
A. Sarkaki
Physiology Research Center, Department of Physiology, Faculty of Medicine
M. Badavi
Physiology Research Center, Department of Physiology, Faculty of Medicine
H. Aligholi
Physiology Research Center, Department of Physiology, Faculty of Medicine
A. Zand Moghaddam
Department of Nutrition, Faculty of Paramedical, Ahvaz Jondishpour University of Medical Sciences, Ahvaz, Iran
Pakistan Journal of Biological Sciences
Year: 2009 | Volume: 12 | Issue: 20 | Page No.: 1338-1345







ABSTRACT
The aim of the present study was to investigate the preventive effect of 4 weeks soy meal (±isoflavone) on post-menopausal cognitive deficiency and body weight alteration in ovariectomized (OVX)-6-hydroxy dopamine (6-OHDA)-induced animal model of Parkinson’s Disease (PD) which mimics status in menopause women. Female Wistar rats (250-300 g, 5-6 months old) were divided into 2 main groups. (1) Control; (2) OVX; included 5 subgroups that were pre-treated with 10 or 20 g soy with isoflavone in 30 g daily diet (10 and 20 groups, respectively), 10 or 20 g soy without isoflavone in 30 g daily diet (-10 and -20 groups, respectively) and 0 g soy (sham treated group) during 4 weeks after OVX. To induce animal model of PD in main second group (OVX rats) the substantia nigra pars compacta (SNpc) was lesioned by 6-hydroxydopamine (6-OHDA) (8 μg kg-1 4 μL-1 normal saline contains 0.1% ascorbate). All animals were trained in Morris water maze for evaluating the spatial learning and memory. The results indicated that pre-treatment of Parkinsonian rats with different doses of dietary soy meal (±isoflavone) improved the spatial learning and memory and prevents increasing the body weight after menopause significantly. Our data show that, long-duration dietary soy meal may have the potential neuroprotective effect against post-menopausal cognitive deficiency induced by degeneration of nigrostriatal dopaminergic system and constant body weight during post-menopausal life cycle.
PDF Abstract XML References Citation
How to cite this article
A. Sarkaki, M. Badavi, H. Aligholi and A. Zand Moghaddam, 2009. Preventive Effects of Soy Meal (±Isoflavone) on Spatial Cognitive Deficiency
and Body Weight in an Ovariectomized Animal Model of Parkinson's Disease. Pakistan Journal of Biological Sciences, 12: 1338-1345.
DOI: 10.3923/pjbs.2009.1338.1345
URL: https://scialert.net/abstract/?doi=pjbs.2009.1338.1345
DOI: 10.3923/pjbs.2009.1338.1345
URL: https://scialert.net/abstract/?doi=pjbs.2009.1338.1345
INTRODUCTION
Parkinson's Disease (PD) is one of the most common neurodegenerative disorders (Shulman, 2002) and is a movement disorder characterized by a progressive loss of nigrostriatal dopaminergic neurons, which results in a dramatic decrease in dopamine content in the corpus striatum (Baluchnejadmojarad et al., 2009; Bureau et al., 2008; Kyuhou, 2008; Occhiuto et al., 2009). The cause of this dopaminergic deficiency is unknown, Genetic and environmental factors may contribute to the pathogenesis and progression of PD. Some clinical studies suggest that PD symptoms may be exacerbated after menopause and delayed or alleviated with hormone replacement therapy. It is defined by the triad of symptoms comprising bradykinesia or akinesia, rigidity and tremor (Gerlach and Riederer, 1996). Although, PD is noted for its motor abnormalities, there is now clear evidence that PD patients have memory deficits (Gabrieli, 1996). Several studies indicate that Preliminary Clinical reports suggest a potential protective effect of estrogen replacement therapy (ERT) on the onset and progression of both PD and PD-related dementia and increase performance on some tests of memory/cognition (El-Bakri et al., 2004; Green and Simpkins, 2000; Heikkinen et al., 2004; Markham et al., 2002). However, estrogen has proliferative and oncogenic effects on non-neural cells which are responsive to estrogen, such as breast and endometrium cells (Bang et al., 2004). For this reason, other estrogenic agents with fewer side effects are needed to develop alternative treatment strategies. For the CNS, the ideal estrogen-like compound would have activity in the brain and none in the periphery (Cyr et al., 2002). Phytoestrogens, plant-derived nonestroidal estrogens found in high abundance in most soy food products, may contain beneficial effects of estrogen without its unwanted side-effects (Lephart et al., 2002). A large body of experimental evidence supports a role for oxidative stress as a mediator of nerve cell death in PD (Gelinas and Martinoli, 2002). Emerging evidence suggests beneficial effects of estrogen and estrogen-like chemicals on neurodegenerative diseases, especially PD. Estrogen and phytoestrogens such as the isoflavones have received considerable attention in Parkinson's disease (PD) research. It has been reported to possess neuroprotective effects on dopaminergic neurons in the substantia nigra pars compacta (SNpc). Soy and isoflavones appear particularly promising for post-menopausal women at risk for PD. Genistein, an isoflavone naturally found in soy products, displays estrogenic properties and has been reported to prevent neuronal degeneration caused by increased oxidative burden. However, little is known about the influence of phytoestrogens on the brain or behavior. By this study, we aim to provide more evidence for the involvement of phytoestrogens found in soy in the process of neuroprotection and to test our hypothesis that some of these compounds may act as neuroprotective molecules and have a lesser hormonal effect than estrogens. So, aim of the present study was to elucidate the preventive effects of 4 weeks soy meal (±isoflavone) on dementia (spatial learning and memory) and body weight in ovariectomized (OVX), 6-hydroxy dopamine (6-OHDA)-induced PD model rats which mimics post-menopausal status in women.
MATERIALS AND METHODS
Subjects: Forty two female Wistar rats (250-300 g, 5-6 months) were purchased from Ahvaz Jondishapour University of Medical Sciences (AJUMS) animal house (from January 13, 2008 to June 25, 2009). Rats were individually housed in standard cages, under a 12 h light/dark cycle (light on at 7:00 am) and 55-60% humidity. Animals were allowed free access to water and food ad libtium. Experiments approved by the Jondishapour academy of sciences ethics local committee Ahvaz, Iran. All rats were gentle handled for 5 days (5 min daily) to familiar them, the animal care and experimental protocols done prior to the onset of behavioral tests.
Animals were divided randomly into 2 main groups: intact/control (C) and overiectomized (OVX). After one week adaptation and handling, ovaries of rats in second main group (OVX), were removed surgically (overiectomized) while under anesthesia induced by injection of ketamine HCl (60 mg kg-1 i.p., Rotex Medica. Trittau. Germany) and Xylazine (10 mg kg-1 i.p., Miles laboratories, Shawnee). All efforts were made to minimize the number of animals used. OVX animals were divided randomly into five subgroups (8 in each). They pre-treated by one of five types of prepared diets during four weeks prior SNpc lesioning by 6-OHDA; normal diet (0), 10 g soy with isoflavone (10), 20 g soy with isoflavone (20), 10 g soy without isoflavone (-10) and 20 g soy without isoflavone (-20) in 30 g daily diet.
Diet preparation: In order to prepare isoflavone free soy, ethanol (80 %) was added to soy powder. After 24 h, the soy was dried under Laboratory room temperature. Before and after alcohol evaporating, total isoflavone concentration of soy was determined by high performance liquid chromatography (HPLC).
Stereotaxic surgery: Four weeks after OVX, In order to create animal model of postmenopausal with Parkinson's disease, the left substantia nigra pars compacta (SNpc) of animals, was lesioned by injection of 6-OHDA (8 μg/4 μL in normal saline contain 0.1% ascorbate, Sigma, USA) under anesthesia induced by ketamine HCl (60 mg kg-1, i.p.,) and Xylazine (10 mg kg-1, i.p.,) and stereotaxic coordinations of anterior-posterior (AP); -4.8 mm to bregma, medial-lateral (Ml); 1.6, dorsal-ventral (DV); -8.2 (Paxinos and Watson, 2008).
Morris water maze
Training apparatus: In the maze training, taken place during the light phase of the cycle between 8:00 am and 5:00 pm. A circular pool was used as described by Morris with some modification (Widy-Tyszkiewecz et al., 2002). It was a black circular pool (120 cm in diameter and 80 cm in height) filled with tap water (27±21°C) with a depth 60 cm. The maze divided geographically into four equal size quadrants and release points designed in each quadrant as North (N), East (E), South (S) and West (W). A hidden circular escape platform (12 cm in diameter), was emerged 2 cm below the water level and was located in the center of the northeast quadrant. Some fixed visual cues including computer, desk, shelves, posters and illumination lights placed on the walls around the pool. A camera positioned above the center of the pool that connected to a computer to record the animal motions. An automated tracking system (Radiab ver. 2, Tehran, Iran) used to measure the escape latency, swimming distance and speed.
Acquisition trials: Spatial training in Morris water maze was began after one week recovery following SNpc lesioning. In this task, the rats were trained to find a submerged platform using extra maze cues. Prior to water maze testing, all rats were habituated to the water using a three-trial shaping procedure. This procedure habituated the rats to the water and taught them to escape from the water by climbing on to a platform. Subjects were trained across one day.
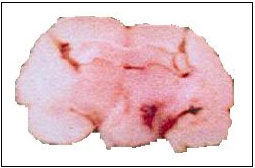 | |
| Fig. 1: | Coronal section of substantia nigra pars compacta (SNpc) which histologically stained with cresyl violet to verifying the site of lesion for making the animal model of Parkinson's disease |
Each rat received 18 trials per day. There was a 20 min break between each 3 trials (6 blocks, each block consist of 3 trials). The location of submerged platform did not change throughout the experiment. For each trial, the subject was released in water facing the edge of the tank from random start points. On each trial, the subject was allowed 60 sec to escape to the submerged platform; rats that failed to escape were led to the platform gently by experimenter and were allowed to remain on it for 30 sec before being removed from the maze and dried off (Norris and Foster, 1999).
Probe trial: One day after acquisition, only one probe trial was order. It was identical to the acquisition trials with one exception. During the probe trial, the submerged platform was removed. The percent time that each subject spent into the target quadrant while platform was removed during probe trial.
Body weight and plasma estrogen: Animal's body weight at the baseline and four weeks after soy treatment was recorded. Serum estrogen level was measured by ELISA test (USCN Life Science and Technology Co., USA).
Histology: At the end of the experiments, rats were deeply and irreversible re-anesthetized (Ketamine HCl, 100 mg kg-1) and sacrificed. To verify the location of lesion site the crystal violet (0.5 μL) injected into SNpc. Their brains were perfused with 5% formalin in normal saline via left carotid artery and brain blood vessels washed completely. The brains removed from scalp and immersed in 10% formalin solution for at least 5 days. Data of corrected lesions analyzed statistically (Fig. 1).
Statistical analysis: A paired T-test analysis was used to determine whether significant differences existed in the OVX group's weight at the baseline and one month after ovariectomy.
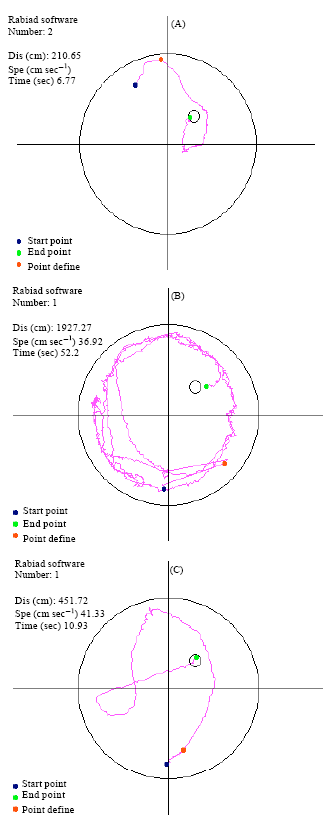 | |
| Fig. 2: | Tracked measures of distance (Dis), speed (Spe) and latency (Time) during spatial training in Morris water maze. (A) Normal rat, (B) Parkinsonian OVX-rat and (C) Parkinsonian OVX-rat with Soy meal |
One-way Analysis of Variance (ANOVA) was run to determine whether group differences existed in terms of percent time spent in the target quadrant and path length during acquisition and probe trials respectively. To further explore the effect of treatment across blocks, separate two-way repeated measures ANOVAs were conducted for each block. One-way Analysis of Variance (ANOVA) was run to determine whether group differences existed in plasma estrogen. All post hoc comparisons were computed using the least significant difference method. P-value less than 0.05, was assumed to denote a significant differences. Tracked measures of distance (Dis), speed (Spe) and latency (Time) during spatial training in Morris water maze is shown in Fig. 2A-C .
RESULTS AND DISCUSSION
Escape latency: As illustrated in Fig. 3A, pre-treated group with normal diet (0), had significantly more percent of total time were spent in target quarter than others (*p<0.05, One-way ANOVA that followed by HSD post hoc test).
Path length: Figure 3B illustrates that pre-treated group with normal diet (0) had significantly longer total path length than others (**p< 0.01, one-way ANOVA and HSD post hoc test).
Probe trials-time: As illustrated in Fig. 4, pre-treated group that received normal diet (0), had significantly lower percent of total time spent in target quarter than others (*p<0.05, one-way ANOVA and HSD post hoc test).
Swim speed: Lesion of SNpc had no significant effect on swim speed during acquisition in the water maze. Also, there were no significant differences between swim speed in all groups during probe trials (p>0.05).
Body weight: Figure 5 shows that body weight at the baseline (weight 1) was significantly lower than at four weeks later (weigth 2) in pre-treated groups (0), (-10) and (-20), respectively (*p< 0.05, one-way ANOVA and HSD post hoc test ).
Plasma estrogen: Figure 6 shows that plasma estrogen level in pre-treated groups (OVX) that received soy (with or without isoflavone) was significantly lower than it in OVX with normal diet (0) and control groups (p<0.001).
Soy isoflavones: Table 1 shows the isoflavone concentration of total soy and alcohol washed soy. Isoflavone concentration was determined by high performance liquid chromatography (HPLC).
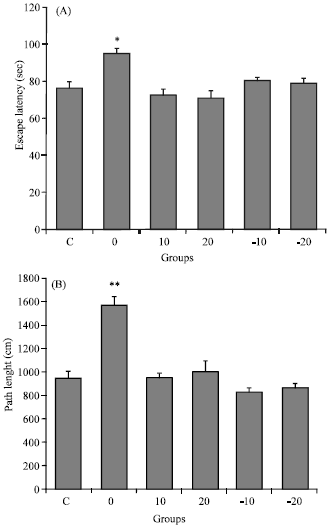 | |
| Fig. 3: | (A) Mean±SEM of total escape latency (sec) to jumping on platform after total acquisition trials in all groups (*p<0.01 vs. other groups) and (B) Mean±SEM total path length to locate on escape platform for total acquisition trials in all groups (**p<0.01 vs. other groups, one-way ANOVA and HSD post hoc test). Control (C); pre-treated groups with normal diet (0); 10 g soy (10); 20 g soy (20); 10 g isoflavone free soy (-10) or 20 g isoflavone free soy (-20) daily for 4 weeks |
The results of the present study indicate that soy consumption apart from containing isoflavone or not in an ovariectomized animal model of Parkinson’s disease improve performance of acquisition in the Morris water maze and prevents body overweight after menopausal life cycle. Soy consumption had positive effect on performance of probe trial, suggesting that soy enhances retention and has memory-enhancing effects. Pre-treated groups receiving soy with or without isoflavone spent significant more percent total time spent in target quadrant while the platform was removed in probe trial than group that has not consume soy in diet.
| Table 1: | The isoflavone concentration (mg 100 mL-1) of total soy and alcohol washed soy that was determined by high performance liquid chromatography (HPLC) |
| *Others: Glycitein, acetyldaidzin, malonylglycitin, malonyldaidzin, genistein | |
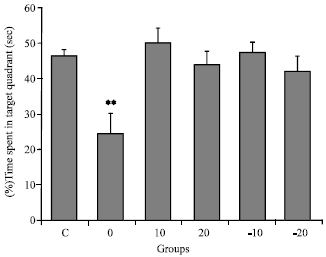 | |
| Fig. 4: | Mean±SEM of percent total time spent in target quarter during probe trial in all groups. Control (C); pre-treated groups with normal diet (0); 10 g soy (10); 20 g soy (20); 10 g isoflavone free soy (-10) or 20 g isoflavone free soy (-20) daily for 4 weeks (**p<0.01 vs. other groups, one-way ANOVA and HSD post hoc test) |
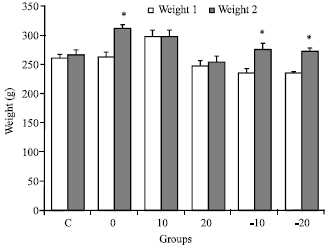 | |
| Fig. 5: | Mean±SEM of body weight at the baseline (weight 1) and four weeks later (weight 2) for all groups: Control (C); pre-treated groups with normal diet (0); 10 g soy (10); 20 g soy (20); 10 g isoflavone free soy (-10) or 20 g isoflavone free soy (-20) daily for 4 weeks (*p<0.05 vs. weight 1, one-way ANOVA and HSD post hoc test) |
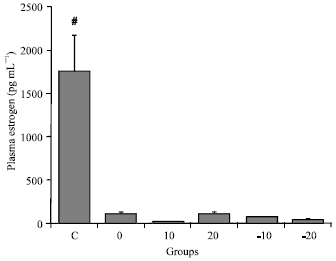 | |
| Fig. 6: | Mean±SEM plasma estrogen level for all groups: control group (C), pre-treated group that received normal diet (0), 10 g soy (10), 20 g soy (20), 10 g isoflavone free soy (-10) or 20 g isoflavone free soy (-20) daily for four weeks (#p<0.001 vs. other groups, one-way ANOVA and HSD post hoc test) |
Mean path length to reach and escape on the platform was longer in (0) group than control and pre-treated groups that received soy (±isoflavone), respectively. These data suggest that lesion of SNpc impairs performance of acquisition in the Morris water maze and soy can prevent acquisition impairment induced by it. It is not clear whether the positive effects of soy that has been seen in this study is due to its isoflavone or other constituents. Data of analyze the soy by HPLC indicated that approximately 15% isoflavone is remained after alcohol washing (Table 1). It is possible; this little amount of isoflavone is responsible for beneficial effects that have seen in our study. Previous studies have shown that at the nanomolar level (5, 10 or 100 nM), genistein has neuroprotective effects. Present findings in this study are consistent with other previous studies (Behl, 1999; Celec et al., 2004; Kritz-Silverstein et al., 2003; Lee et al., 2004; Lund et al., 2001). Short-term high dose of soy bean intake could altered the level of total plasma testosterone and improved spatial cognition in women. Isoflavone supplementation in postmenopausal women diet has a favorable effect on cognitive function particularly verbal memory (Daniel and Lee, 2004). Consumption of dietary phytoestrogens resulting in very high plasma isoflavone levels can significantly alter sexually dimorphic brain regions, anxiety, learning and memory. Dietary Soy derived phytoestrogens can influence learning and memory and alter the expression of proteins involved in neural protection and inflammation in rats. Soy isoflavones can influence the brain cholinergic system and reduce age-related neuronal loss and cognition decline in male rats. Oxidative stress-induced neuronal cell death has been implicated in different neurological disorders and neurodegenerative diseases such as Alzheimer's disease and Parkinson's. These findings imply that the neuroprotective effect of isoflavones extract is partly associated with its antioxidant activity. Further, results of these investigations indicate that although isoflavones extract exert a neuroprotective effect .There are proposed mechanisms for the neuroprotective effects of isoflavones. Genistein, a phytoestrogen that is capable of crossing the blood-brain barrier, has been reported to have an antioxidative effect against the insults of ultraviolet (UV) and chemicals. A large body of evidence indicates that oxidative stress is a salient pathological feature in many neurodegenerative diseases, including Amyotrophic lateral sclerosis, Alzheimer's disease and Parkinson's disease. In addition to signs of systemic oxidative stress, at the biochemical and neuropathological level, neuronal degeneration in these disorders has been shown to coincide with several markers of oxidative damage to lipids, nucleic acids and proteins in affected brain regions. Neuroinflammatory processes, often associated with the induction of free radical generating enzymes and the accumulation of reactive astrocytes and microglial cells, are considered as a major source of oxidative stress given the pathogenic impact of oxidative stress and neuroinflammation, therapeutic strategies aimed to blunt these processes are considered an effective way to confer neuroprotection. Recently, the nuclear transcription factor Nrf2, that binds to the antioxidant response element (ARE) in gene promoters, has been reported to constitute a key regulatory factor in the co-ordinate induction of a battery of endogenous cytoprotective genes, including those encoding for both antioxidant- and anti-inflammatory proteins (Van Muiswinkel and Kuiperij, 2005). In recent years, it has been accepted that oxidative stress is critically involved in the etiopathology of Parkinson's disease (PD) and as a result new therapeutic targets for reduction of oxidant injury and neuroprotection can be defined. Data generated by various groups indicate that the transcription factor nuclear factor erythroid-2-related factor 2 (Nrf2) induces the expression of a group of cytoprotective, antixenobiotic and antioxidant enzymes that include heme oxygenase-1, NAD(P)H: quinone oxidoreductase and enzymes of glutathione (GSH) metabolism such as gamma-glutamyl cysteine ligase, GSH transferases and so on (Cuadrado et al., 2009). Genistein protects cells from H2O2-induced toxicity. H2O2 is a Reactive Oxygen Species (ROS) which can damage the neurons (Holscher, 1998; Morris, 2003; Ramassamy et al., 2000). This antioxidative effect of soy can protects human from neurodegenerative diseases. Previous findings suggest that the mechanisms by which phytoestrogens, especially genistein, protect neuronal cells include not only by the physiological properties of genistein, such as its antioxidative activity, but also activation of estrogen receptors (ERs) and up-regulation of brain-derived neurotrophic factor. Unlike estrogen, genistein, the main soy phytoestrogen, did not trigger proliferation of cells. Because genistein is a selective ER β agonist, it is possible that ER α, but not ER β, mediates the proliferation of endometrium. In the other hand, ER β has a higher level of expression than ER α in brain regions critical to memory function . Administration of 6-OHDA or 1-methyl-4-phenyl-1, 2, 3, 6-tetrahydropyridine (MPTP) to induce animal model of PD, significantly decrease the levels of dopamine (DA) and its metabolites dihydroxyphenylacetic acid (DOPAC) and homovanillic acid (HVA) in the striatum, which could be restored by genistein or estrogen pretreatment (Ferguson et al., 2002). MPTP or 6-OHDA-challenge with genistein or estrogen pre-treatment demonstrated reduced neurotoxicity, with tyrosine hydroxylase-immunoreactive (TH-IR) neurons in the substantia nigra pars compacta (SNpc) affected to a significantly lesser extent as compared to the MPTP or 6-OHDA treated control (Liu et al., 2008).
An evidence indicated that dietary exposure to 500 ppm genistein significantly potentiated amphetamine-stimulated dopamine (DA) release in males and a similar trend was apparent in females, but not statistically significant. These results suggest that dietary genistein exposure may act similarly to estradiol in augmenting amphetamine-stimulated DA release (Pan et al., 1999). In OVX rats, on the other hand, phytoestrogen treatments resulted in a dose-dependent improvement of Visual Spatial Memory (VSM). This improvement in cognitive ability in phytoestrogen treated females may be due in part to the increased presence of choline acetyltransferase messenger RNA in the frontal cortex, which has been shown to be associated with protection and enhancement of cognitive function. Furthermore, phytoestrogens significantly affect the brain calcium-binding protein calbindin (CALB), which acts as a buffer by binding intracellular calcium and plays an important role in mediating cell proliferation, programmed cell death (apoptosis) and neurotoxicity. On the other hand, previous studies show that isoflavones delivered in a soy protein matrix attenuate tau phosphorilations in a primate model of menopause (Kim et al., 2000).
As mentioned earlier, beside isoflavone, other soy constituents may act as neuroprotective agent. Other soy constituents include Protease inhibitors (that have anti-cancer and anti-inflammatory effects), Lignans (that have phytoestrogenic, anti-tumor and anti-viral activity), Comestans (with phytoestrogenic effects), Saponins (that have anti-cancer, antioxidant and anti-mutagenic properties) and Phytates (with antioxidant and anti-cancer effects) may have favorable effects on cognition and body weight alteration during menopausal life cycle (Kritz-Silverstein et al., 2003; Lund and Lephart, 2001).
In conclusion, present findings vin this study suggest the potential usefulness of soy in the improvement of cognition and control of post-menopause body overweight in women. These effects of soy may exert via its potentials as an antioxidant and phytoestrogenic natural substance without any activation of ER α, elevation the plasma level of estrogen and estradiol replacement side effects. Future studies should address the effects of other soy constituents on cognition distinctly.
ACKNOWLEDGMENTS
This study was supported by the research affair of Ahvaz Jondishapour University of Medical Sciences (AJUMS), grant No. 84u47. We wish to thank the AJUMS Physiology Research Center scientists and staff, AJUMS Center Lab Endocrine and Metabolism Research Institute Shaheed Beheshti University of Medical Sciences (Tehran, Iran) for measuring the estrogen of our samples.
REFERENCES
- Baluchnejadmojarad, T., M. Roghani, M.R. Nadoushan and M. Bagheri, 2009. Neuroprotective effect of genistein in 6-hydroxydopamine Hemi-parkinsonian rat model. Phytother. Res., 23: 132-135.
PubMed - Bang, O., H. Hong, D. Kim, J. Boo, H. Kyoon and I. Mook-Jung, 2004. Neuroprotective effect of genistein against beta amyloid-induced neurotoxicity. Neurobiol. Dis., 16: 21-28.
CrossRef - Behl, C., 1999. Alzheimer's disease and oxidative stress: Implications for novel therapeutic approaches. Progr. Neurobiol., 57: 301-323.
CrossRefDirect Link - Bureau, G., F. Longpre and M.G. Martinoli, 2008. Resveratrol and quercetin, two natural polyphenols, reduce apoptotic neuronal cell death induced by neuroinflammation. J. Neurosci. Res., 86: 403-410.
PubMed - Celec, P., D. Ostatnikova, M. Caganova, S. Zuchova and J. Hodosy et al., 2004. Endocrine and cognitive effects of short-time soybean consumption in women. Gynecol. Obst. Invest., 3: 62-66.
CrossRefDirect Link - Cuadrado, A., P. Moreno-Murciano and J. Pedraza-Chaverri, 2009. The transcription factor Nrf2 as a new therapeutic target in Parkinson's disease. Expert. Opin. Ther. Targets., 13: 319-329.
PubMed - Cyr, M., F. Calon, M. Morissette and T. Paolo, 2002. Estrogenic modulation of brain activity: Implications for schizophrenia and Parkinson's disease. J. Psychiatr. Neurosci., 27: 12-27.
Direct Link - Daniel, J.M. and C.D. Lee, 2004. Estrogen replacement in ovariectomized rats affects strategy selection in the Morris water maze. Neurobiol. Learn. Memory, 82: 142-149.
Direct Link - El-Bakri, N.K., A. Islam, S. Zhu, A. Elhassan, A. Mohammed, B. Winblad and A. Adem, 2004. Effects of estrogen and progesterone treatment on rat hippocampal NMDA receptors: Relationship to Morris water maze performance. J. Cell Mol. Med., 8: 537-544.
CrossRefDirect Link - Ferguson, S.A., K.M. Flynn, K.B. Delclos, R.R. Newbold and B.J. Gough, 2002. Effects of lifelong dietary exposure to genistein or nonylphenol on Amphetamine-stimulated striatal dopamine release in male and female rats. Neurotoxicol. Teratol., 24: 37-45.
Direct Link - Gabrieli, J.D., 1996. Memory systems analyses of mnemonic disorders in aging and Age-related diseases. Proc. Natl. Acad. Sci. US., 93: 13534-13540.
Direct Link - Gelinas, S. and M.G. Martinoli, 2002. Neuroprotective effect of estradiol and phytoestrogens on MPP+-induced cytotoxicity in neuronal PC12 cells. J. Neurosci. Res., 70: 90-96.
CrossRefDirect Link - Gerlach, M. and P. Riederer, 1996. Animal models of Parkinson's disease: An empirical comparison with the phenomenology of the disease in man. J. Neural. Transm., 103: 987-1041.
Direct Link - Green, P.S. and J.W. Simpkins, 2000. Neuroprotective effects of estrogens: Potential mechanisms of action. Int. J. Dev. Neurosci., 18: 347-358.
CrossRef - Heikkinen, T., J. Puoliv�li and H. Tanila, 2004. Effects of long-term ovariectomy and estrogen treatment on maze learning in aged mice. Exp. Gerontol., 39: 1277-1283.
CrossRef - Holscher, C., 1998. Possible causes of Alzheimer's disease: amyloid fragments, free radicals and calcium homeostasis. Neurobiol. Dis., 5: 129-141.
CrossRefDirect Link - Kim, H., H. Xia, L. Li and J. Gewin, 2000. Attenuation of neurodegeneration-relevant modifications of brain proteins by dietary soy. Biofactors, 12: 243-250.
Direct Link - Kritz-Silverstein, D., D. von Muhlen, E. Barrett-Connor and M.A. Bressel, 2003. Isoflavones and cognitive function in older women: The soy and postmenopausal health in aging (SOPHIA) study. Menopause, 10: 196-202.
PubMedDirect Link - Kyuhou, S., 2008. Preventive effects of genistein on motor dysfunction following 6-hydroxydopamine injection in ovariectomized rats. Neurosci. Lett., 448: 10-14.
CrossRef - Lee, Y.B., H.J. Lee, M.H. Won, I.K. Hwang, T.C. Kang and J.Y. Lee, 2004. Soy isoflavones improve spatial delayed matching-to-place performance and reduce cholinergic neuron loss in elderly male rats. J. Nutr., 137: 1827-1831.
Direct Link - Lephart, E.D., T.W. West, K.S. Weber, R.W. Rhees, K.D.R. Setchell, H. Adlercreuz and T.D. Lund, 2002. Neurobehavioral effects of dietary soy phytoestrogens. Neurotoxicity Teratol., 24: 5-16.
CrossRefDirect Link - Liu, L.X., W.F. Chen, J.X. Xie and M.S. Wong, 2008. Neuroprotective effects of genistein on dopaminergic neurons in the mice model of Parkinson's disease. Neurosci. Res., 60: 156-161.
PubMed - Lund, T.D., T. West, L. Tain, L. Bu and D. Simmons et al., 2001. Visual spatial memory is enhanced in female rats (but not in males) by dietary soy phytoestrogens. Biomed. Central Neurosci., 2: 1-13.
Direct Link - Markham, J., J.C. Pych and J.M. Juraska, 2002. Ovarian hormone replacement to aged ovariectomized female rats benefits acquisition of Morris water maze. Hormones Behav., 42: 284-293.
CrossRef - Norris, C.M. and T.C. Foster, 1999. MK-801 improves retention in aged rats: Implications for altered neural plasticity in age-related memory deficits. Neurobiol. Learn. Memory, 71: 194-206.
CrossRefDirect Link - Pan, Y., M. Anthony and T.B. Clarkson, 1999. Effect of estradiol and soy phytoestrogens on choline acetyltransferase and nerve growth factor mRNAs in the frontal cortex and hippocampus of female rats. Proc. Soc. Exp. Biol. Med., 221: 118-125.
Direct Link - Paxinos G. and C. Watson, 2008. The Rat Brain in Stereotaxic Coordintes. 6th Edn., Academic Press, New York, pp: 456.
Direct Link - Ramassamy, C., D. Averill, U. Beffert, L. Theroux and S. Lussier et al., 2000. Oxidative insults are associated with apolipoprotein E genotype in Alzheimer's disease brain. Neurobiol. Dis., 7: 23-37.
CrossRef - Shulman, L.M., 2002. Is there a connection between estrogen and Parkinson's disease? Parkinsonism Relat. Disord., 8: 289-295.
PubMed - Van Muiswinkel, F.L. and H.B. Kuiperij, 2005. The Nrf2-ARE Signalling pathway: Promising drug target to combat oxidative stress in neurodegenerative disorders. Curr. Drug Targets. CNS. Neurol. Disord., 4: 267-281.
CrossRefDirect Link - Widy-Tyszekoeiwecz, E., A. Peichal, B. Gajkowska and M. Smailek, 2002. Tellurium-induced cognitive deficits in rats are related to neuropathological changes in the central nervous system. Toxocol. Lett., 131: 203-214.
PubMed