
Roohollah Ramezani
Department of Biology, Faculty of Sciences, Ferdowsi University of Mashhad, Iran
Ali Moghimi
Department of Biology, Faculty of Sciences, Ferdowsi University of Mashhad, Iran
Hassan Rakhshandeh
Department of Physiology and Pharmacology, Qaem Hospital,
Mashhad University of Medical Sciences, Iran
Hamid Ejtehadi
Department of Biology, Faculty of Sciences, Ferdowsi University of Mashhad, Iran
Masoumeh Kheirabadi
Department of Biology, Faculty of Sciences, Ferdowsi University of Mashhad, Iran
Pakistan Journal of Biological Sciences
Year: 2008 | Volume: 11 | Issue: 5 | Page No.: 746-751







ABSTRACT
We investigated the effect of Rosa damascena Mill, essential oil on the development of induced amygdala kindling seizures. Male Wistar rats were implanted with one tripolar and two monopolar electrodes in right basolateral amygdala and dura surface, respectively. The control group was injected solvent of essential oil and two experimental groups were injected 750 and 1000 mg kg-1 of essential oil (ip), 30 min before a daily kindling stimulation. The number of stimulations required for the first appearance of seizure stages was significantly larger in two experimental groups than in control group. Mean after discharge duration was significantly different and essential oil reduced the increase of after discharge duration. Mean after discharge amplitude was also shorter in the groups treated with essential oil than in control group. Duration time for 5th stage of seizure at fully-kindled rats was significantly shorter in two experimental groups than control group. These results suggest that Rosa damascena essential oil significantly retarded the development of seizure stages and possesses the ability to counteract kindling acquisition. The flavonoids of Rosa damascena may act via GABAA receptors as previous studies have proposed for flavonoids of other medicinal plants. More detailed studies are recommended to define the effective component(s) of Rosa on different types of epilepsy.
PDF Abstract XML References Citation
How to cite this article
Roohollah Ramezani, Ali Moghimi, Hassan Rakhshandeh, Hamid Ejtehadi and Masoumeh Kheirabadi, 2008. The Effect of Rosa damascena Essential Oil on the Amygdala Electrical Kindling Seizures in Rat. Pakistan Journal of Biological Sciences, 11: 746-751.
DOI: 10.3923/pjbs.2008.746.751
URL: https://scialert.net/abstract/?doi=pjbs.2008.746.751
DOI: 10.3923/pjbs.2008.746.751
URL: https://scialert.net/abstract/?doi=pjbs.2008.746.751
INTRODUCTION
Epileptic patients have impaired physical, psychological and social functioning, which may lead to economic loss and diminished quality of life. Epilepsy, a chronic disorder of the nervous system affecting 1% of the world population, is characterized by the abnormal synchronized firing of a large number of neurons. The large synchronized event is known as a seizure, paroxysmal discharge, or ictal event (Durand and Bikson, 2001; Smedt et al., 2005). Experimental epilepsy studies have showed enhancement of transmission in excitatory systems, particularly glutamatergic systems and the enhancement mechanisms such as elevated presynaptic neurotransmitter release, increased postsynaptic receptor sensitivity and altered glutamate metabolism and failure of inhibitory systems such as GABAergic systems can lead to seizure activity. Temporal Lobe Epilepsy (TLE) is the most common and drug-resistant type of adult focal epilepsy. The epileptic focus in TLE patients often resides in mesial temporal structures, such as the hippocampus or amygdala (or both) and the seizures are complex partial (Morimoto et al., 2004). Goddard reported in 1967 that repeated electrical stimulation of the rat amygdala caused an epileptiform response that increased progressively in severity. This increase in epileptiform response is referred to as kindling and has subsequently become widely accepted as a model of temporal lobe epilepsy (Michael et al., 1998). Kindling is a phenomenon whereby repetitive, focal sub convulsive stimulation ultimately results in intense partial and generalized convulsive seizures (Sarkisian, 2001). The amygdala is the most sensitive structure for the induction of kindling (Shea et al., 2000; Yamashita et al., 2004). In amygdala kindling the convulsive seizures appear in frontal motor cortex and initially became bilateral through callosal projections to the contralateral motor cortex (Coulter et al., 2002). Researches to find out antiepileptic drugs with minimal side effects are going in many laboratories for decades but yet we need to find out more effective medications capable of diminishing various symptoms of seizures. Therefore, it is recommended to use new chemicals with natural origins. Rosa damascena is one of the most important Rosa species for the flavor and fragrance industries that has been traditionally used for centuries as an odor with relaxant and anxielytic effects. In Iranian tradition Rosa leaves are using as choleretic and laxative (Zargari, 1992) and several therapeutic effects such as hypnotic (Rakhshandeh et al., 2004), antispasmodic and relaxant (Boskabady et al., 2005), treatment of abdominal and chest pain and also inotropic effects on the heart have been described for the flowers of Rosa damascena. Recent studies show that Rosa damascena has anti-HIV (Mahmood et al., 1996), antioxidant and antibacterial activities (Basim and Basim, 2003), hepatoprotective (Achuthan et al., 2003) and antitussive effects (Shafei et al., 2003). The toxicological studies of Rose essential have confirmed its safety (Tisserand, 1995). In the present study, we investigated the anticonvulsant effects of Rosa damascena essential oil on the development of amygdala kindling seizures in Wistar rats.
MATERIALS AND METHODS
Animals: Male Wistar rats (300-350 g) were used throughout this study. The animals were housed in standard Plexiglas cages with free access to food (standard laboratory rodent`s chow) and water. The animal house temperature was maintained at 24-25°C with a 12/12 h light/dark cycle, light on at 7:00 am to 7:00 pm.
Stereotaxic surgery: The rats were anesthetized with ketamine (60 mg kg-1, i.p.) and xylazine (10 mg kg-1, i.p.). Then, they were stereotaxically implanted with bipolar stimulating and monopolar recording stainless steel (Advent Ltd) Teflon-coated electrodes (twisted into a tripolar configuration), in the basolateral amygdala (coordinates: A = 2.5 mm from bregma; L = 4.8 mm from bregma and DV = 7.5 mm from dura) of the right hemisphere and Two screw electrodes (earth and differential) were placed touching in dura surface (Paxinos and Watson, 1986). The electrodes were fixed to the skull with dental acrylic. The animals were given 10 days for recovery after surgery, before the kindling protocol was started.
Kindling procedure: Ten days after surgery, electrical kindling was begun by determination of the After Discharges (AD) threshold in each animal. Kindling stimulation (2-s, 100 Hz, biphasic square wave pulses of 0.5 ms per half waves), was initially delivered at 10 µA, with an ascending series of 10 µA at 5 min intervals, until at least 5 s AD was recorded (Saberi and Pourgholami, 2003). In this study, animals were stimulated daily. Convulsive stages were identified according to paradigm of (Racine, 1972): Stage 0, no response or motor arrest; Stage 1 (S1), facial or jaw movements; Stage 2 (S2), addition of head nodding; Stage 3 (S3), unilateral forelimb clonus; Stage 4 (S4), rearing with bilateral forelimb clonus and Stage 5 (S5), rearing, forelimbs clonus and loss of equilibrium. The animals were considered kindled after two consecutive stage 5 of convulsions.
Animals were cared and handled in accordance with the Iranian society of animal care (member of international animal care society) and also local institutional ethics for animal care.
Preparing the essential oil of Rosa damascena: The Rosa leaves were prepared from the fields of Golabe Nader Co (Iran), after identifying the species of the plant by the botanical specialists of the biological department of Ferdowsi Univ. (Mashhad, Iran). The essential oil was isolated from the leaves of the plant by steam distillation (hydro-distilled) according to the standard method in the European pharmacopoeia and explained method (Tyler et al., 1988). This part of procedure was conducted during spring season in the physiology and pharmacology Department of Ghaem Hospital, Mashhad Med. Univ.
Effects of Rosa damascena essential oil on the development of kindling: To investigate oil rose effects on kindling development, rats were randomly assigned to a control group and two experimental groups (n = 8 for each group). The control group was injected solvent of rose essential oil (normal saline and tween) and two experimental groups were injected 750 and 1000 mg kg-1 of rose essential oil, intraperitonealy, 30 min before a daily kindling stimulation. This procedure was repeated once daily until full kindling was reached. While After Discharges Duration (ADD) was recorded after each stimulation, convulsive stage and responses were identified as described above.
Histology: At the end of the experiments, rats were killed with an overdose of ether. Their brains were removed, sectioned and examined under microscope for electrode tip placement verification. If any abnormalities were seen, the data from the particular animal were not included in the results.
The experiments were conducted in the Physiology research lab of the biology department of Ferdowsi University of Mashhad.
Statistical analysis: The dose-response data were evaluated by a one-way analysis of variance and an a priori test for multiple comparisons of means (Tukey HSD test) was used to compare individual treatment groups.
RESULTS
The number of stimulations required for the first appearance of 1st, 2nd and 3rd stages of seizure was significantly larger in two experimental groups than
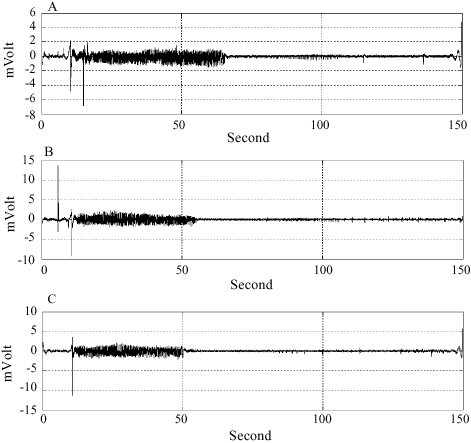 | |
| Fig. 1: | Evoked after discharge of 5th stage at fully-kindled rats. (A) control group, (B) group treated with 750 mg kg-1 of rose essential oil and (C) group treated with 1000 mg kg-1 of rose essential oil |
| Table 1: | The effect of rose essential oil on the number of stimulations to reach specific kindling stages |
 | |
| Each group of animals (n = 8) received daily essential oil and solvent of rose oil (control) and were then stimulated 30 min after injection. *: Significantly different when compared to control group at 0.05 level | |
in control group. The number of stimulations required for the first appearance of 4th and 5th stages of seizure was significantly larger in group treated with 1000 mg kg-1 of rose essential oil than control group (Table 1).
The effect of rose essential oil on the duration of after discharge was significantly different and essential oil could reduce the after discharge duration (Fig. 1). Mean after discharge duration in 2nd, 3rd and 5th stages of seizure was also significantly shorter in the experimental group pretreated with 1000 mg kg-1 of rose essential oil than control group (Table 2).
Mean after discharge amplitude was also shorter in the group treated with 1000 mg kg-1 of rose essential oil than control group but these effects did not show significantly different (Table 3).
| Table 2: | The effect of rose essential oil on the afterdischarge duration at specific kindling stages (unit is second) |
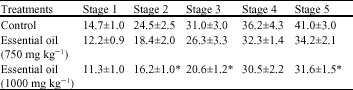 | |
| Each group of animals (n = 8) received daily essential oil and solvent of rose oil (control) and were then stimulated 30 min after injection. *: Significantly different when compared to control group at 0.05 level | |
| Table 3: | The effect of rose essential oil on the after discharge amplitude at specific kindling stages (unit is millivolt) |
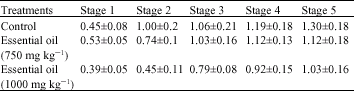 | |
| Each group of animals (n = 8) received daily essential oil and solvent of rose oil (control) and were then stimulated 30 min after injection | |
| Table 4: | The effect of rose essential oil on the duration of 5th stage at fully-kindled rats (unit is second) |
| | |
| Each group of animals (n = 8) received daily essential oil and solvent of rose oil (control) and were then stimulated 30 min after injection. *: Significantly different when compared to control group at 0.05 level | |
Duration of 5th stage of seizure at fully-kindled rats was significantly shorter in two experimental groups than control group (Table 4).
DISCUSSION
Kindling phenomenon is increased neuronal excitability by several mechanisms and led to seizure activity. An enhanced activity of the glutamatergic excitatory neurotransmitter systems and reduction in the activity of the inhibitory GABAergic neurotransmitter systems has also been observed after amygdala electrical kindling (Morimoto et al., 2004; Raisinghani and Faingold, 2005). Electrical kindling also induces increased acetylcholine concentrations in the amygdala and reduced GABA or enhanced cholinergic receptor function in kindled rats (Lin et al., 2002). After kindling, changes are observed in the voltage-dependent inactivation of the sodium current in hippocampal CA1 neurons (Vreugdenhil et al., 1998). In the kindled amygdala, extracellular glutamate is increased by 200-300%, while GABA was reduced by 67% during kindled seizures (Morimoto et al., 2004). Excessive activation of ionotropic glutamate receptors, namely N-methyl-D-aspartate (NMDA) receptors, a-amino-3 hydroxy-5-methyl-4-isoxazolepropionic acid (AMPA) receptors and kainate receptors, is involved in the induction and propagation of seizures and pathological changes after kindling (Yamashita et al., 2004). The studies suggest that inhibitory effects of drugs on development of amygdala kindling might be attributed to action on glutamate release and NMDA-receptor mediated events (Yoshida et al., 2001). The anticonvulsant efficacy believed to be based on the depression of membrane excitability by blocking of sodium, potassium or calcium channels (Ebert et al., 1997; Stratton et al., 2003; Leppik et al., 2006). Kindling model of epilepsy is also a model of synaptic plasticity or Long-Term Potentiation (LTP) and the antiepileptogenic drugs suppress induction of LTP by Inhibition of calcium currents and glutamate release from neurons (Sayyah et al., 2003, 2005). Several drugs have mediated GABAergic neurotransmission by decreasing GABA metabolism, preventing GABA reuptake, increasing GABA synthesis or potency of the GABA receptors (Murao et al., 2002; Leppik et al., 2006; Rogawski, 2006). GABAergic inhibition suppress seizures in most models of epilepsy, including the amygdala-kindling model of temporal lobe epilepsy (Wang et al., 2002; Freichel et al., 2004). Amygdala kindling leads to a significant decrease of GABAergic interneurons in central Piriform Cortex (PC) and elevation of GABA levels in this sub region of the PC is capable of suppressing amygdala kindling (Schwabe et al., 2005).
Table 1 results that essential oil of rose significantly retarded appearance of 1st, 2nd and 3rd stages of seizure in two experimental groups than control group, while appearance of 4th and 5th stages of seizure significantly retarded in group treated with 1000 mg kg-1 of rose essential oil than control group. As in the kindling model of epilepsy, the behavioral seizure stages of 1, 2 and 3 most often originate from foci within the limbic system (including the amygdala) and are considered as focal seizures, while the 4 and 5 stages represent secondary generalized motor seizures (Sayyah et al., 2005). Therefore, it can be conclude that effects of rose essential oil on the kindling is dose-dependent and it retarded focal seizures and generalized seizures acquisition respectively in low and high levels of dose.
Table 2 results that essential oil of rose significantly reduced the increase of after discharge duration in 2nd, 3rd and 5th stages of seizure in the group treated with 1000 mg kg-1 of rose essential oil than control group. Thus, it can be proposed that rose essential oil in high dose with effects on brain structures and neurons of amygdala and limbic system increases the neuronal hyperpolarization and inhibit neuronal synchronization leading to decrease of after discharge duration.
Table 3 can conclude that essential oil of rose decreases after discharge amplitude in two experimental groups but these effects did not show significantly different. As amplitude of waves shown potency of action potential in neurons, on this basis it can be conclude that essential oil of rose non-ability to decreased potency of action potential or severity of seizures.
Table 4 shows that essential oil decreased duration of 5th stage of seizure at fully-kindled rats in both experimental groups. As explained by other researchers, flavonoids are an important component of rosa and as suggested by Dekermendjian et al. (1999), Paladini et al. (1999) and Marder and Paladini (2002), we can propose that such components of Rosa damascena act on the GABAA receptors in the brain of rats. In addition, researchers have shown that flavonoids can enhance the effects of benzodiazepines on GABA receptors that may confirm the anticonvulsive effects of Rosa damascena (Erica et al., 2004). Other component of Rosa damascena essence is geraniol that behavioral studies have shown its hypnotic and anti-epileptic effects (Sayyah et al., 2005). The other component of Rosa damascena is eugenol, which many studies have shown its anti-epileptic effects (Wie et al., 1996).
In conclusion, our study demonstrates that Rosa damascena essential oil retarded the development of behavioral seizures in amygdala electrical kindling and possesses the ability to counteract kindling acquisition.
ACKNOWLEDGMENTS
This research was supported by the grant of the research office of Mashhad University and special helps from the research center for medicinal plants of Mashhad Medical University, Iran.
REFERENCES
- Achuthan, C.R., B.H. Babu and J. Padikkala, 2003. Antioxidant and hepatoprotective effects of Rosa damascena. Pharm. Biol., 41: 357-361.
CrossRefDirect Link - Basim, E. and H. Basim, 2003. Antibacterial activity of Rosa damascena essential oil. Fitoterapia, 74: 394-396.
CrossRefDirect Link - Boskabady, M.H., S. Kiani and H. Rakhshandah, 2006. Relaxant effects of Rosa damascena on guinea pig tracheal chains and its possible mechanism(s). J. Ethnopharmacol., 106: 377-382.
Direct Link - Coulter, D.A., D.C. Mclntyre and W. Loscher, 2002. Animal models of limbic epilepsies: What can they tell us? Brain Pathol., 12: 240-256.
Direct Link - Dekermendjian, K., P. Khanberg, M.R. Witt, O. Sterner, M. Nielsen and T. Liljefors, 1999. Structure-activity relationships and molecular modeling analysis of flavonoids binding to the benzodiazepine site of the rat brain GABAA receptor complex. J. Med. Chem., 42: 4343-4350.
CrossRefDirect Link - Erica, L., Campbel, M.C. Graham and A.R. Johnston, 2004. The dietary flavonoids apigenin and epigallocatechin gallate enhance the positive modulation by diazepam of the activation by GABA of recombinant GABAA receptors. Biochem. Pharm., 68: 1631-1638.
Direct Link - Freichel, C., U. Ebert, H. Potschka and W. Loscher, 2004. Amygdala-kindling does not induce a persistent loss of GABA neurons in the substantia nigra pars reticulata of rats. Brain Res., 1025: 203-209.
CrossRef - Leppik, I.E., K. Kelly, L.D. Toledo-morrell, P.R. Patrylo, R.J. DeLorenzo, G.W. Mathern and H.S. White, 2006. Basic research in epilepsy and aging. Epilepsy Res., 68: 21-37.
Direct Link - Lin, M.T., J.J. Wong and M.S. Young, 2002. The protective effect of dl-tetrahydropalmatine against the development of amygdala kindling seizures in rats. Neurosci. Lett., 320: 113-116.
CrossRef - Mahmood, N., S. Piacente, C. Pizza, A. Burke, A.I. Khan and A.J. Hay, 1996. The anti-HIV activity and mechanisms of action of pure compounds isolated from Rosa damascena. Biochem. Biophys. Res. Commun., 229: 73-79.
CrossRefPubMedDirect Link - Marder, M. and A.C. Paladini, 2002. GABAA receptor ligands of flavonoid structure. Curr. Top. Med. Chem., 2: 853-867.
Direct Link - Morimoto, K., M. Fahnestock and R.J. Racine, 2004. Kindling and status epilepticus models of epilepsy: Rewiring the brain. Prog. Neurobiol., 73: 1-60.
CrossRef - Murao, K., K. Shingu, E. Miyamoto, S. Ikeda, S. Nakao, M. Masuzawa and M. Yamada, 2002. Anticonvulsant effects of sevoflurane on amygdaloid kindling and bicuculline-induced seizures in cats: Comparison with isoflurane and halothane. Anesthesia, 16: 34-43.
CrossRef - Paladini, A.C., M. Marder, H. Viola, C. Wolfman, C. Wasowski and J.H. Medina, 1999. Flavonoids and the central nervous system: From forgotten factors to potent anxiolytic compounds. J. Pharm. Pharmacol., 51: 519-526.
CrossRefDirect Link - Racine, R.J., 1972. Modification of seizure activity by electrical stimulation: II. Motor seizure. Electroencephalography Clin. Neurophysiol., 32: 281-294.
CrossRefDirect Link - Raisinghani, M. and C.L. Faingold, 2005. Neurons in the amygdala play an important role in the neuronal network mediating a clonic form of audiogenic seizures both before and after audiogenic kindling. Brain Res., 1032: 131-140.
CrossRef - Rogawski, M.A., 2006. Molecular targets versus models for new antiepileptic drug discovery. Epilepsy Res., 68: 22-28.
Direct Link - Saberi, M. and M.H. Pourgholami, 2003. Estradiol alters afterdischarge threshold and acquisition of amygdale kindled seizures in male rats. Neurosci. Lett., 340: 41-44.
CrossRef - Sarkisian, M.R., 2001. Overview of the current animal models for human seizure and epileptic disorders. Epilepsy Behav., 2: 201-216.
Direct Link - Sayyah, M., I.T. Najafabadi, S. Beheshti and S. Majzoob, 2003. Lipopolysaccharide retards development of amygdala kindling but does not affect fully-kindled seizures in rats. Epilepsy Res., 57: 175-180.
Direct Link - Sayyah, M., S. Beheshti, M.A. Shokrgozar, A. Eslamifar, Z. Deljoo, A.R. Khabiri and A.H. Rohani, 2005. Antiepileptogenic and anticonvulsant activity of interleukin-1 in amygdala-kindled rats. Exp. Neurol., 191: 145-153.
CrossRef - Schwabe, K., U. Ebert and W. Loscher, 2005. 2005Bilateral microinjections of vigabatrin in the central piriform cortex retard amygdala kindling in rats. Neuroscience, 129: 425-429.
CrossRef - Shafei, M.N., H. Rakhshandah and M.H. Boskabady, 2003. Antitussive effect of Rosa damascena in Guinea pigs. I.J.P., 2: 231-234.
Direct Link - Shea, M.S.O., J.B. Rosen, R.M. Post and S.R.B. Weiss, 2000. Specific amygdaloid nuclei are involved in suppression or propagation of epileptiform activity during transition stage between oral automatisms and generalized clonic seizures. Brain Res., 873: 1-17.
Direct Link - Smedt, T.D., K. Vonck, R. Raedt, S. Dedeurwaerdere, P. Claeys, B. Legros, T. Wyckhuys, W. Wadman and P. Boon, 2005. Rapid kindling in preclinical anti-epileptic drug development: The effect of levetiracetam. Epilepsy Res., 67: 109-116.
Direct Link - Stratton, S.C., C.H. Large, B. Cox, G. Davies and R.M. Hagan, 2003. Effects of lamotrigine and levetiracetam on seizure development in a rat amygdala kindling model. Epilepsy Res., 53: 95-106.
Direct Link - Wie, M.B., M.H. Won, K.H. Lee, J.H. Shin, J.C. Lee, et al., 1996. Eugenol protects neuronal cells from excitotoxic and oxidative injury in primary cortical cultures. Neurosci. Lett., 225: 93-98.
CrossRefDirect Link - Yamashita, H., K. Ohno, H. Inmai, J.I. Shishikura, S. Sakamoto, M. Okada and T. Yamaguchi, 2004. Suppression of fully kindled seizure and retardation of kindling acquisition by YM928 in the rat kindling model of epilepsy. Eur. J. Pharm., 494: 147-154.
Direct Link - Yoshida, M., E. Noguchi, N. Tsuru and N. Ohkoshi, 2001. Effect of riluzole on the acquisition and expression of amygdala kindling. Epilepsy Res., 46: 101-109.
Direct Link