
M. Malek
Physiology Research Center, Department of Physiology, Faculty of Medicine, Ahwaz Jondishapour University of Medical Sciences, Ahwaz, Iran
S. Zahedi Asl
Endocrine Research Center, Research Institute for Endocrine Sciences, Shaheed Beheshti University of Medical Sciences, Tehran, Iran
A. Sarkaki
Physiology Research Center, Department of Physiology, Faculty of Medicine, Ahwaz Jondishapour University of Medical Sciences, Ahwaz, Iran
Y. Farbood
Physiology Research Center, Department of Physiology, Faculty of Medicine, Ahwaz Jondishapour University of Medical Sciences, Ahwaz, Iran
A.H. Doulah
Faculty of Nursing and Midwifery, Ahwaz Branch, Azad Islamic University, Ahwaz, Iran
Pakistan Journal of Biological Sciences
Year: 2009 | Volume: 12 | Issue: 18 | Page No.: 1237-1245







ABSTRACT
The aim of this study was to evaluate the effect of intra-hippocampal injection of Growth Hormone (GH) on impaired spatial cognition in rats with Alzheimer’s Disease (AD). Growth hormone replacement therapy leading to improved cognition and well-being has mainly been carried in GH-deficient patients. Neverthelss, relatively only a few studies have investigated the function of GH in the brain. Aged Wistar male rats (350-400 g, 18-20 months old) were randomly divided into 6 groups (7 in each): Control (healthy aged); L; L+Veh; L+GH10; L+GH20 and L+GH40. Rats with AD-like cognitive deficiency was induced by injection of ibotenic acid into Nucleus Basalis of Meynert (NBM) bilaterally (5 μg 0.5 μL-1, each side). A guide cannula was implanted in the right hippocampus under stereotaxic surgery for injection of human recombinant GH (10, 20 and 40 μg 2 μL-1, during 5 min, twice daily, 9:00 am and 3:00 pm, for 7 days). All rats were trained in Morris water maze to evaluate the spatial learning and memory. Escape latency, traveled distance to find hidden platform and percent time spent in gaol qudrant did not differ between L and L+Veh groups, while latency and distance were reduced significantly. But percent time spent in gaol quadrant (without hidden platform) was increased significantly in NBM-lesioned rats treated with GH (L+GH groups) dose dependently to compare with vehicle treated group. These results suggest that intra-hippocampal injection of GH to aged rats with dementia type of AD (with NBM lesioned) could improve spatial cognition.
PDF Abstract XML References Citation
How to cite this article
M. Malek, S. Zahedi Asl, A. Sarkaki, Y. Farbood and A.H. Doulah, 2009. The Effect of Intra-Hippocampal Injection of Growth Hormone on Spatial Learning and Memory in Animal Model of Alzheimer’s Disease. Pakistan Journal of Biological Sciences, 12: 1237-1245.
DOI: 10.3923/pjbs.2009.1237.1245
URL: https://scialert.net/abstract/?doi=pjbs.2009.1237.1245
DOI: 10.3923/pjbs.2009.1237.1245
URL: https://scialert.net/abstract/?doi=pjbs.2009.1237.1245
INTRODUCTION
Growth Hormone (GH) is a cytokine family hormone which is produced primarily in the anterior pituitary and exerts its effects either directly or indirectly through insulin like growth factor I (IGF-I) (Donahue et al., 2006) on different metabolic and growth processes. Growth hormone is also suggested to be produced endogenously within the hippocampal formation, a brain structure associated with learning and aspect of emotional experiences (Donahue et al., 2002). Synthesis and secretion of GH can be modulated by age, sex and presence of other hormones including estrogen. Studies have demonstrated that GH secretion reaches a peak around the time of the pubertal growth spurt and then declines steadily after 40 years of age with a rate of approximately 14% per decade (Lee et al., 2004). Phenotypic and functional characteristics in old age are comparable to changes seen in GH deficient patients. Nowadays, these manifestations are suggested to be due to a senile decline in GH, termed somatopause, that contribute to frailty and morbidity in old age (Blackman et al., 2002). At the present, decline in high-amplitude GH secretion and plasma IGF-1 concentrations are some of the most robust and well-characterized endocrine alterations that occur with age (Ramsey et al., 2004, 2005). The profound effects of GH on the Central Nervous System (CNS) have become more apparent in the past decade (Frago et al., 2002). Recent studies suggest that the GH-IGF-I axis may play an important role in CNS functions, including those associated with neuronal growth, development, protection (Nyberg, 2000; Scheepens et al., 2000) and certain aspects of mood and cognition (Sonntag et al., 2005; Van Dam and Aleman, 2004). The decline of cognitive function with age coincides with progressive decrease in circulating levels of GH and this decline can be ameliorated through GH treatment (Sun et al., 2005). Direct action of GH on the CNS is supported by the fact that its receptor is expressed in diverse areas of the brain such as choroids plexus, thalamus, hypothalamus, pituitary, putamen and hippocampus, the areas involved in memory and cognition (Creyghton et al., 2004; Lobie et al., 1993; Smith, 2000). This hormone can cross the blood-brain barrier in specific regions (Coculescu, 1999). With Intracerebroventricular (icv) injection of GH, neuronal loss in the frontoparietal cortex, hippocampus and dorsolateral thalamus was reduced while was left unchanged in the striatum (Aberg et al., 2006). This matches the regional distribution of GH receptors and supports the hypothesis that these particular neuroprotective effects are mediated directly by GH and not via IGF-1 (Schneider et al., 2003).
Ageing is associated with certain problems related to neuroendocrine phenomena including Alzheimer’s Disease (AD), dementia and cognitive dysfunction (Rehman and Masson, 2001). The AD is a senile brain disorder with specific pathological changes resulting in neurodegeneration and progressive development of dementia (Fumagalli et al., 2006) and characterized by severe memory loss (Peng et al., 2005). The etiology and pathogenesis of AD are still under intensive investigations (Khachaturian, 2000). Various mechanisms may account for the association between upregulation of the GH/IGF-1 axis and cognitive benefits. Rivastigmine (a selective drug for treatment of AD) and cerebral selective cholinesterase inhibitor (ChEI) (Kennedy et al., 1999), powerfully enhanced the GH release to repeated GHRH stimulation in healthy elderly men dose dependently (Obermayr et al., 2003). This finding suggests that GH administration might be effective in preventing the development or progression of AD (Shin et al., 2005).
Growth hormone also may be used as an exogenous pharmacological agent for enhancing cell genesis in the CNS (Aberg et al., 2006). It has been used in childrens with physical and mental disorders for many years and its therapeutic profile is very well established, therefore it may be worthy to explore its potential in AD (Carro and Torres-Aleman, 2004), Thus, in aged rats we induced lesion in the basal nucleus of Meynert (NBM) to obtain animals with AD-like cognitive deficiency.
MATERIALS AND METHODS
This study was done during April 4, 2008 to February 12, 2009 in neuroscience Lab., Ahwaz Physiology Research Center (PRC), Iran.
Subjects: Aged male Wistar rats (350-400 g, 18-20 months) obtained from Ahwaz Jondishapour University of Medical Sciences (AJUMS), IRAN, animal house center (Toshinai et al., 2007) were housed in a temperature (22±2°C) and humidity-controlled room (55-60%) with a 12:12 h light/dark cycle. Access to food (Pars rat's chow diet factory, Tehran, Iran) and water were ad libitum during the experiments. All experimental procedures were in accordance with Ahwaz Joundishapour academic local ethics commitee for the care and use of laboratory animals. The animals divided randomly into the six groups, 6-7 in each: (1) Control; aged healthy rats (2) L; aged rats with Nucleus Basalis of Meynert (NBM) lesion 3-5) L+GH10, L+GH20 and L+GH40; NBM-lesioned groups that treated intra-hippocampaly with human recombinant GH (10, 20 and 40 μg 2 μL-1, respectively, Novo Nordisk, Bagsvaerd, Denmark, 1.3 mg, that is packaged by Exir Pharmaceutical Co., Iran) and 6) L+Veh; NBM-lesioned rats that received GH related solvent (0.9% Benzyl alcohol and 99.1% normal saline) as a vehicle group.
NBM lesioning: Animals were anesthetized with intraperitoneal (i.p.) injections of Ketamine (100 mg kg-1 b.wt.) and Xylazine (10 mg kg-1 b.wt.) (Alfasan, Woerden-Holland) (Sarkaki et al., 2008). After fixing the head of animals in a steriotaxic instrument (Narishige, Tokyo, Japan), the lesion was induced by injection of ibotenic acid (5 μg 0.5 μL-1 for 5 min in each side, Sigma-Aldrich Chemical Co., USA) dissolved in distillated water into NBM bilaterally (AP; -1.3, L; ±2.3, V; -6.6) coordinates were chosen based on a rat brain atlas (Paxinos and Watson, 2006). Injection was made through 2 μL Hamilton syringe connected to a short piece of polyethylene tube and an injection needle (gage 27). All animals were allowed to recovery period (7-10 days).
Intra-hippocamal injection of GH: A guide cannula (gage 22) was implanted in right hippocampus (AP; -2.3, L; -1.2, V; -3.4) under stereotaxic surgery. Different doses of GH (10, 20, 40 μg 2 μL-1, during 8 min, twice daily, 9:00 am and 3:00 pm, for 7 days) was infused by a 10 μL Hamilton syringe that was connected to an infusion pump (WPI, 101i, USA) through a short piece of polyethylene tube. For all injections, the needle was left in place for a further 3 min to prevent backflow and to allow the infusion. There was no reference for intra-hippocampal injection of GH in animal model of AD. So, we used dose-response procedure (0, 10, 20, 40 μg 2 μL-1 or 0, x, 2x and 4x times of GH).
Training appratus: The water maze was a black circular pool (110 cm in diameter and 70 cm in height) filled with tap water (27°C) with a depth 25 cm. The maze was divided geographically into four equal size quadrants and release points were designed in each quadrant as North (N), East (E), South (S) and West (W). A hidden circular escape platform (12 cm in diameter), was emerged 2 cm below the water level and was located in the center of the northeast quadrant. Some fixed visual cues including computer, desk, shelves, posters and illumination lights were placed on the walls around the pool. A camera was positioned above the center of the pool which was connected to a computer to record the animal motions. An automated tracking system (Radiab ver. 2, Tehran, Iran) was used to measure the escape latency, swimming distance and speed.
Training procedure: From 24 h after the last infusion of GH (8th day) animals were subjected to the training procedure of one session of four trials (block) daily for four consecutive days in the water maze. In each trial, the animals were allowed 60 sec to find the platform, after then were allowed to remain there for 30 sec, if did not find the platform within 60 sec animals were gently guided to the platform. After the completion of a trial, animals were returned to a holding cage for an intertrial interval of 60 sec. After 24 h of the last trial, platform was removed and rats were released from southwest as a probe trial (consisted of a 60 sec free swim period) and the time spent in the target quadrant was recorded (Alaeia et al., 2008).
Histology: At the end of the experiments, the animals were sacrificed with overdose of anaesthetic. Their brains were removed and fixed in 10% formalin for the assessment of the location of the injections. Only data from animals with correct locations of the lesions and injections were used in the analysis (Fig. 4a, b).
GH measurement: Blood samples were prepared from tail vein of rats in both healthy young and aged groups. Their serum GH level (ng mL-1) was measured by ELISA (USCN Life Science and Technology Co., USA).
Statistical analysis: Data expressed as Mean±SEM and analyzed with using two-factor repeated-measures ANOVA, to compare groups in each session, one-way ANOVA to compare groups for total sessions and t-test to compare the serum level of GH between young and aged rats by SPSS (version 15) software package for Windows. Post-hoc Tukey’s test was performed for inter-groups comparisons. The *p<0.05, **p<0.01 and #p<0.001 were considered as significant.
RESULTS AND DISCUSSION
Escape latency: Escape latency to find the hidden platform had no difference between L and L+Veh groups during 1-4 training sessions. The latency was longer significantly in L group than control during 1-4 sessions (*p<0.05 for session 1 and #p<0.001for sessions 2-4, Fig. 1a). Escape latency in L+GH20 Group reduced significantly at 2nd (**p<0.01), 3rd and 4th (#p<0.001) sessions respectively and in L+GH40 reduced significantly at 1st (*p<0.05), 2nd (**p<0.01) and 3rd and 4th (#p<0.001) sessions, respectively when compared to L+Veh group (Fig. 1b, two-factor repeated-measures ANOVA followed by post hoc Tukey’s test). Latency in L+GH10 was reduced significantly at 3rd and 4th sessions compared to L+Veh (**p<0.01). There also was no difference between L and L+Veh groups after all four training sessions, but was significantly longer in L group than control (#p<0.001, Fig. 1c). Latencies after all four sessions in animals treated with 10, 20 and 40 μg 2 μL-1 GH (L+GH10, L+GH20 and L+GH40 groups) were reduced significantly when compared to L+Veh group (**p<0.01 for L+GH10 vs. L+Veh and #p<0.001 for both L+GH20 and L+GH40 groups vs. L+Veh respectively, Fig. 1c, one-way ANOVA followed by post hoc Tukey’s test).
Traveled distance: Traveled distance had no difference between L and L+Veh significantly during 1-4 training sessions. The distance was shorter significantly in control than L group (*p<0.05 and **p<0.01, Fig. 2a). However, in both L+GH20 and L+GH40 groups it was reduced significantly for each day during 2-4 sessions when compared to L+Veh (**p<0.01, Fig. 2b, two-factor repeated-measures ANOVA followed by post hoc Tukey’s test). Mean±SEM of traveled distance after all four sessions had no significant difference between L and L+Veh group, but it increased significantly in L group compared to control rats (#p<0.001, Fig. 2c). Treatment with 10 μg GH (L+GH10 group) could not reduce significantly increased traveled distance due to NBM lesioning, while in both L+GH20 and L+GH40 groups was reduced significantly when compared to L+Veh (**p<0.01, Fig. 2c, one-way ANOVA followed by post hoc Tukey’s test).
Swimming speed: Comparing the swimming speed between the various groups did not indicate any differences during four days spatial training in water maze (p>0.05).
Probe trial: In the probe trial, L group spent less time in the target quadrant significantly when compared to control (#p<0.001). L+GH10, L+GH20 and L+GH40 groups that treated with 10, 20 and 40 μg 2 μL-1 of GH, respectively, spent more time in the target quadrant significantly compared to L+Veh group (*p<0.05 between L+GH10 vs L+Veh and **p<0.01 between both L+GH20 and L+GH40 vs L+Veh, Fig. 3, one-way ANOVA followed by post hoc Tukey’s test). There was no significant difference between L+GH20 and L+GH40 groups with control.
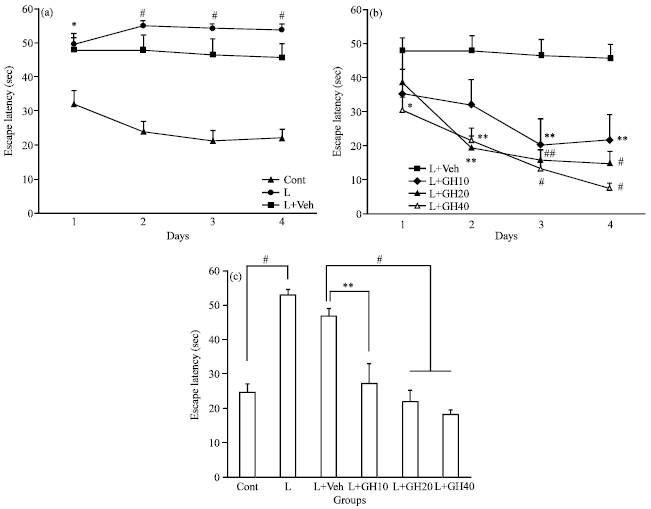 | |
| Fig. 1: | (a, b): Mean ±ESM of escape latency of tested groups during 1-4 sessions. There was no difference between L, L+Veh groups. It was increased in L group compared to control (*p<0.05 for session 1 and # p<0.001 for sessions 2-4). Latency in L+GH20 group was reduced significantly at 2nd (**p<0.01), 3rd and 4th (#p<0.001) sessions and in L+GH40 reduced significantly at 1st (*p<0.05), 2nd (**p<0.01), 3rd and 4th (#p<0.001) sessions, respectively, when compared to L+Veh group (two-factor repeated-measures ANOVA followed by post hoc Tukey’s test) in L+GH10 group it was decreased during sessions 3-4 (**p<0.01). (c): Mean±ESM of escape latency of tested groups after all four training sessions. Latency had no difference between L and L+Veh groups, but was longer in L significantly than control (#p<0.001). However, in all three L+GH10, L+GH20 and L+GH40 groups latency was reduced significantly when compared to L+Veh (**p<0.01 between L+GH10 vs. L+Veh, #p<0.001 between both L+GH20 and L+GH40 vs. L+Veh) |
GH level: Serum GH level was lower in healthy aged (1.09±0.32) than healthy young group (2.18±0.91) (n = 6 in each).
The results of the present study demonstrate that intra-hippocampal injection of GH ameliorates spatial learning and memory deficits in animals serving to model AD in a dose-dependent manner. This finding add more to the previous findings in the favour of the beneficial effect of the GH on spatial learning and memory. To our knowledge this is the first study in its kind that direct administered GH in the hippocampus.
Only a limited number of studies have addressed possible neuroprotective effects of GH administration and most studies have focused on IGF-1, whereas the neuroprotective effect of GH correlates with GH receptor distribution (but not IGF-1 either circulating or locally), this indirectly indicates that GH has some neuroprotective effects itself, independent from IGF-1. The age-related decline in serum GH (as shown above in results) and IGF-1 has been shown to be a contributing factor in some aspects of aging including brain aging, decreased glucose metabolism (Lynch et al., 2001), neurogenesis (Lichtenwalner et al., 2001), angiogenesis (Sonntag et al., 2000), synaptic plasticity (Shi et al., 2005) as well as learning and memory.
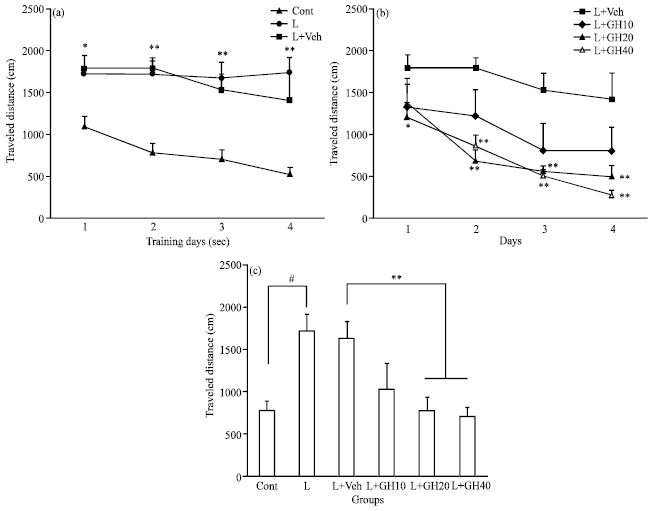 | |
| Fig. 2: | (a): Mean±SEM of traveled distance had no difference between L, L+Veh groups for each session. It increased significantly in L group compared to control during sessions 1-4 (**p<0.01). (b): Traveled distance in both L+GH20 and L+GH40 during sessions 2-4 was reduced significantly compared to L+Veh (**p<0.01). (c): After all four training sessions traveled distance had no difference between L and L+Veh groups. It was longer significantly in L than control (#p<0.001). However, in both L+GH20 and L+GH40 groups was reduced significantly when compared to L+Veh (**p<0.01) |
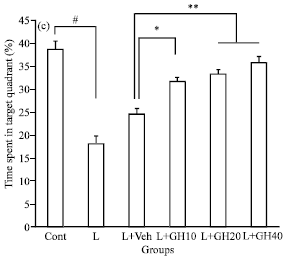 | |
| Fig. 3: | Percent of time spent in target quadrant while escape platform removed during probe trial. #p<0.001 indicate difference between L vs. control. **p<0.01 indicate difference between L+GH20 and L+GH40 groups vs. L+Veh. *p<0.05 indicate difference between L+GH10 group vs. L+Veh |
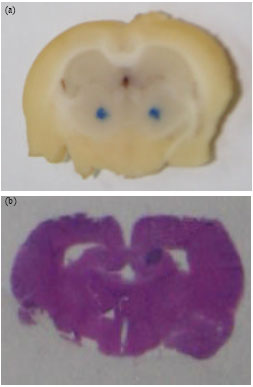 | |
| Fig. 4: | Coronal section of NBM and hippocampal dentate gyrus which histologically stained with cresyl violet to verifying (a) the site of lesion and (b) injection of GH, respectively |
In rodents, a two-fold increase in GH receptors has been observed with age but this increase fails to compensate for the reduction in GH secretion (Xu et al., 1995). A more detailed investigation revealed that the dissociation constant (Kd) and apparent size of the GH receptor were not altered with age (Khan et al., 2002). Our results support the previous studies indicating that GH improves spatial learning and memory in healthy elderly rats and in hypophysectomized male rats after 1 week of daily injection of recombinant human GH (Le Grevès et al., 2002). The findings also support the report by another study which provided evidence for endogenous expression of hippocampal GH and increase in its expression after acquisition of hippocampal-dependent learning task (Donahue et al., 2002). The hypothesis of this study was that since endogenous GH production might be reduced in subjects with AD, so, intra hippocampal GH replacement therapy may have beneficial effect in the disease. In addition, elevation of GH in cerebrospinal fluid of patients treated with GH subcutaneously as a dose dependent support a hypothesize that GH may pass the blood-brain barrier (Burman et al., 1996), another hypothesize in this study is that the decline in GH releasing in elderly subjects can cause less brain GH levels and intra-hippocampal injection of GH can compensate cerebral GH deficiency.
Growth hormone is known to increase energy and motivation but still the swimming speed did not differ (p>0.05) among animals in the different groups. It shows that intra-hippocampal injection of growth hormone had no effect on locomotion system, indicating that observed changes in the latency and traveled distance are not due to changes in the motor function.
Although, the specific mechanisms remain elusive, various mechanisms may account for the association between upregulation of GH/IGF1 axis and cognitive benefits. Both concentration and function of acetylcholine (Ach), an essential neurotransmitter required for learning and memory processing, is decreased in patients with AD (Francis, 2005) not only in the cerebral cortex, but also in subcortical structures, the hippocampus and hypothalamus (Lamperti et al., 1992). Atrophy of cholinergic neurons (up to 90%) in the basal forebrain formation have been well documented in end-stage of AD. It appears that GH and in some extents IGF-1 affects most of the major neurotransmitters differently in several brain regions, including the noradrenergic, dopaminergic, glutaminergic (at least at the receptor level), opoidergic and cholinergic systems. Moreover GH or IGF-1 may be used as exogenous pharmacological agents for enhancing cell genesis in the CNS. An increased plasma level of homocysteine as a strong independent risk factor, may cause dementia and AD (Seshadri et al., 2002) and GH-replacement therapy in GH-deficient subjects decreased fasting homocysteine level (Sesmilo et al., 2001). An increase in aspartate, the ligand of the N-methyl-D-aspartate (NMDA) receptor has been found in the CSF of GH-deficient patients after GH replacement (Burman et al., 1993) and hormone may mediate its memory-promoting effects through an interaction with the NMDA receptor subunits (Le Grevès et al., 2002). This receptor has been shown to be important for synaptic plasticity and also a common switch for many forms of learning and memory (Stevens, 1996; Tang et al., 2001). Furthermore, GH/IGF-1 increases glutamate receptor density in aged rats and there is evidence that GH/IGF-1 may ameliorate age deficits in local cerebral glucose utilization, a function believed to be correlated with neuronal activity and cerebral blood flow (Arwert et al., 2005; Sonntag et al., 1997). In aged rats, subcutaneous administration of growth hormone for 28 days was found to increase microvascular density and blood supply on brain cortical surface. Some evidences showed that GH administration reduces oxidative stress in hippocampus of aged rats by enhancing mitochondrial efficiency, limiting the generation of free radicals, or increasing the activity of enzymes that regulate oxidative stress.
In conclusion, this study for the first time shows that central administration of GH ameliorate impairment of spatial learning and memory in aged rats with AD-like cognitive deficiency and it may be useful in the treatment of men. Surely this usefulness need more and deep human studies.
ACKNOWLEDGMENTS
This study was supported by Physiology Research Center (grant No. PRC-22), Ahwaz Jondishapour University of Medical Sciences (AJUMS) and Endocrine Research Center, Institute of Endocrine and Metabolism, Shaheed Beheshti University of Medical Sciences, Tehran.
REFERENCES
- Aberg, N.D., K.G. Brywe and J. Isgaard, 2006. Aspects of growth hormone and insulin-like growth factor-I related to neuroprotection, regeneration and functional plasticity in the adult brain. Scientific World J., 6: 53-80.
PubMed - Alaei, H., R. Moloudi and A. Sarkaki, 2008. Effects of treadmill running on mid-term memory and swim speed in the rat with Morris water maze test. J. Bodyw. Mov. Ther., 12: 72-75.
CrossRefPubMedDirect Link - Arwert, L.I., D.J. Veltman, J.B. Deijen, A.A. Lammertsma, C. Jonker, M.L. Drent, 2005. Memory performance and the growth hormone/insulin-like growth factor axis in elderly: A positron emission tomography study. Neuroendocrinology, 81: 31-40.
CrossRefDirect Link - Blackman, M.R., J.D. Sorkin, T. Munzer, M.F. Bellantoni and J. Busby-Whitehead et al., 2002. Growth hormone and sex steroid administration in healthy aged women and men: A randomized controlled trial. JAMA, 288: 2282-2292.
Direct Link - Burman, P., J. Hetta and A. Karlsson, 1993. Effect of growth hormone on brain neurotransmitters. Lancet, 342: 1492-1493.
PubMed - Burman, P., J. Hetta, L. Wide, J.E. Mansson, R. Ekman and F.A. Karlsson, 1996. Growth hormone treatment affects brain neurotransmitters and thyroxine [see comment]. Clin. Endocrinol. (Oxf)., 44: 319-324.
PubMed - Carro, E. and I. Torres-Aleman, 2004. Insulin-like growth factor I and Alzheimer's disease: Therapeutic prospects? Expert. Rev. Neurother, 4: 79-86.
PubMed - Coculescu, M., 1999. Blood-brain barrier for human growth hormone and insulin-like growth factor-I. J. Pediatr. Endocrinol. Metab, 12: 113-124.
PubMed - Creyghton, W.M., P.S. van Dam and H.P. Koppeschaar, 2004. The role of the somatotropic system in cognition and other cerebral functions. Semin. Vasc. Med., 4: 167-172.
PubMed - Donahue, C.P., R.V. Jensen, T. Ochiishi, I. Eisenstein, M. Zhao, T. Shors and K.S. Kosik, 2002. Transcriptional profiling reveals regulated genes in the hippocampus during memory formation. Hippocampus, 12: 821-833.
PubMed - Frago, L.M., C. Paneda, S.L. Dickson, A.K. Hewson, J. Argente and J.A. Howen, 2002. Growth hormone (GH) and GH-releasing peptide-6 increase brain insulin-like growth factor-I expression and activate intracellular signaling pathways involved in neuroprotection. Endocrinology, 143: 4113-4122.
Direct Link - Francis, P.T., 2005. The interplay of neurotransmitters in Alzheimer's disease. CNS. Spectr., 10: 6-9.
PubMed - Fumagalli, F., G. Racagni and M.A. Riva, 2006. The expanding role of BDNF: a therapeutic target for Alzheimer's disease? Pharmacogenomics. J., 6: 8-15.
Direct Link - Kennedy, J.S., R.J. Polinsky, B. Johnson, P. Loosen and A. Enz et al., 1999. Preferential cerebrospinal fluid acetylcholinesterase inhibition by rivastigmine in humans. J. Clin. Psychopharmacol., 19: 513-521.
PubMed - Khachaturian, Z.S., 2000. Toward a comprehensive theory of Alzheimer's disease-challenges, caveats, and parameters. Ann. N. Y. Acad. Sci., 924: 184-193.
PubMed - Khan, A.S., D.C. Sane, T. Wannenburg and W.E. Sonntag, 2002. Growth hormone, insulin-like growth actor-1 and the aging cardiovascular system. Cardiovasc. Res., 54: 25-35.
CrossRefDirect Link - Lamperti, E., D. Cocchi, E.A. Parati, T. Caraceni and E.E. Muller, 1992. Growth hormone responses to cholinergically active drugs in patients with dementia of the Alzheimer type. Alzheimer Dis. Assoc. Disord., 6: 44-52.
PubMed - Le Grev�s, M., P. Steensland, P. Le Grev�s and F. Nyberg, 2002. Growth hormone induces age-dependent alteration in the expression of hippocampal growth hormone receptor and N-methyl-D-aspartate receptor subunits gene transcripts in male rats. Proc. Natl. Acad. Sci. USA., 99: 7119-7123.
CrossRefPubMedDirect Link - Lee, K.O., L. Liao and J.J. Mukherjee, 2004. Does growth hormone prevent aging in the healthy elderly with low serum insulin-like growth factor-I ? Mech. Ageing Dev., 125: 291-295.
CrossRefDirect Link - Lichtenwalner, R.J., M.E. Forbes, S.A. Bennett, C.D. Lynch, W.E. Sonntag and D.R. Riddle, 2001. Intracerebroventricular infusion of insulin-like growth factor-I ameliorates the age-related decline in hippocampal neurogenesis. Neuroscience, 107: 603-613.
PubMed - Lobie, P.E., J. Garcia-Aragon, D.T. Lincoln, R. Barnard, J.N. Wilcox and M.J. Waters, 1993. Localization and ontogeny of growth hormone receptor gene expression in the central nervous system. Brain Res. Dev. Brain Res., 74: 225-233.
Direct Link - Lynch, C.D., D. Lyons, A. Khan, S.A. Bennett and W.E. Sonntag, 2001. Insulin-like growth factor-1 selectively increases glucose utilization in brains of aged animals. Endocrinology, 142: 506-509.
Direct Link - Nyberg, F., 2000. Growth hormone in the brain: Characteristics of specific brain targets for the hormone and their functional significance. Front Neuroendocrinol., 21: 330-348.
PubMed - Obermayr, R.P., L. Mayerhofer, M. Knechtelsdorfer, K.H. Tragl and G. Geyer, 2003. The reduced release of GH by GHRH in 8 subjects aged 65-69 years is augmented considerably by rivastigmine, a drug for Alzheimer's disease. Gerontology, 49: 191-195.
CrossRefDirect Link - Peng, S., J. Wuu, E.J. Mufson and M. Fahnestock, 2005. Precursor form of brain-derived neurotrophic factor and mature brain-derived neurotrophic factor are decreased in the pre-clinical stages of Alzheimer's disease. J. Neurochem., 93: 1412-1421.
PubMed - Ramsey, M.M., M.M. Adams, O.J. Ariwodola, W.E. Sonntag and J.L. Weiner, 2005. Functional characterization of des-IGF-1 action at excitatory synapses in the CA1 region of rat hippocampus. J. Neurophysiol., 94: 247-254.
CrossRefDirect Link - Ramsey, M.M., J.L. Weiner, T.P. Moore, C.S. Carter and W.E. Sonntag, 2004. Growth hormone treatment attenuates age-related changes in hippocampal short-term plasticity and spatial learning. Neuroscience, 129: 119-127.
PubMedDirect Link - Rehman, H.U. and E.A. Masson, 2001. Neuroendocrinology of ageing. Age Ageing, 30: 279-287.
Direct Link - Sarkaki, A., R. Assaei, F. Motamedi, M. Badavi and N. Pajouhi, 2008. Effect of parental morphine addiction on hippocampal long-term potentiation in rats offspring. Behav. Brain Res., 186: 72-77.
CrossRefDirect Link - Scheepens, A., C.E. Williams, B.H. Breier, J. Guan, P.D. Gluckman, 2000. A role for the somatotropic axis in neural development, injury and disease. J. Pediatr. Endocrinol. Metab., 6: 1483-1491.
PubMed - Schneider, H.J., U. Pagotto and G.K. Stalla, 2003. Central effects of the somatotropic system. Eur. J. Endocrinol., 149: 377-392.
CrossRefDirect Link - Sesmilo, G., B.M. Biller, J. Llevadot, D. Hayden, G. Hanson, N. Rifai and A. Klibanski, 2001. Effects of growth hormone (GH) administration on homocyst(e)ine levels in men with GH deficiency: A randomized controlled trial. J. Clin. Endocrinol. Metab., 86: 1518-1524.
PubMed - Shi, L., M.C. Linville, E.W. Tucker, W.E. Sonntag and J.K. Brunso-Bechtold, 2005. Differential effects of aging and insulin-like growth factor-1 on synapses in CA1 of rat hippocampus. Cereb. Cortex, 15: 571-577.
CrossRefDirect Link - Shin, E.J., J.H. Jhoo, T. Nabeshima, W.K. Jhoo and M.S. Kwon et al., 2005. Growth hormone releaser attenuates beta-amyloid (1 - 42)-induced memory impairment in mice. J. Pharmacol. Sci., 99: 117-120.
PubMed - Smith, R.G., 2000. The aging process: where are the drug opportunities? Curr. Opin. Chem. Biol., 4: 371-376.
CrossRefDirect Link - Sonntag, W.E., C.D. Lynch, P.T. Cooney and P.M. Hutchins, 1997. Decreases in cerebral microvasculature with age are associated with the decline in growth hormone and insulin-like growth factor 1. Endocrinology, 138: 3515-3520.
Direct Link - Sonntag, W.E., C. Lynch, P. Thornton, A. Khan, S. Bennett and R. Ingram, 2000. The effects of growth hormone and IGF-1 deficiency on cerebrovascular and brain ageing. J. Anat., 4: 575-585.
CrossRefDirect Link - Sonntag, W.E., M. Ramsey and C.S. Carter, 2005. Growth hormone and insulin-like growth factor-1 (IGF-1) and their influence on cognitive aging. Ageing Res. Rev., 4: 195-212.
CrossRefDirect Link - Stevens, C.F., 1996. Spatial learning and memory: the beginning of a dream. Cell, 87: 1147-1148.
PubMed - Sun, L.Y., K. Al-Regaiey, M.M. Masternak, M.M. Wang and A. Bartke, 2005. Local expression of GH and IGF-1 in the hippocampus of GH-deficient long-lived mice. Neurobiol. Ag., 26: 929-937.
PubMed - Tang, Y.P., H. Wang, R. Feng, M. Kyin and J.Z. Tsien, 2001. Differential effects of enrichment on learning and memory function in NR2B transgenic mice. Neuropharmacology, 41: 779-790.
PubMed - Toshinai, K., M.S. Mondal, T. Shimbara, H. Yamaguchi, Y. Date, K. Kangawa and M. Nakazato, 2007. Ghrelin stimulates growth hormone secretion and food intake in aged rats. Mech. Ageing Dev., 128: 182-186.
CrossRefDirect Link - Van Dam, P.S. and A. Aleman, 2004. Insulin-like growth factor-I, cognition and brain aging. Eur. J. Pharmacol, 490: 87-95.
PubMed - Xu, X., S.A. Bennett, R.L. Ingram and W.E. Sonntag, 1995. Decreases in growth hormone receptor signal transduction contribute to the decline in insulin-like growth factor I gene expression with age. Endocrinology, 136: 4551-4557.
Direct Link