
S.R. Mostafavian
Department of Agronomy and Plant Breeding, Sari Higher Education Complex of Agricultural Sciences and Natural Resources, University of Mazandaran, Sari, Iran
H. Pirdashti
Department of Agronomy and Plant Breeding, Sari Higher Education Complex of Agricultural Sciences and Natural Resources, University of Mazandaran, Sari, Iran
M.R. Ramzanpour
Mazandaran Agricultural and Natural Resources Research Center, Sari, Iran
A.A. Andarkhor
Mazandaran Agricultural and Natural Resources Research Center, Sari, Iran
A. Shahsavari
Department of Agronomy and Plant Breeding, Sari Higher Education Complex of Agricultural Sciences and Natural Resources, University of Mazandaran, Sari, Iran
Pakistan Journal of Biological Sciences
Year: 2008 | Volume: 11 | Issue: 6 | Page No.: 826-835







ABSTRACT
A field experiment carried out in a calcareous soil with a low available phosphorus to evaluate effectiveness of biofertilizers, mycorrhizae (Glomus intraradices) and Thiobacillus sp. inoculation individually or in combination on seed yield, oil, protein and some elements (P, Fe, Mn, Zn) concentration in two soybean [Glycine max (L.) Merr.] cultivars. The applied treatments were different fertilizers with 6 levels (including: NP (control, 12 kg N ha 1 as urea, 46 kg P2O5 ha 1 as triple super phosphate); NPK (NP + 75 kg K2O ha 1 as potassium sulphate); NPKS [NPK+ S (100 kg S ha 1)]; NPKST (NPKS + seed inoculation with Thiobacillus bacteria); NPKM (NPK + Seed inoculation with mycorrhizae fungi) and NPKSTM (NPKS + seed inoculation with Thiobacillus and mycorrhizae) and two cultivars (JK and 032). Before planting, soybean seeds were inoculated by Bradyrhizobium japonicum in all treatments. Results showed that combined inoculation of biofertilizers increased yield, however the highest yield was observed in treatment NPKST. Increasing oil content (percentage) was more pronounced in treatments NPKM, while most protein content (percentage) increasing was observed in NPKS and NPKM. Fe and Zn concentrations were unaffected significantly by fertilizer treatments, but NPKSTM showed significantly higher value of seed`s Mn concentration compared to treatments NP and NPK. Although no significant difference was observed in terms of P concentration of 032 line among fertilizer treatments, JK cultivar and NPKSTM caused a significant increasing in P concentration compared to NP, NPKS and NPKM. Present results suggested that applying biofertilizers i.e., mycorrhizae and Thiobacillus increased soybean yield compared to control (NP). Overall, this study demonstrated that soybean seed yield and its chemical composition could be affected by biofertilizer inoculation.
PDF Abstract XML References Citation
How to cite this article
S.R. Mostafavian, H. Pirdashti, M.R. Ramzanpour, A.A. Andarkhor and A. Shahsavari, 2008. Effect of Mycorrhizae, Thiobacillus and Sulfur
Nutrition on the Chemical Composition of Soybean [Glycine max (L.)]
Merr. Seed . Pakistan Journal of Biological Sciences, 11: 826-835.
DOI: 10.3923/pjbs.2008.826.835
URL: https://scialert.net/abstract/?doi=pjbs.2008.826.835
DOI: 10.3923/pjbs.2008.826.835
URL: https://scialert.net/abstract/?doi=pjbs.2008.826.835
INTRODUCTION
During the 19th and early 20th centuries inorganic compounds containing nitrogen, potassium and phosphorus (NPK) were synthesized and used as fertilizers (Loynachan et al., 1993). Due to the growth in human population, fertilizers were used to increase crop production and meet the rising demands for food. Increases in the production cost and the hazardous nature of chemical fertilizers for the environment has led to a resurgence of interest in the used of biofertilizers for enhanced environmental sustainability, lower cost production and good crop yield (Egamberdiyeva, 2007). There is now increasing evidence that these beneficial microbial population (as biofertilizer) can also enhance plant resistance to adverse environmental stress, e.g., water and nutrition deficiency and heavy metal contamination (Wu et al., 2005). Soybean Glycine max (L.) Merr. is an important source of protein and oil. Most of commercially grown soybean cultivars contain about 40% protein and 20% oil (Krishnan et al., 2000). However, both protein and oil concentration can vary greatly with genotype and environment. In order to produce a higher protein meal, in soybean larger amount of N is needed. Because nitrogen is a constituent of amino acids, which are required to synthesize proteins and other related compounds, it plays a positive role in almost all plant metabolic processes (Tucker, 1999). As nitrogen is a limiting factor for soybean productivity, the activity of Bradyrhizobium is extremely important both from economic and environmental viewpoints. Soybean can obtain their total nitrogen needs from the air, when these nitrogen-fixing Rhizobia bacteria are present in sufficient numbers (Panzieri et al., 2000). Normal plant growth cannot achieve without phosphorus. It is a constituent of nucleic acids, phospholipids and the coenzymes, DNA, NADP and the most important ATP. Arbuscular Mycorrhizal Fungi (AMF) are important component of the microbiota, mutualistic symbiotic soil fungi that colonize the roots of most crop plants (Douds et al., 2005). They can equisite and cycle the nutrition (Snyder et al., 2001), have the ability to enhance host uptake of relatively immobile nutrients particularly phosphorus (P) and zinc (Zn) (Abdelhafez and Abdel-Monseif, 2006), Manganese (Mn) (Ardakani et al., 2002) and iron (Fe) have also been reported to be transported in higher amounts towards the plant roots (Azcon et al., 2003). Although AM Fungi are indigenous to most soils, inoculation with these fungi has increased the yield of numerous field-grown crops, including soybean (Bethlenflavayet al., 1997), pepper (Douds and Reider, 2003), tomato (Mohandas, 1978), potato (Duffy and Cassells, 2000) and onion (Sharma and Adholeya, 2000). Moreover, it was demonstrated that mycorrhizae can affect the seed composition. Bethlenflavay et al. (1997) reported that soybean seed protein and oil content affected with mycorrhiza fungi, but not significant relationship was found between seed phosphorus content and protein concentration. On the other hand, there was a negative correlation between seed oil and phosphorus content. Sulfur has long been recognized as an essential element for plant growth. Sulfur is an essential component for synthesis of amino acids (Tucker, 1999) that is required for production of chlorophyll and utilization of phosphorus and other essential nutrients (Kachhave et al., 1997; Tucker, 1999). The microorganisms are mainly responsible to make available sulphate from elemental or reduced forms of sulfur through its oxidation process with the intermediate formation of thiosulphate (S2O32–) and tetrathionate (S4O62–) in soils (Anandham and Sridar, 2004). The role of chemolitotrophic bacteria of the genus Thiobacillus in this process is usually much emphasized (Killham, 1994; Shinde et al., 1996b; Stamford et al., 2002). These bacteria are able to oxidize elemental sulfur to sulfate, a process that results in lower soil pH and nutrients available for plants (Altland, 2006). Sulfur is ranked equal to nitrogen for optimizing crop yield and quality. It enhances the efficiency of nitrogen for protein manufacture (Tucker, 1999). It has been reported that sulphate nutrition through elemental sulfur increased the yield of maize, soybean and wheat (Shinde et al., 1996b). As the sulfur oxidation is a biologic process, high population of oxidizing bacteria is needed. The most and common bacteria are Thiobacillus sp. (Killham, 1994). Anandham and Sridar (2004) concluded that double inoculum of Thiobacillus and Rhizobium increased the oil content of groundnut seed. Other reports showed that combined inoculation of Thiobacillus thioxidans and Aspergillus niger, Trichoderma harzianum, Myrothecium cinctum, Aspergillus terreus with sulfur improved soybean grain yield and protein content (Shinde et al., 2004). Since the chemical composition of crops varied with the crop cultivars, soil and climate conditions of the area, the objective of present study was to evaluate the effect of biofertilizers, mycorrhiza and Thiobacillus on soybean seed composition.
MATERIALS AND METHODS
Study area: The experiment was conducted from May to October, 2006 at Dashtenaz experimental farm of Mazandaran Agricultural and Natural Resources Research Center (Sari, Iran). The research site (36°42`N, 53°13`E, altitude of 16m) is cold and humid in winter, hot and relatively humid in summer, has an average annual precipitation of 564.6 mm with a seasonal range of 10.6- 86.1 mm. During performing this experiment, average relative humidity was about 50-54% and maximum and minimum air temperature were 31.8 and 12.3°C, respectively.
Soil analysis: To determine physicochemical characteristics of soil, a mixed sample of soil from the depth of 0-30 cm was chosen prior to perform this experiment. Soil samples were mechanically analyzed according to the method described by Piper (1950). The Electrical Conductivity (EC) was measured in saturated soil paste based on Jackson (1958) method. Soluble anions, cations and pH were determined in saturated soil paste using method described by Richards (1954). Data of physical and chemical analysis are given in Table 1.
Experimental design and plant sampling: This study assessed the effect of biofertilizers, Thiobacillus and mycorrhizae, in a split plot experiment based on randomized complete block design with three replicates
for each treatment. The treatments were took placed in 2 levels as main factor including Sari (JK-695) cultivar and promising line of 032 (the semi-determinate genotypes, maturity group V) and six levels of fertilizer treatments as sub plot (including NP (control, 12 kg N ha 1 as urea, 46 kg P2O5 ha 1 as triple super phosphate); NPK (NP+ 75 kg K2O ha 1 as potassium sulphate); NPKS [NPK+ S (100 kg S ha 1)]; NPKST (NPKS + Seed inoculation with Thiobacillus bacteria); NPKM (NPK + Seed inoculation with mycorrhizae fungi) and NPKSTM (NPKS + seed inoculation with Thiobacillus and mycorrhizae). Amount of chemical fertilizers were applied according to the soil tests. Bradyrhizobium japonicum with 108 alive cells of bacteria per 1 g of biofertilizer, inoculated to the all treatments before planting. For 20 kg of soybean seed, 250 g mycorrhizae and for each 100 kg sulfur, 250 g Thiobacillus were applied. Soybeans were planted in four rows, spaced 60 cm apart, in 6 m long for each plot. During plant growth season, weed population were manually controlled and sprinkle irrigation was applied when necessary. Soybeans were manually harvested at the maturity, in R8 growth stage (Fehr and Caviness, 1977), with the moisture of 13%, from the two middle rows of each plot and an area about 3 m2 to determine the seed yield (kg ha 1) and 1000 grain weight (g).
Chemical analysis
Elements concentration:Seed samples of each plot were oven dried at 75°C for 48 h. To evaluate seed protein, oil and elements (P, Fe, Mn, Zn) concentration, the oven dried seeds were applied. Dried soybean seeds were ground to a fine powder and were ashed (500°C), digested in 2N HCl. Phosphorus (P) content measured using the vanadate-molybdate-yellow procedure with spectrophotometer (Kacar, 1984) and other elements (Fe, Mn, Zn) by atomic absorption spectrometer (AOAC, 1995). Each sample was analyzed thrice and the mean data are reported herein.
Crude protein analysis:The crude nitrogen of the soy flour ground from the dried seeds was determined by the kjeldahl method (AOAC, 1995), using an auto analyzer [Kjeltec Auto 1030 Analyzer, (Foss Tecator AB, Hoganas, Sweden)] and it was converted to protein content by using the conversion factor 5.71 (FAO, 2003).Table 4 shows the mean value of observations in three independent samples.
Oil extraction: Dried soy flour was extracted with n-hexane in a soxhlet system for 5 h (AOCS, 1997).
Statistical analysis: Data, based on three replications, were analyzed using analysis of variance by SAS statistical software, version 6.12 (SAS, 1997). Duncan`s Multiple Range Test (DMRT) has been applied to make comparisons among means at the 0.05 level of significance.
RESULTS AND DISCUSSION
Oil and protein content: Results in Table 2 show the significant difference between fertilizer treatments in terms of seed oil and protein concentration. Among all fertilizer treatments, NPKM and NPKS revealed the maximum value of protein percentage, while the most value of oil content was related to NPKM (Table 2). Cultivar JK and 032 line showed more percentages of seed oil (19.5%) and protein (37%), respectively (Table 2). Composition of soybean seed can be affected by cultivar (Helms and Orf, 1998), planting date (Helms et al., 1990) and environmental factors (i.e., N, P, K and lime application) (Stark, 1924; Yin and Vyn, 2005). Study the interaction effects of fertilizers and cultivars showed a significant difference among treatments in oil and protein concentration, as the most value of oil concentration were observed in NPKM in 032 line, as well as NPK, NPKS and NPKM treatments in JK cultivar (Table 4). The most value for protein content was belong to mycorrhizal treatment (NPKM) in promising line 032 (Table 4). Data showed the significant increase in protein content in treatments NPKS, NPKST, NPKM and NPKSTM in JK cultivar compared to control. These results are in agreement with Al-Karaki and Clark (1999), suggested that root colonization by AMF may affect protein and lipid composition of plants by altering P nutrition or by eliciting other metabolic responses in the host plant. Variation between two cultivars in their responses to mycorrhiza in seed yield and its composition confirm that genetic variability of plant cultivars can influence the efficiency of arbuscular mycorrhiza isolates (Clark and Zeto, 2000; Linderman and Davis, 2004). Similar report of increasing in N uptake by combined inoculation of Rhizobium and mycorrhiza in soybean was reported by Kumatha et al. (2004). There are many similarities between Rhizobial and VAM symbiosis, which suggest common properties in interaction with plants (Xi et al., 1995). Both microsymbionts are surrounded in the established stage of the symbiosis by plat-derived membranes: the pribacteriod membranes in the infected nodule cells and the prehaustorial membranes around arbuscules in the mycorrhizal roots, respectively. These interfaces are characterized by symbiosis-specific proteins (Perotto et al., 1994). Applying sulfur, individually or in
| Table 2: | Means comparison of grain yield, 1000 grain weight, oil and protein yield and concentration affected by cultivar and fertilizer in soybean (n = 3) |
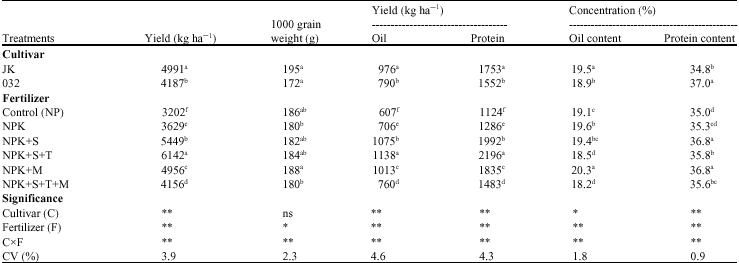 | |
| N = Nitrogen; P = Phosphorus; K = Potassium; S = Sulfur; T = Thiobacillus; M = Mycorrhizae, ns, * and ** means not significant, significant at 5% and 1% probability levels, respectively, n = No. of replications, Means value in each column with the same letter(s) do not differ significantly by DMRT (p = 0.05) | |
combination with Thiobacillus had stimulating effect on seed yield and seed protein concentration. These results are in compatible with previous findings which demonstrated the positive effects of inoculation of sulfur oxidizing bacteria like Thiobacillus sp. and some fungus on grain yield and grain protein content in wheat (Shinde et al., 1996a) and soybean (Shinde et al., 2004). As a result, the natural oxidation of elemental sulfur caused to increase soil sulphate content. The availability of SO42 is important for protein synthesis in plant. Seed protein production puts a large demand on the soybean`s vegetative tissues for mobilization of stored nitrogen (N) (Shibles and Sundberg, 1998) and sulfur (Anderson and Fitzgerald, 2001). Sulfur is first delivered to the aerial portion of plants as SO42 via xylem (Sunarpi and Anderson, 1996). Smith and Lange (1988) reported that sulfur is transported to mature leaves and exported, unchanged, quickly via phloem. These mature leaves appear to act as a transfer station for sulfur. Barley (Hordeum vulgare L.) matured leaves import SO42 , but this is remaining in a pool isolated from endogenous S. The later was probably primarily in the form of protein or sulfolipid (Naeve and Shibles, 2005).
Oil and protein yield: Although oil concentration decreased by applying Thiobacillus in treatment NPKST and NPKSTM in 032 line compare to NP, the overall oil yield (kg ha 1) increased because of the large increase of seed yield as a result of Thiobacillus inoculation (Table 4). The large increases in seed yield compensated the decline in oil concentration. Increasing in oil content by application of sulfur with Thiobacillus (oxidizing bacteria) was suggested by Anandham and Sridar (2004) in groundnut, however, our results did not agree with their finding. No effect of sulfur on seed oil concentration was reported in other soybean study (Seguin and Zheng, 2006). Significant varieties of seed compositions for same cultivar, when grown in different years or under different environments in the same year, have been observed (Yin and Vyn, 2005). Protein yield (kg ha 1) showed a significant difference among the treatments. The most protein yield was observed in treatment NPKST, because of the large increase in seed yield (as a result of Thiobacillus inoculation with sulfur) (Table 4). The coefficient correlation between seed yield and oil yield (r = 0.98**) and protein yield (r = 0.99**) were significantly positive, referring to Table 6, no significant correlation were observed between seed yield and oil (r = 0.19ns) and protein (r = -0.18ns) concentration. Although oil concentration in treatment with high seed yield were statistically lower than those under low and medium yield (Table 4), in general our results are in agreement with those of Stark (1924), who reported that an increase in soybean seed yield was frequently associated with a statistically significant but small decrease in oil concentration. However, our results disagreed with his finding that revealed soybean yield was positively associated with seed protein concentration, other soybean studies, found a negative (Burton, 1985) and not significant correlation between seed yield and seed protein concentration (Yin and Vyn, 2005). The differences among all these studies confirmed, soybean seed composition can be affected by cultivar, management practices and environmental factors.
Seed elements: Table 3 shows that elements concentration in seed did not affected by fertilizer treatments and cultivar×fertilizer interaction. In addition there was no significant difference between cultivars except for manganese (Mn) concentration, which was significantly higher in promising line 032 than Jk (Table 3). Variation between cultivars element concentrations in seed supported other studies out comes of existing genetic variability for the accumulation of mineral elements in soybean germplasm (Caires et al., 2006). The maximum values of phosphorus concentration in seed were observed in NPK, NPKST and NPKSTM treatments in JK cultivar (Table 5). The most Mn concentration in seed was found in NPKST, NPKM and NPKSTM treatments in 032 line (Table 5). It should be considered that positive effects of mycorrhiza on increasing phosphorus and other elements nutrition were found in soil nutrient deficiency (Bethlenflavay et al., 1997; Wu et al., 2005) or drought stress (Al-Karaki and Al-Radad, 1997; Subermanian and Charest, 1999). Current study can not exactly support above researches, although they determined root, shoot and leaves nutrients concentration in host plant and less seed concentration was reported. It might be due to soil fertility and loam texture, which was not such poor in nutrition. This conclusion is in line with most other findings which reported that extensive colonization occurs mainly in plants growing in soils of low fertility (Gehring and Whitham, 1994). Moreover, field and greenhouse studies demonstrated that crops growing in nutrient-poor soils had higher levels of mycorrhizal colonization than crop growing in rich soils (Gehring and Whitham, 1994). Our findings were similar to several studies which were conducted by plants grown under normal seedling growing conditions and differed from those which conducted under stress condition (Sensoy et al., 2007). Similar results have been found on soybean (Khalil et al., 1994) and solanum (Diop et al., 2003) cultivars indicating that P uptake by mycorrhizal plants fluctuates with fungal isolates and genetic variability between cultivars. Although there was not difference in seed nutrients concentration, mycorrhizal plants showed significantly higher yield than NP and NPK (Table 4). It can be concluded that influence of mycorrhiza symbiosis on plant development maybe totally unrelated to plant mineral physiology (Smith and Gianinazzi-Pearson, 1988). It is important to note that, while mycorrhizal association may be beneficial, they do not necessarily enhance P uptake sufficiently to maximize crop yield. Ryan and Ash (1999, 2000) reported that in spite of enhanced mycorrhizal association in biodynamic pastures, the level of P in the forage was below than conventionally fertilized pasture. In other studies in Australia, neither pea nor autumn-sown wheat showed a benefit in yield or P uptake from enhanced mycorrhizal colonization, even under low-P conditions, possibly due to growing season temperature or moisture regime which affect crop growth rate and P
| Table 3: | Means comparisons of seed element concentration affected by cultivar and fertilizer in soybean (n = 3) |
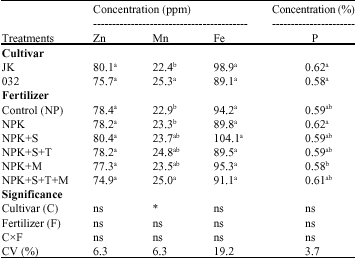 | |
| n = No. of replications, Means value in each column with the same letter(s) do not differ significantly by DMRT (P = 0.05).*N = Nitrogen; P = Phosphorus; K = Potassium; S = Sulfur; T = Thiobacillus; M = Mycorrhizae, ns, * and ** means not significant, significant at 5 and 1% probability levels, respectively | |
| Table 4: | Mean comparison of seed traits affected by cultivar and fertilizer in soybean (n = 3) |
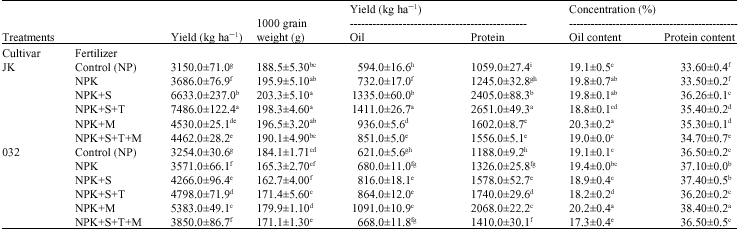 | |
| n = No. of replications, Means±SE value in each column with the same letter(s) do not differ significantly by DMRT (p = 0.05) *N = Nitrogen; P = Phosphorus; K = Potassium; S = Sulfur; T = Thiobacillus; M = Mycorrhizae | |
| Table 5: | Mean comparison of seed traits affected by cultivar and fertilizer in soybean (n = 3) |
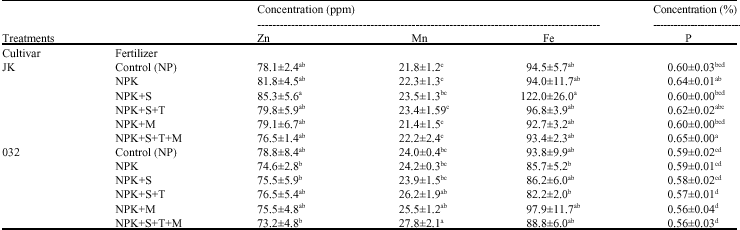 | |
| n = No. of replications, Means±SE value in each column with the same letter(s) do not differ significantly by DMRT (p = 0.05) *N = Nitrogen; P = Phosphorus; K = Potassium; S = Sulfur; T = Thiobacillus; M = Mycorrhizae | |
availability or to the type of AM fungal community present (Ryan and Angus, 2003). Studies showed the increasing uptake of zinc (Zn) (George, 2000;Turk et al., 2006) and iron (Fe) (Purakayastha et al., 1998;Azcon et al., 2003) uptake in mycorrhizal plant, while in other studies decreasing uptake of Zn (Goh et al., 1997), Fe (Tarafdar and Marschner, 1994) and not significant effect on Fe uptake with mycorrhizal plant (Caris et al., 1998) were reported. Our finding showed not significant effect of mycorrhiza on Fe and Zn concentration in seed compared to non mycorrhizal treatments. Not significant difference in Fe concentration in seed might be due to seed inoculation with Bradyrhizobium japonicum bacteria for all treatments at present study that was lead to nodulation. It was reported that nodulated legume under field condition facilitate uptake of iron (Fe) from the environment, by production of sidrophores (Schippers et al., 1987). Results in Table 5 showed a higher value of Fe concentration in NPKS treatment and JK cultivar compared to NPK and NPKST treatments in 032 line and Zn concentration compared to NPK and NPKS treatments in 032 line. The positive effect of sulfur on increasing Fe uptake is in agreement with Kumawat et al. (2006) that found the same result in mungbean (Vigna radiata L.). These results are in agreement with other studies reported that quality and quantity of crops yield affected by sulfur in sorghum (Kaplan and Orman, 1998) sunflower (Gangardhara et al., 1990) and common bean (Liuch et al., 1983). Diverse response of two cultivar in sulfur application supply, was consistent with others studies that demonstrated soybean cultivars were different in elements accumulation (Caires et al., 2006). Frequently mycorrhizal plants showed a decrease in Mn concentration (Heggo and Barakah, 1993; Posta et al., 1995), but our results showed an increase in mycorrhizal treatments in 032 line (Table 5). The present study is in agreement with Nogueira and Cardoso (2003) that found Mn availability increased in the clay soil with AMF inoculated soybean and expressed increasing of Mn availability resulted in the increase of Mn concentration in the plant. Moreover, Bethlenflavay and Franson (1989) observed that, although mycorrhizal plants presented greater Mn concentrations, there were no toxicity symptoms, this might have occurred because of an increase of internal tolerance to Mn (Foy et al., 1978) by plants better fed with P (Nogueira and Cardoso, 2003). Although seed elements, oil and protein concentration with yield have not shown a significant correlation, a positive and significant correlation between 1000 grain weight and Fe concentration (r = 0.48**), Zn concentration (r = 0.42*) and oil concentration (r = 0.42*), also a negative and significant correlation between 1000 grain weight and seed protein concentration (r = -0.48**) and seed Mn concentration (r = -0.50**) were found (Table 6). The relation among elements showed significantly negative correlation between Zn (r = -0.39*) and Mn, also between P (r = -0.46**) and Mn (Table 6). These results are in line with other reports that confirmed the negative correlation between Mn, Zn and P (Malakouti and Gheibi, 1999; Hashemi-Rad and Karimian, 2001). Ozkutlu et al. (2006) also expressed that in a calcareous soil, zinc deficiency resulted in higher concentration of Mn and Fe in soybean and wheat; moreover, adding Zn to soil was effective in reducing Mn and Fe concentration in shoots of both plants. In addition, they concluded that these reductions of Mn and Fe are related to the dilution effect caused by increasing shoot growth. Seed phosphorus (P) concentration and oil concentration were not significantly correlated (r = 0.05), but a significant and negative correlation (r = -0.48**) between seed protein concentration and phosphorus (P) concentration was observed (Table 6). There are contradictory reports of relation between seed phosphorus concentration and oil and protein
| Table 6: | Pearson`s correlation matrix among studied traits in soybean (n = 36) |
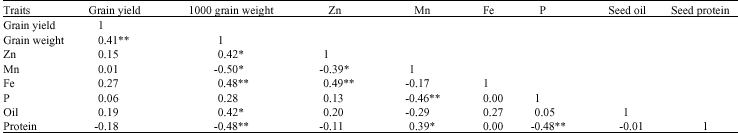 | |
| *,** Means significant at 5 and 1% probability levels, respectively | |
concentration. Bethlenflavay et al. (1997) have proposed that seed P and protein concentrations were not significantly correlated (r = 0.162), but a highly negative significant correlation (r = -0.95**) between seed P and lipid concentrations was observed.
CONCLUSION
According to the above results, inoculation with VAM fungi increased seed oil and protein content of soybean. Seed element concentration did not affected by fertilizers and most importantly, VAM and Thiobacillus inoculation alone and in combination increased seed yield of soybean compared to control. However dual inoculation of Thiobacillus and mycorrhizae revealed less seed yield increase compared to each in separate. It might be the result of the interaction of two microorganisms. Considering the wide variety of responses to biofertilizer inoculation for plant cultivars in same or different environment, as demonstrated in this and other studies, it would be suggested to study the effect of biofertilizers with different amount of P and S fertilizers and other cultivars.
ACKNOWLEDGMENTS
We would like to express our thanks to Abbas-Ali Rostamzad and Ebrahim Khandan for excellent technical assistance with seed analysis and Dr. Gholam-Ali Ranjbar for critical review of the manuscript.
REFERENCES
- Abdelhafez, A.A.M. and R.A. Abdel-Monseif, 2006. Effects of VA mycorrhizal inoculation on growth, yield and nutrient content of cantaloupe and cucumber under different water regimes. Res. J. Agric. Biol. Sci., 2: 503-508.
Direct Link - Al-Karaki, G.N. and R.B. Clark, 1999. Mycorrhizal influence on protein and lipid of durum wheat grown at different soil phosphorus levels. Mycorrhiza, 9: 97-101.
CrossRefDirect Link - Anderson, J.W. and M.A. Fitzgerald, 2001. Physiological and metabolic origin of sulfur for the synthesis of seed storage proteins. J. Plant Physiol., 158: 447-456.
Direct Link - Azcon, R., E. Ambrosano and C. Charest, 2003. Nutrient acquisition in mycorrhizal lettuce plants under different phosphorus and nitrogen concentration. Plant Sci., 165: 1137-1145.
Direct Link - Caires, E.F., S. Churka, F.J. Garbuio, R.A. Ferrari and M.A. Morgano, 2006. Soybean yield and quality as a function of lime and gypsum applications. Sci. Agric. Piracicaba, Brazil, 63: 370-379.
Direct Link - Diop, T.A., T. Krasova-Wade, A. Diallo, M. Diouf and M. Gueye, 2003. Solanum cultivar responses to arbuscular mycorrhizal fungi: Growth and mineral status. Afr. J. Biotechnol., 2: 429-433.
Direct Link - Douds, D.D. and C. Reider, 2003. Inoculation with mycorrhizal fungi increases the yield of green peppers in a high P soil. Biol. Agric. Hort., 21: 91-102.
Direct Link - Douds, Jr. D.D., G. Nagahashi, P.E. Pfeffer, C. Reider and W.M. Kayser, 2006. On-farm production of AM fungus inoculum in mixtures of compost and vermiculite. Bioresour. Technol., 97: 809-818.
CrossRefDirect Link - Duffy, E.M. and A.C. Cassells, 2000. The effect of inoculation of potato (Solanum tuberosum L.) microplants with arbuscular mycorrhizal fungi on tuber yield and tuber size distribution Applied Soil Ecol., 15: 137-144.
CrossRefDirect Link - Egamberdiyeva, D., 2007. The effect of plant growth promoting bacteria on growth and nutrient uptake of maize in two different soils. Appl. Soil Ecol., 36: 184-189.
CrossRefDirect Link - Foy, C.D., R.L. Chaney and M.C. White, 1978. The physiology of metal toxicity in plants. Ann. Rev. Plant Physiol., 29: 511-566.
CrossRefDirect Link - Khalil, S., T.E. Loynachan and M.A. Tabatabai, 1994. Mycorrhizal dependency and nutrient uptake by improved and unimproved corn and soybean cultivars. Agron. J., 86: 949-958.
CrossRefDirect Link - Kumawat, R., R. Praveen, N. Narayan and M. Mahesh, 2006. Effect of sulfur and iron on enzymatic activity and chlorophyll content of mungbean. J. Plant Nutr., 29: 1451-1467.
Direct Link - Naeve, S.L. and R.M. Shibles, 2005. Distribution and mobilization of sulfur during soybean reproduction. Crop Sci., 45: 2540-2551.
Direct Link - Nogueira, M.A. and E.J.B.N. Cardoso, 2003. Mycorrhizal effectiveness and manganese toxicity in soybean as affected by type and endophyte. Sci. Agricola, 60: 329-335.
Direct Link - Panzieri, M., N. Marchettini and T.G. Hallam, 2000. Importance of the Bradyrhizobium japonicum symbiosis for the sustainability of soybean cultivation. Ecol. Model., 135: 301-310.
Direct Link - Ryan, M. and J. Ash, 2000. Phosphorus controls the level of colonization by arbuscular mycorrhizal fungi in conventional and biodynamic irrigated dairy pasture. Aust. J. Exp. Agric., 40: 663-670.
Direct Link - Ryan, M.H. and J.F. Angus, 2003. Arbuscular mycorrhizae in wheat and field pea crops on a low P soil: Increased Zn-uptake or yield. Plant Soil, 250: 225-239.
Direct Link - Sensoy, S., S. Demir, O. Turkmen, C. Erdinc and O.B. Saur, 2007. Responses of some different pepper (Capsicum annum L.) genotypes to inoculation with two different arbuscular mycorrhizal fungi. Sci. Hortic., 113: 92-95.
CrossRef - Sharma, M.P. and A. Adholeya, 2000. Enhanced growth and productivity following inoculation with indigenous AM fungi in four varieties of onion (Allium cepa L.) in an alfisol. Biol. Agric. Horticult. Int. J. Sustain. Prod. Syst., 18: 1-14.
CrossRefDirect Link - Stamford, N.P., A.J.N. Silva, A.D.S. Freitas, J.T. Araujo Filho, 2002. Effect of sulfur inoculated with Thiobacillus on soil salinity and growth of tropical tree legumes. Bioresour. Technol., 81: 53-59.
Direct Link - Tarafdar, J.C. and H. Marschner, 1994. Efficiency of VAM hyphae in utilization of organic phosphorus by wheat plants. Soil Sci. Plant Nutr., 40: 593-600.
CrossRef - Xie, Z.P., C. Staehelin, H. Vieheilig, A. Wiemken and S. Jabbouri et al., 1995. Rhizobial nodulation factors stimulate mycorrhizal colonization of nodulating and nonnodulating soybean. Plant Physiol., 108: 1519-1525.
PubMedDirect Link - Yin, X. and T.J. Vyn, 2005. Relationships of isoflavone, oil and protein in seed with yield of soybean. Agron. J., 97: 1314-1321.
Direct Link - Wu, S.C., Z.H. Cao, Z.G. Li, K.C. Cheung and M.H. Wong, 2005. Effects of biofertilizer containing N-fixer, P and K solubilizers and AM fungi on maize growth: A greenhouse trial. Geoderma, 125: 155-166.
CrossRefDirect Link