (1991).
Incubation temperature is indeed another significant parameter that greatly influences enzyme production. Lipase was maximally produced at 30°C for P. aeruginosa and C. albicans with an enzyme activity of 1830 U mL-1 and 1960 U mL-1, respectively (Fig. 4). For A. flavus the optimum temperature was 37°C, with an activity of 2310 U mL-1. The psychrophilic nature of P. aeruginosa may support our observation. In the present study, the highest production of lipase at 37°C by A. flavus (2310 U mL-1) may be supported by the report of Ghosh et al. (1996) which suggests that lipases are generally produced in the temperature range between 20 and 45°C. In a similar study conducted by Pera et al. (2006), A. niger was found to produce lipase at pH 6.5 and 30-35°C.
Agitation speed in submerged fermentation has a variable effect on lipase production. Optimization studies carried out with both P. aeruginosa and C. albicans showed maximum enzyme production at 150 rpm and for A. flavus at 200 rpm (Fig. 5). Moderate agitation speed facilitates extracellular lipase production from P. aeruginosa, while greater agitation resulted in decreased lipase production (Ghosh et al., 1996).
The level of enzyme production varies with the time course of the fermentation process. The effect of incubation time was monitored on a daily basis for 5 days. All the test organisms produced maximum amount of lipase after an incubation time of 5 days (Fig. 6). A maximum enzyme activity of 1934 U g-1 was obtained from Aspergillus species using olive oil when incubated for 4 days (Adinarayana et al., 2003). In the present study A. flavus demonstrated maximum lipase activity of 1940 U mL-1 when incubated for 5 days.
Following partial purification, the dialyzed lipase fraction from P. aeruginosa, C. albicans and A. flavus revealed an activity of 400, 410 and 460 U mL-1, respectively. Lipase produced previously from a strain of P. aeruginosa at 37°C showed maximum activity of 470.82 Units mL-1 (Amara and Salem, 2009) which is in close accordance with our results.
The dialyzed enzymes were analyzed using SDS PAGE. Three clear protein bands were observed for the extracellular lipase obtained from P. aeruginosa with molecular weights of 47, 49 and 51 kDa. Three clear bands were also detected for the lipase obtained from C. albicans revealing molecular weights of 16.5, 27 and 51 kDa. The dialyzed enzyme obtained from A. flavus revealed a single band having a molecular weight of 47 kDa. Previously reported extracellular lipase from P. aeruginosa exhibited molecular weight of 54.97 kDa (Saeed et al., 2006). Another lipase from P. aeruginosa BN-1 had a molecular weight of 60 kDa (Syed et al., 2010). Majority of yeast lipases are extracellular, monomeric glycoproteins with molecular weight ranging between 33 and 65 kDa, but there are reports suggesting the presence of multiple isoenzymes among yeasts like Candida albicans and C. rugosa (Vakhlu and Kour, 2006). In our study, the observation of three protein bands in lipase from C. albicans may be attributed to the existence of lipase isoforms.
ACKNOWLEDGMENTS
We wish to extend our sincere gratitude to Dr. Chenraj Jain, Chairman, Jain Group of Institutions, Bangalore, for providing us with the financial and laboratory facilities required for this research work. We also wish to thank the entire supporting staff of the laboratory whose help has been invaluable for the successful completion of our research work.
REFERENCES
- Adinarayana, K., P. Ellaiah and D.S. Prasad, 2003. Purification and partial characterization of thermostable serine alkaline protease from a newly isolated Bacillus subtillis PE-11. Pharm. Sci. Technol., 4: E56-E64.
Direct Link - Amara, A.A. and S.R. Salem, 2009. Degradation of castor oil and lipase production by Pseudomonas aeruginosa. Am. Eurasian J. Agric. Environ. Sci., 5: 556-563.
Direct Link - Anbu, P, M.J. Noh, D.H. Kim, J.S. Seo, B.K. Hur and K.H. Min, 2011. Screening and optimization of extracellular lipases by Acinetobacter species isolated from oil-contaminated soil in South Korea. Afr. J. Biotechnol., 10: 4147-4156.
Direct Link - Aravindan, R., P. Anbumathi and T. Viruthagiri, 2007. Lipase application in food industry. Indian J. Biotechnol., 6: 141-158.
Direct Link - Benjamin, S. and A. Pandey, 2001. Isolation and characterization of three distinct forms of lipases from Candida rugosa produced in solid state fermentation. Braz. Arch. Biol. Technol., 44: 213-221.
CrossRefDirect Link - Bhumibhamon, O., K. Achara and F. Suptawee, 2002. Biotreatment of high fat and oil wastewater by lipase producing microorganisms. Kasetsart J. Nat. Sci., 36: 261-267.
Direct Link - Das, A., S. Bhattacharya and L. Murali, 2010. Production of cellulase from a thermophilic Bacillus sp. isolated from cow dung. Am. Eurasian J. Agric. Environ. Sci., 8: 685-691.
Direct Link - Fadilo-Lu, S. and O. Erkmen, 2002. Effects of carbon and nitrogen sources on lipase production by Candida rugosa. Turk. J. Eng. Environ. Sci., 26: 249-254.
Direct Link - Freire, D.M., E.M.F. Teles, E.P.S. Bon and G.L. Sant' Anna Jr., 1997. Lipase production by Penicillium restrictum in a bench-scale fermenter: Effect of carbon and nitrogen nutrition, agitation and aeration. Applied Biochem. Biotechnol., 63: 409-421.
CrossRefPubMedDirect Link - Fu, X., X. Zhu, K. Gao and J. Duan, 1995. Oil and fat hydrolysis with lipase from Aspergillus sp. J. Am. Oil Chem. Soc., 72: 527-531.
CrossRefDirect Link - Ghosh, P.K., R.K. Saxena, R. Gupta, R.P. Yadav and S. Davidson, 1996. Microbial lipases: Production and applications. Sci. Prog., 79: 119-157.
PubMedDirect Link - Gilbert, E.J., A. Cornish and C.W. Jones, 1991. Purification and properties of extrcellular lipase from Pseudomonas aeruginosa EF2. J. Gen. Microbiol., 137: 2223-2229.
Direct Link - Gitlesen, T., M. Bauer and P. Adlercreutz, 1997. Adsorption of Lipase on polypropylene powder. Biochim. Biophys. Acta, 1345: 188-196.
CrossRefDirect Link - Gulati, R., J. Isar, V. Kumar, A.K. Prasad, V.S. Parmar and R.K. Saxena, 2005. Production of a novel alkaline lipase by Fusarium globulosum using neem oil and its applications. Pure Applied Chem., 77: 251-262.
CrossRefDirect Link - Gupta, R., N. Gupta and P. Rathi, 2004. Bacterial lipases: An overview of production, purification and biochemical properties. Applied Microbiol. Biotechnol., 64: 763-781.
CrossRefDirect Link - Hasan, F., A.A. Shah, S. Javed and A. Hameed, 2010. Enzymes used in detergents: Lipases. Afr. J. Biotechnol., 9: 4836-4844.
Direct Link - Houde, A., A. Kademi and D. Leblanc, 2004. Lipase and their industrial applications: An overview. Appl. Biochem. Biotechnol., 118: 155-170.
PubMed - Jaeger, K.E.H., S. Ransac, B.W. Dijkstr, C. Colson, M. van Heuvel and O. Misset, 1994. Bacterial lipases. FEMS Microbiol. Rev., 15: 29-63.
Direct Link - Jaeger, K.E., B.W. Dijkstra and M.T. Reetz, 1999. Bacterial biocatalysts: Molecular biology, three-dimensional structures and biotechnological applications of lipases. Annu. Rev. Microbiol., 53: 315-351.
CrossRef - Jaeger, K.E. and T. Eggert, 2002. Lipases for biotechnology. Curr. Opin. Biotechnol., 13: 390-397.
CrossRefPubMedDirect Link - Kshmiri, M.A., A. Adnan and B.W. Butt, 2006. Production, purification and partial characterization of lipase from Trichoderma viride. Afr. J. Biotechnol., 5: 878-882.
Direct Link - Liu, J. and Y. Zhang, 2011. Optimisation of lipase production by a mutant of Candida antarctica DSM-3855 using response surface methodology. Int. J. Food Sci. Tech., 46: 695-701.
CrossRef - Lopez, N., M.A. Pernas, L.M. Pastrana, A. Sanchez, F. Valero and M.L. Rua, 2004. Reactivity of pure Candida rugosa lipase isoenzymes (Lip1, Lip2, and Lip3) in aqueous and organic media. Influence of the isoenzymatic profile on the lipase performance in organic media. Biotechnol. Prog., 20: 65-73.
CrossRef - Mateos Diaz, J.C., J.A. Rodriguez, S. Roussos, J. Cordova, A. Abousalham, F. Carriere and J. Baratti, 2006. Lipase from the thermotolerant fungus Rhizopus homothallicus is more thermostable when produced using solid-state fermentation than liquid fermentation procedures. Enzyme Microb. Technol., 39: 1042-1050.
CrossRef - Nesbit, E.A. and M. Gunasekaran, 1993. Influence of cultural conditions on growth and lipolytic activity in Nocardia asteriodes. Folia Microbiol., 38: 451-455.
CrossRef - Pandey, A., S. Benjamin, C.R. Soccol, P. Nigam, N. Krieger and V.T. Soccol, 1999. The realm of microbial lipases in biotechnology. Biotechnol. Applied Biochem., 29: 119-131.
PubMedDirect Link - Pera, L.M., C.M. Romero, M.D. Baigori and G.R. Castro, 2006. Catalytic properties of lipase extracts from Aspergillus niger. Food Technol. Biotechnol., 44: 247-252.
Direct Link - Rajendran, A. and V. Thangavelu, 2007. Sequential optimization of culture medium composition for extracellular lipase production by Bacillus sphaericus using statistical methods. J. Chem. Technol. Biotechnol., 82: 460-470.
CrossRef - Rani, C. and A. Panneerselvam, 2009. Influence of environmental and nutritional parameters on lipase production. ARPN J. Agric. Biol. Sci., 4: 39-43.
Direct Link - Saeed, H.M., I.T. Zaghloul, I.A. Khalil and T.M. Abdelbaeth, 2006. Molecular cloning and expression in Escherichia coli of Pseudomonas aeruginosa lipase gene. Biotechnology, 5: 62-68.
CrossRefDirect Link - Saxena, R.K., P.K. Ghosh, R. Gupta, W.S. Davidson, S. Bradoo and R. Gulati, 1999. Microbial lipases: Potential biocatalysts for the future industry. Curr. Sci., 77: 101-115.
Direct Link - Sharma, R., Y. Chisti and U.C. Banerjee, 2001. Production, purification, characterization and applications of lipases. Biotechnol. Adv., 19: 627-662.
CrossRef - Stuer, W., K.E. Jaeger and U.K. Winkler, 1986. Purification of extracellular lipase from Pseudomonas aeruginosa. J. Bacteriol., 168: 1070-1074.
Direct Link - Syed, M.N., S. Iqbal, S. Bano, A.B. Khan, S. Ali-ul-Qader and A. Azhar, 2010. Purification and characterization of 60 kD lipase linked with chaperonin from Pseudomonas aeruginosa BN-1. Afr. J. Biotechnol., 9: 7724-7732.
Direct Link - Vakhlu, J. and A. Kour, 2006. Yeast lipases: Enzyme purification, biochemical properties and gene cloning. Electron. J. Biotechnol., 9: 69-85.
CrossRefDirect Link