
Serife Evrim Arici
Department of Plant Protection, Faculty of Agriculture, University of Suleyman Demirel, Isparta, Turkey
Namik Kemal Koc
Department of Plant Protection, Faculty of Agriculture, University of Cukurova, Adana, Turkey
Pakistan Journal of Biological Sciences
Year: 2010 | Volume: 13 | Issue: 3 | Page No.: 138-142







ABSTRACT
The aim of the current study was to estimate the genetic diversity of F. graminearum and F. culmorum isolated from wheat in Adana province of Turkey by investigating by RAPD-PCR polymorphisms. Fusarium sp. affecting wheat crops were surveyed for 2 years during growing seasons in Adana, Turkey. Thirty two isolates of Fusarium were isolated from seeds and basal stem nodes of wheat displaying disease symptoms. These isolates were identified as F. avenecaum, F. culmorum, F. graminearum and F. croockwellense Fusarium graminearum was the predominant pathogen isolated, followed by F. culmorum F. avenecaum and F. croockwellense, respectively. Genetic distances of F. graminearum and F. culmorum isolates appeared more intense in wheat fields in Adana region were determined by RAPD-PCR (10 primers). One primer (OPU-17) with F. culmorum and two primers (OPU-19 and OPU-17) with F. graminearum did not react. Amplification products gave 27 polymorphic bands for F. culmorum and 9 bands for F. graminearum. The RAPD-PCR analysis showed that F. graminearum and F. culmorum isolates collected from Adana/Turkey were genetically varied.
PDF Abstract XML References Citation
How to cite this article
Serife Evrim Arici and Namik Kemal Koc, 2010. RAPD-PCR Analysis of Genetic Variation among Isolates of Fusarium graminearum and Fusarium culmorum from Wheat in Adana Turkey. Pakistan Journal of Biological Sciences, 13: 138-142.
DOI: 10.3923/pjbs.2010.138.142
URL: https://scialert.net/abstract/?doi=pjbs.2010.138.142
DOI: 10.3923/pjbs.2010.138.142
URL: https://scialert.net/abstract/?doi=pjbs.2010.138.142
INTRODUCTION
Fusarium diseases caused by various Fusarium species are among of the most economically important wheat disease in humid and semi-humid wheat growing areas worldwide (Parry and Nicholson, 1996; McMullen et al., 1997; Gilbert and Tekauz, 2000). Infection of wheat by the pathogen reduces grain yield and quality, cause kernel contamination with deoxynivalenol (DON) and other mycotoxins, which are known to be harmful to livestock and pose a safety concern in human food (Miller, 1995; Placinta et al., 1999; Bai et al., 2001; Shaner, 2003). Currently, there is no satisfactory cultural or chemical control for Fusarium diseases (McMullen et al., 1997; Shaner, 2003; Tekauz et al., 2003).
Turkey is one of the largest wheat producers in the world with 16-21 million tones of production from 9.35 Mha at an average yield of 2 t ha-1 (Braun et al., 2001). In Turkey wheat is grown with natural rainfall alone or with irrigation in cereal fallow rotation. Fusarium species contribute to significant yield losses ranging from 24-36% in cultivated winter wheat varieties commonly grown in Central Anatolian Region (Aktaş et al., 1996; Hekimhan et al., 2004). Wheat has been widely grown for many years in Adana province of Turkey. In the last few or several years Fusarium Head Blight (FHB) and Crown Rot (CR) symptoms have been observed in this region. However, to our knowledge no studies have been carried out to determine the Fusarium species that are found on wheat in this region.
The aim of the current study was to estimate the genetic diversity of F. graminearum and F. culmorum isolated from wheat in Adana province of Turkey by using by (RAPD-PCR) polymorphisms and to assess the usefulness of these characters in identifying F. graminearum and F. culmorum isolates.
MATERIALS AND METHODS
Sampling: Samples of wheat plants infected with Fusarium were collected from stem and ears exhibiting symptoms of Fusarium sp., in Adana province. The study was undertaken over the period April-May 2002-2003. Symptomatic plants were collected within each field. In order to isolate Fusarium sp. from infected plants, both tissue and grains of infected wheat were surface-sterilized. Intact subcrown internodes were excised from the top of the plants and washed thoroughly in tap water. Dead basal leaf-sheaths were removed from the intact subcrown internodes and cut into 3-5 mm length sections. The tissue sections and the grains were surface sterilized by immersion in 1% sodium hypochlorite for 1 min and, for 10 min, respectively. They were rinsed throughly in sterile distilled water, dried on sterile filter paper and placed onto Potato Dextrose Agar (PDA) supplemented with 100 mg L-1 streptomycin sulfate in 9 cm petri dishes. The dishes were incubated for 4-8 d at 25°C under conditions of alternating light/dark photoperiod (16 h light/8 h dark). Uncontaminated mycelium was transferred to fresh PDA medium for identification. Single-spore isolation was performed for each sample. Cultures were identified based on the morphology of macroconidia, microconidia, chlamydospores, conidiophores, general colony morphology and taxonomic descriptions as suggested by Nelson et al. (1983) and Nicholson et al. (1998). Single-spore cultures of selected isolates were transferred to PDA medium in tubes for preservation.
DNA extraction: F. graminearum and F. culmorum isolates appeared more intense in wheat fields in Adana region were cultured at 26°C on Potato Dextrose Agar (PDA). Mycelium was scraped from the surface of 7 days old colonies on PDA, inoculated into 100 mL Erlenmeyer-flasks containing 20 mL liquid media (40 g glucose, 5 g peptone, 3 g yeast extract, 3 g malt extract and water up to 1 L) and incubated at 26°C on rotary shaker (150 rpm) for 5 days. Mycelium of F. graminearum and F. culmorum were harvested by filtration from liquid cultures and ground to fine powder in liquid nitrogen, DNA was extracted and purified for molecular analysis according to the methods of Peever et al. (1999). Mycelium of each isolate was extracted with lysis buffer containing 50 mM EDTA, 100 mM Tris-(TE), pH:8.0 and 3% sodium dodecyl sulfate (SDS) for 30-45 min at 65°C. Mycelium was pelleted by centrifugation at 14.000 rpm for 15 min and the supernatant was precipitated with 8 M potassium acetate at -20°C for 15 min. After spinnin the supernatant was subjected to two rounds of phenol:chloroform:isoamyl alcohol (25:24:1) extraction and chloroform:isoamyl alcohol (24:1 v/v) extraction. The final supernatant was precipitated with 2 volumes of 100% ethanol and 0.5 M NaCl. Pellets were resuspended in water, precipitated by 14% polyethylene glycol and 1 M NaCl and finally resuspended in 50 μL of TE buffer (10 mM, 1 mM EDTA). RNA was digested with RNAse A (Sigma Chemical Co., St. Lois) at 20 μL and 37°C for 3 h. Extracted DNA samples was stored at -20°C until required.
RAPD-PCR Analysis: The total reaction volume was 25 μL and the conditions were 1.5 mM MgCl2; 100 μM each dNTP; 200 nM oligonucleotide primers (for RAPD-PCR reactions), 1 u of Taq DNA polymerase, 10-20 ng template DNA, 2.5 μL PCR buffer (10 mM Tris-HCl, pH:8.3, 50 mM KCl). The primers used for RAPD-PCR were from Operon Technologies Inc. (Alameda CA). Ten primers OPT-01, OPT-04, OPT-06, OPT-16, OPT-18, OPU-13, OPU-15, OPU-19, OPU-17 and UBC-85 were selected for a preliminary screen of several kits. Amplifications were performed in a DNA Thermocycler (Techne, Genius). For RAPD-PCR amplifications, the thermocycler was programmed for one cycle at 94°C for 1 min, followed by 45 cycles of denaturation at 94°C for 1 min, annealing at 36°C for 1 min and extension at 72°C for 2 min. The final cycle had 5 min extension step at 72°C. The PCR fragments were analyzed through 1.5% agarose gel electrophoresis, stained with ethidium bromide (0.5 g mL-1) and visualized under ultraviolet light.
Data analysis: Data were scored as 1 for the presence and 0 for the absence of a DNA bands for each isolates. The data matrix was entered into the MVSP (MultiVariete Statistical Package) program.
RESULTS
Four species of Fusarium were isolated from wheat in Adana province. A total of 35 Fusarium isolates were obtained and identified as F. avenaceum, F. culmorum, F. graminearum and F. crookwellense (Fig. 1a, b-4a, b). From these isolates, 43% were identified as F. graminearum, 29% as F. culmorum, 17% as F. crookwellense and 11% as F. avenaceum.
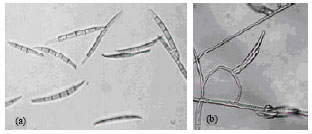 | |
| Fig. 1: | (a) Macroconidia and (b) conidiophores of Fusarium avenaceum (X400) |
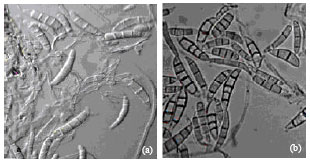 | |
| Fig. 2: | (a-b) Macroconidia of Fusarium culmorum (X400) |
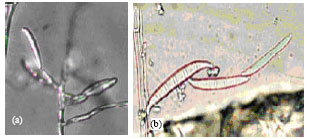 | |
| Fig. 3: | (a) Conidiophores and (b) macroconidia of Fusarium graminearum (X400) |
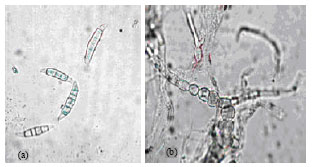 | |
| Fig. 4: | (a) Macroconidia and (b) chlamydospores of Fusarium crookwellense (X400) |
No sexual stage has been identified for F. culmorum. The present investigation showed that F. avenaceum, F. culmorum, F. graminearum and F. croockwellense caused diseases on wheat and F. graminearum was the most widely distributed species on winter wheat in Adana province. (data not shown).
RAPD-PCR analysis was performed on 10 isolates of F. graminearum and F. culmorum appeared more intense in wheat fields in Adana region were determined by RAPD-PCR using 10 different random primers. The genetic variability of Adana province F. graminearum and F. culmorum isolates was investigated. One primer (OPU-17) with F. culmorum and two primers (OPU-19 and OPU-17) with F. graminearum did not react. Amplification products gave 27 polymorphic bands for F. culmorum and 9 bands for F. graminearum. Fusarium culmorum isolates were highly varied with most primers, whereas F. graminearum was not. Fusarium graminearum isolates, all obtained from wheat in Adana province, yielded a 0.4 kb product, whereas F. culmorum isolates from wheat yielded a 0.3 kb product (Fig. 5, 6). The data obtained for ten isolates and 10 primers were pooled and a total of 134 bands were scored. Primer OPT.8 amplified a fragment of approximately 0.8 kb from DNA of all isolates of F.culmorum and primer OPT-15 amplified a fragment of approximately 0.7 kb from DNA of all isolates of F. graminearum.
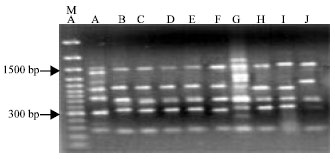 | |
| Fig. 5: | Agarose gel showing random amplified polymoric DNA (RAPD-PCR) profiles of F. culmorum amplified with arbitrary primer OPT-01 (MA: 100 bp DNA ladder, A: B-1, B: B-2, C: B-3, D: B-4, E: B-5, F: B-8, G: B-9, H: B-10, I: B-11, J: B-12 isolates) |
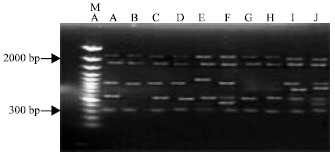 | |
| Fig. 6: | Agarose gel showing random amplified polymoric DNA (RAPD-PCR) profiles of F.gramenarum amplified with arbitrary primer OPT-06 (MA: 100 bp DNA ladder, A: A-15, B: A-8, C: A-6, D: A-2, E: A-3, F: A-4, G: A-11, H: A-13, I: A-16, J: A-9 isolates) |
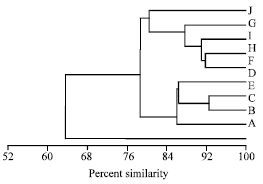 | |
| Fig. 7: | Dendrogram showing genetic relationships of Fusarium culmorum isolates studied. The data matrix was entered into the MVSP (MultiVariete Statistical Package) program |
The MVSP analysis by genetic distance failed to identify spatial clustering among the Adana region different geographic regions (Fig. 7). F (B-8), G (B-9), H (B-10), I (B-11), J (B-12) isolates of F. culmorum were placed in one group. The isolates of F. culmorum could be clustered in 3 groups.
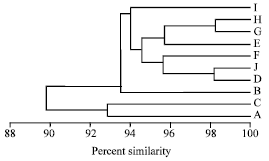 | |
| Fig. 8: | Dendrogram showing genetic relationships of F. graminearum isolates studied. The data matrix was entered into the MVSP (MultiVariete Statistical Package) program |
The most genetic similarity was observed between isolates F (B-8)/G (B-9). Isolates A (B-1) appeared to be the most genetically distinct.
B (A-8), D (A-2), E (A-3), F (A-4), J (A-9) and I (A-16) isolates of F. graminearum were placed in one group. Isolates of I (A-16) and J (A-9) were the most genetically similar. The isolates of F. graminearum could be clustered in 2 groups. Isolates A (A-15) and C (A-6) constituted a different group and were the most genetically distinct (Fig. 8). As a result, the RAPD-PCR analysis revealed that F. graminearum and F. culmorum isolates collection from Adana/Turkey were genetically varied.
DISCUSSION
In this study the fungal isolates were identified a combination of morphological and molecular marker techniques. Fusarium avenaceum, F. culmorum, F. graminearum and F. crookwellense have been reported to cause foot rot and head blight on winter wheat in Europe (Parry and Nicholson, 1996; Ahmad Khan et al., 2005). The present investigation showed that F. avenaceum, F. culmorum, F. graminearum and F. crookwellense caused diseases on wheat and F. graminearum was the most widely distributed species on Winter wheat in Adana province. The results clearly support the results of other European surveys indicating the increasing importance of F. graminearum as a major wheat pathogen. F. graminerum, F. moniliforme, F. culmorum, F. sporodotricoides, F. triticana, F. heterosporium and F. avenaceum had previously been reported on wheat plants in Turkey (Aktaş et al., 1996, 1999). The present study is the report of Fusarium species of wheat in Adana, Turkey.
The objective of this study was to estimate the extent of genetic diversity of F. graminearum and F. culmorum appeared more intense in wheat fields in Adana region, Turkey by means of RAPD-PCR markers. It was successfully applied for differentiation of F. graminearum and F. culmorum. One primer (OPU-17) with F. culmorum and two primers (OPU-19 and OPU-17) with F. graminearum did not react. Amplification products have given 27 polymorphic bands for F. culmorum and 9 bands for F. graminearum. RAPD-PCR analysis revealed a high level of genetic variability in populations of F. graminearum and F. culmorum. The RAPD-PCR analysis suggests that the Adana/Turkey F.graminearum and F. culmorum isolate collection was genetically varied. Some researchers have reported high levels of genotypic variation using RFLP analysis of F. culmorum and F. avenecaum strains in England (Mieadaner and Schilling, 1996). On the other hand, little variation was observed among isolates of F. culmorum based on RAPD-PCR analysis (De Nijs et al., 1997). The fungal isolates identified using a combination of morphological and DNA marker techniques. Each isolate had to show the correct morphology on PDA and yield 0.8 kb from DNA of all isolates of F. culmorum and a fragment of approximately 0.3 kb from DNA of all isolates of F. graminearum in the RAPD-PCR analysis. RAPD-PCR assays are more convenient, because there is no need for radioactive probes. Results obtained so far indicate that it will be continued to generate molecular markers necessary to DNA fingerprint F. graminearum and F. culmorum. Isolates of F. graminearum and F. culmorum from wheat kernels and stem were analyzed. RAPD-PCR analysis revealed a high level of genetic variability in populations of F. culmorum. Other researchers have reported high levels of genetic diversity in local populations of F. culmorum. Schilling et al. (1996) found a high degree of genetic variability for aggressiveness within single field populations of F. culmorum and F. graminearum. De Nijs et al. (1997) reported that RAPD-PCR analysis of 17 F. culmorum isolates revealed that most of the genetic diversity was within population while genetic diversity between populations represented a small proportion of the total.
In conclusion F. graminearum and F. culmorum isolates collection from Adana/Turkey was genetically varied. The ability of RAPD-PCR analysis for isolates of F. graminearum and F. culmorum provided a tool for further investigations into these pathogens. Knowledge of Fusarium population diversity in Turkey is essential for developing effective disease management strategies. This information will be useful for ecological and epidemiological studies, especially in the development of resistant cultivars through improved screening procedures and will also optimize chemical and biological controls.
ACKNOWLEDGMENTS
The authors are thankful to Prof. Dr. Berna Tunali for the identification of Fusarium isolates and Çukurova University Research Fund for partial financial support of this research.
REFERENCES
- Bai, G.H., R. Plattner, A. Desjardins and F.L. Kolb, 2001. Resistance to Fusarium head blight and deoxynivalenol accumulation in wheat. Plant Breed., 120: 1-6.
Direct Link - De Nijs, M., J.S. Larsen, W. Gams, F.M. Rombouts, K. Wernars, U. Thrane and S.H. Notermans, 1997. Variations in random amplified polymorphic DNA patterns and secondary metabolite profiles with in Fusarium species from cereals from various parts of The Netherlands. J. Food Microbiol., 14: 449-457.
CrossRefDirect Link - Gilbert, J. and A. Tekauz, 2000. Review: Recent developments in research on Fusarium head blight of wheat in Canada. Can. J. Plant Pathol., 22: 1-8.
Direct Link - Ahmad Khan, I., F.S. Awan, A. Ahmad, Y. Fu and A. Iqbal, 2005. Genetic diversity of Pakistan wheat germplasm as revealed by RAPD markers. Genet. Resour. Crop Evol., 52: 239-244.
Direct Link - McMullen, M.P., R. Jones and D. Gallenberg, 1997. Scab of wheat and barley: A re-emerging disease of devastating impact. Plant Dis., 81: 1340-1348.
CrossRefDirect Link - Mieadaner, T. and A.G. Schilling, 1996. Genetic variation of Aggressivenesin individual field populations of Fusarium graminearum and Fusarium culmorum tested on young plants of winter rye. Eur. J. Plant Pathol., 102: 823-830.
Direct Link - Miller, J.D., 1995. Fungi and mycotoxins in grain: Implications for stored product research. J. Stored Prod. Res., 31: 1-16.
CrossRefDirect Link - Nelson, P.E., T.A. Toussoun and W.F.O. Marasas, 1983. Fusarium Species: An Illustrated Manual for Identification. 1st Edn., Pennsylvania State University Press, University Park, University Park, PA., USA., ISBN-13: 978-0271003498, Pages: 226.
Direct Link - Nicholson, P., D.R. Simpson, G. Weston, H.N. Rezanoor, A.K. Lees, D.W. Parry and D. Joyce, 1998. Detection and quantification of Fusarium culmorum and Fusarium graminearum in cereals using PCR assays. Physiol. Mol. Plant Pathol., 53: 17-37.
CrossRefDirect Link - Parry, D.W. and P. Nicholson, 1996. Development of a PCR assay to detect Fusarium poae in wheat. Plant Pathol., 45: 383-391.
CrossRefDirect Link - Peever, T.L., Y. Canihos, L. Olsen, A. Ibanez, Y.C. Liu and L.W. Timmer, 1999. Population genetic structure and host specificity of Alternaria sp. causing brown spot of Minneola tangelo and rough lemon in Florida. Phytopathology, 89: 851-860.
Direct Link - Placinta, C.M., J.P.F. D'Mello and A.M.C. Macdeoxynivalenolald, 1999. A review of worldwide contamination of cereal grains and animal feed with Fusarium mycotoxins. Anim. Feed Sci. Technol., 78: 21-37.
CrossRefDirect Link - Schilling, A.G., E.M. Moller and H.H. Geiger, 1996. Polymerase chain reaction-based assays for species-specific detection of Fusarium culmorum, F. graminearum and F. avenaceum. Phytopathology, 86: 515-522.
Direct Link - Shaner, G., 2003. Epidemiology of Fusarium Head Blight of Small Grain Cereals in North America. In: Fusarium Head Blight of Wheat and Barley, Leonard, K.J. and W.R. Bushnell (Eds.)., American Phytopathological Society Press, St. Paul, MN, pp: 84-119.
Direct Link