
M. Ramin
Department of Animal Science, Faculty of Agriculture, 43400 UPM Serdang, Selangor, Malaysia
A.R. Alimon
Department of Animal Science, Faculty of Agriculture, 43400 UPM Serdang, Selangor, Malaysia
J.M. Panandam
Department of Animal Science, Faculty of Agriculture, 43400 UPM Serdang, Selangor, Malaysia
K. Sijam
Department of Plant Protection, Faculty of Agriculture, Universiti Putra Malaysia, 43400 UPM Serdang, Selangor, Malaysia
A. Javanmard
Department of Animal Science, Faculty of Agriculture, 43400 UPM Serdang, Selangor, Malaysia
N. Abdullah
Institute of Tropical Agriculture, 43400 UPM Serdang, Selangor, Malaysia
Pakistan Journal of Biological Sciences
Year: 2008 | Volume: 11 | Issue: 4 | Page No.: 583-588







ABSTRACT
The digestion and Volatile Fatty Acid (VFA) production from rice straw and oil palm fronds by cellulolytic bacteria isolated from the termite Coptotermes curvignathus were investigated. The bacteria were Acinetobacter strain Raminalimon, Enterobacter aerogenes strain Razmin C, Enterobacter cloacae strain Razmin B, Bacillus cereus strain Razmin A and Chryseobacterium kwangyangense strain Cb. Acinetobacter strain Raminalimon is an aerobic bacterium, while the other species are facultative anaerobes. There were significant differences (p<0.05) among the bacteria for Dry Matter (DM) lost and acetic acid production from rice straw and Acinetobacter strain Raminalimon showed the highest activity. The facultative bacteria C. kwangyangense strain Cb (cfu mL-1 231x10-6, OD: 0.5), E. cloacae (cfu mL-1 68x10-7, OD: 0.5) and E. aerogenes (cfu mL-1 33x10-7, OD: 0.5) were used for digestion study with the rumen fluid microflora. The in vitro gas production technique was applied for the comparative study and the parameters measured were pH, gas (volume), dry matter lost, acetic acid, propionic acid and butyric acid concentrations. pH was not significantly (p<0.05) different among the five treatments. The bacterium C. kwangyangense strain Cb showed the highest activity (p<0.05) for DM lost, acetic acid, propionic acid and butyric acid production from rice straw when compared to the other bacterial activities. There was no significance (p<0.05) difference between the three bacteria for the dry matter lost of oil palm fronds but the production of Volatile Fatty Acids (VFA) was significantly (p<0.05) high in the treatment which was inoculated with C. kwangyangense strain Cb. The Gen Bank NCBI/EMBL accession numbers for the bacterial strains are EU332791, EU305608, EU305609, EU294508 and EU169201.
PDF Abstract XML References Citation
How to cite this article
M. Ramin, A.R. Alimon, J.M. Panandam, K. Sijam, A. Javanmard and N. Abdullah, 2008. Digestion of Rice Straw and Oil Palm Fronds by Microflora
from Rumen and Termite Bacteria, in vitro. Pakistan Journal of Biological Sciences, 11: 583-588.
DOI: 10.3923/pjbs.2008.583.588
URL: https://scialert.net/abstract/?doi=pjbs.2008.583.588
DOI: 10.3923/pjbs.2008.583.588
URL: https://scialert.net/abstract/?doi=pjbs.2008.583.588
INTRODUCTION
Termites are important in the degradation of plant matter which includes cellulose, hemicellulose, lignin and flavonoids (Harazono et al., 2003). The intestinal tracts of termites comprise of one or several dilated hindgut compartments, which harbor the bulk of the intestinal microbiota and were initially considered as fermentation chambers analogous to the rumen of sheep and cattle (Brune and Friedrich, 2000). The gut microorganisms of termites consist of protozoa, bacteria, spirochetes and fungi. The most important metabolic activities traditionally attributed to the gut microbiota are, first, hydrolysis of cellulose and hemicellulose, second, fermentation of the depolymerization products to short-chain fatty acids, which are then resorbed by the host and third, intestinal nitrogen cycling and dinitrogen fixation (Breznak and Brune, 1994; Brune, 1998). The term cellulase traditionally includes two types of enzymes, exoglucanase that hydrolyses cellulose from its non reducing or reducing ends and endo-ß-1,4-glucanase that hydrolyses along the glucan chain of cellulose fibres randomly. In termites, it has been proposed that endo-ß-1, 4-glucanase plays a primary role in cellulose digestion (Konig, 2005). Carboxymethylcellulose (CMC), which measures endo-ß-1, 4-glucanase activity is one of the most popular artificial substrates for measuring cellulose activity because of its high solubility in water. Thus carboxymethyl-cellulose has been preferentially used in most studies of cellulose digestion in termites (Tokuda et al., 2005). An in vitro gas production technique has been recently used to study activities of rumen microflora on various substrates. A feedstuff is incubated with buffered rumen fluid and the gases as well as the end product of fermentation are indicators of fermentation kinetics (Rymer et al., 2005). Agriculture by-products such as Oil Palm Frond (OPF) and others have been shown to be available roughage feeds for ruminant animals (Khamseekhiew et al., 2002), however, their high fiber contents are the main factors limiting their digestibility and intake by ruminants. The objective of this study was to conduct an in vitro study to measure the effects of the cellulolytic bacteria isolated from the termite gut on the digestion of a fiber substrate and to compare the in vitro digestibility of rice straw and oil palm fronds by rumen microbial flora and termite bacteria.
MATERIALS AND METHODS
Isolates were prepared in the Microbiology Laboratory of Universiti Putra Malaysia (UPM) and after preparation they were brought to the Animal Nutrition Laboratory, UPM for in vitro studies.
Termite bacteria: Five bacterial species were previously isolated from the gut of the termite Coptotermes curvignathus. The bacterial species were identified as Acinetobacter strain Raminalimon (EU332791), Enterobacter aerogenes strain Razmin C (EU305608), Bacillus cereus Strain Razmin A (EU294508), Enterobacter cloacae strain Razmin B (EU305609) and Chryseobacterium kwangyangense strain Cb (EU169201).
In vitro digestion of rice straw by bacterial species from the termite gut: Each bacterial species was inoculated in 10 mL Nutrient Broth (NB) and the Optical Density (OD) was adjusted to 0.5. Then it was mixed with 20 mL buffer medium (Getachew et al., 2005) the total volume was 30 mL. Approximately 200 mg rice straw was ground to a maximum particle size of 1 mm and were added to each syringe. The cultures in triplicate were incubated at 39°C for 24 h. T1 with nutrient broth without bacteria (Control), T2 with Acinetobacter, T3 with Enterobacter aerogenes, T4 with Enterobacter cloacae, T5 with Bacillus cereus and T6 with Chryseobacterium Kwangyangense strain Cb and T7 was a mix culture of all the bacteria.
In vitro true dry matter digestibility (IVTDMD): All conditions were same as described before with some differences; a blank treatment was added without any substrates (rice straw or oil palm frond) for subtraction with treatments for the appearance of the particles
| Table 1: | Experimental design showing treatments |
 | |
| Thirty five milliliter buffered rumen fluid was mixed with each treatment (5 mL), Blank: Thirty five milliliter buffered rumen + 5 mL nutrient broth, NB: Nutrient Broth | |
inside the rumen fluid. The amount of substrates was approximately 500 mg. The buffer was flushed with CO2 to remove the oxygen in the medium and the total medium in each syringe was 40 mL. The substrates were oven dried for overnight at 60°C, ground through 1 mm screen sieve and stored for the in vitro studies. Two in vitro studies were performed, one for rice straw and another for oil palm frond.
Rumen fluid collection: Rumen fluid was collected before morning feeding from Kedah-Kelantan (KK) cattle of about 4 years of age with an average body weight of ±300 kg, fitted with a ruminal cannulae. The rumen content was collected into 1 L capacity pre-warmed water flask. The cattle were fed twice per day during the experiment, once at 09:00 h and again at 16:30 h. Drinking water was freely available to the animal throughout the experiment.
Treatments: The treatment for blank consisted of 30 mL buffer (Getachew et al., 2005), 5 mL rumen fluid and 5 mL Nutrient Broth (NB) without any substrate. Five treatments consisted of 30 mL buffer, 5 mL rumen fluid, NB and distilled water and were inoculated with 5 mL of one of the bacteria. Approximately 500 mg rice straw was added to each syringe. The last treatment was as the blank except that it had distilled water instead of NB (Table 1).
Dry Matter (DM) lost, VFA production and gas production (after 24 h) was calculated for blank and then subtracted from all treatments.
True dry matter digestibility (TDMD): After 24 h of incubation the in vitro true dry matter digestibility was determined by refluxing the residue of each treatment into 600 mL beakers with 70 mL Neutral-Detergent Solution (NDS) for 1 h (Blummel et al., 1997). True digestibility was calculated as the weight of substrate incubated minus the weight of the residue after NDS treatment. The dry matter lost of each treatment was subtracted with the blank.
VFA production: Volatile Fatty Acids from each treatment were analyzed by the method of Minato and Kudo referred to by Jetana et al. (1996).
Gas production and pH determination: Volumes of the gas produced were read after 24 h of incubation. The total amount of the gas of each treatment was subtracted with the blank. The pH of each treatment immediately after incubation was determined using pH meter.
Statistical analyses: The mean of the parameter measured: pH, dry matter lost, VFAs and gas production, were analyzed by analysis of variance (ANOVA) using the Statistical Analysis System Institute software (SAS, 1997). The differences between the treatment means were tested using the Least Significant Difference (LSD) method.
RESULTS AND DISCUSSION
More details about some of the bacterial isolates are given in Ramin et al. (2008). All data presented in Table 2 should be subtracted with the blank; the amount of dry matter lost in the blank relates to insoluble fraction of rice straw. There were significant (p<0.05) differences among the treatments. The amount of Dry Matter (DM) lost and acetic acid concentrations were higher for the treatments with bacterial isolates, including Acinetobacter (Table 2). Acinetobacter is an aerobic bacterium. There was no significant difference between Enterobacter aerogenes, Enterobacter cloacae and Chryseobacterium kwangyangense strain Cb for dry matter lost and concentration of acetic acid. The concentrations of the other volatile fatty acids were not so high. The mixed culture did not significantly show a high effect on rice straw digestion. For all treatments the production of gas was not so high. The second and third in vitro studies were conducted to determine the best bacteria from termites with rumen microflora for rice straw and oil palm fronds digestion. For in vitro studies with the aid of rumen microflora an anaerobic condition is necessary so the facultative bacteria with the highest effect from the previous experiment were selected; the bacteria were: E. aerogenes, (cfu mL-1 33x10-7, OD: 0.5) E. cloacae (cfu mL-1 68x10-7, OD: 0.5) and C. kwangyangense strain Cb (cfu mL-1 231x10-6, OD: 0.5).
The activity of the bacterial inoculate in each treatment with rumen fluid was determined. The bacterium Chryseobacterium kwangyangense strain Cb significantly (p<0.05) showed the best effect for DM lost, acetic acid, propionic acid and butyric acid concentrations when compared to the other treatments (Table 3).
In all treatments, the pH did not differ significantly except for the treatment inoculated with Enterobacter aerogenes, which had a negligible difference. The amount of gas production did not differ significantly (p<0.05)
| Table 2: | Mean of the dry matter lost and acetic acid concentrations of termite gut bacteria |
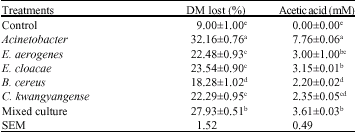 | |
| a,b,c: Means in the same column with different subscripts are different significantly (p<0.05), Each treatment had three replicates, Substrate: Rice straw, Control: Nutrient broth | |
among the treatments, of which the gas production was lowest for the treatment with distilled water. The concentration of volatile fatty acids was higher (p<0.05) in the treatment inoculated with Chryseobacterium kwangyangense strain Cb. In order to compare the effect of the bacteria on another fiber content substrate, Oil Palm Fronds (OFP) were selected as a substrate due to their high fiber contents. There was no significance difference between the three bacterial species when compared for the digestion of oil palm fronds (Table 4), but there were significance (p<0.05) differences when the bacterial treatments were compared with Nutrient Broth (NB) and distilled water. In the latter case the three bacterial species showed the higher activity on dry matter lost. There was no significance difference for pH between the treatments. However, the high pH recorded seems to suggest that the condition were conducive to cellulolytic bacteria fermentation; it could also be due to the low VFA production from the highly indigestible OPF-based substrates (Khamseekhiew et al., 2002). The production of gas was lowest for the treatment with distilled water and it was significantly (p<0.05) higher for the treatments which were inoculated with C. kwangyangense and E. cloacae (Table 4). The production of Volatile Fatty Acids especially acetic acid was significantly higher for the treatment which included C. kwangyangense strain Cb (Table 4).
The ability of Termite bacteria to digest lignocellulose compounds has been published. Acinetobacters, Bacillus cereus and Enterobacters have been reported to be able to degrade 34-62% cellulose and 14-32% hemicellulose and in addition Enterobacters are also able to degrade 18-39% lignin (Konig, 2005), Acinetobacters are classified under the aerobic group with the capability to degrade caffeic acid and syringic acid (lignin monomers) and Enterobacters are grouped among bacteria with the capability of modifying lignin monomers and other aromatic compounds (Kuhnigk et al., 1994; Varma et al., 1994). Enterobacters are also able to assimilate different phenolic compounds considered as
| Table 3: | Mean of the dry matter lost, pH, gas production, acetic acid, propionic acid and butyric acid concentrations of the treatments |
 | |
| a, b, c: Means in the same column with different subscripts are different significantly (p<0.05), Three replicates for each treatment, Bac A: E. aerogenes, Bac B: E. cloacae, Bac C: C. Kwangyangense, NB: Nutrient broth, Substrate: Rice straw | |
| Table 4: | Mean of the dry matter lost, pH, gas production, acetic acid, propionic and butyric acid production of the treatments |
 | |
| a, b, c: Means in the same column with different subscripts are different significantly (p<0.05), Three replicates for each treatment, Bac A: E. aerogenes, Bac B: E. cloacae, Bac C: C. kwangyangense, NB: Nutrient broth, Substrate: Oil palm fronds | |
lignin related simple monomers (Deschamps et al., 1980). Borji et al. (2003) has isolated and identified some bacteria capable of degrading straw lignin and polysaccharides and they were identified as Bacillus sp., Enterobacter sp. and Ocrobacterium. The bacterium from the Enterobacters family had a faster growth rate than the other two species. Schafer et al. (1996) has identified some bacteria from Enterobacteriaceae and Acinetobacters family with the ability to degrade hemicellulose compounds. The first investigations for isolation of facultative anaerobes come from Adams and Boopathy (2005), who in their study identified some facultative anaerobes as Enterobacter families from the termite Coptotermes formosanus. Purwadaria et al. (2003) have investigated the fiber hydrolytic enzyme activity from termite gut extract as poultry feed applications and they demonstrated that the extract which comes from the microorganisms inside termite gut were more appropriate in digesting feedstuffs with high lignocellulose (fiber) such as rice bran. Volatile Fatty Acids (VFA) are the chief end-products of cellulose digestion in the rumen, in the hindgut of termites and acetic acid is the most common volatile fatty acid produced (Mcfarlane and Alli, 1985). Odelson and Breznak (1983) have also demonstrated that acetate dominated the extracellular pool of volatile fatty acids in the hindgut fluid of Reticulitermes flavipes, Zootermopsis angusticollis and Incisitermes schwarzi, where it occurred at concentrations of 57.9 to 80.9 mM. The production of acetate is also reported by Droge et al. (2006) from Spirochaeta coccoides sp. nov from the hindgut of the termite Neotermes castaneus. There are few reports on the isolation and identification of Chryseobacterium species from insect gut specially termites. Organisms tentatively described as Flavobacterium had only once been isolated from insect intestinal tracts which were described by Eutick et al. (1978) and the cellulolytic activity was not reported. Dugas et al. (2001) is the second person who had identified a Chryseobacterium species from the gut of the American cockroach fed with high fiber content diet (30% crude fiber by weight). Some species of this bacterium are able to degrade feather as described by Brandelli and Riffel (2005). David Landry has patented (US20020072104A1) a pure glycosidase enzyme which was obtained from the genus Chryseobacterium. These days feed additives or Direct Fed Microbial (DFM) are among the most important supplements for ruminants (Yang et al., 2004), examples are Saccharomyces cerevisiae (Lynch and Martin, 2002) and Enterococcus faecium (Ghorbani et al., 2002). It can be concluded that all bacteria were able to digest rice straw and produce Volatile Fatty Acids (VFA) especially acetate. Termite bacteria showed an improvement on rice straw and oil palm frond digestion and production of VFA which proves their ability to digest fiber materials. The bacterium Chryseobacterium kwangyangense strain Cb showed the best effect on the dry matter lost, concentration of acetic acid when compared to the other bacterial species used in the treatments. Further studies need to be conducted to determine the potential of microorganisms, especially bacteria from termite gut, for industrial applications such as feed additives for poultry or ruminants diet.
ACKNOWLEDGMENTS
We thank Mr. Mohammad Zawawi and Mr. Ali Nargeskhani for their technical supports in this study.
REFERENCES
- Adams, L. and R. Boopathy, 2005. Isolation and characterization of enteric bacteria from the hindgut of Formosan termite. J. Bioresour. Technol., 96: 1592-1598.
Direct Link - Blummel, M., H. Steingaβ and K. Becker, 1997. The relationship between in vitro gas production, in vitro microbial biomass yield and 15N incorporation and its implications for the prediction of voluntary feed intake of roughages. Br. J. Nutr., 77: 911-921.
CrossRefDirect Link - Borji, M., S. Rahimi, G. Ghorbani, J. Vand Yoosefi and H. Fazaeli, 2003. Isolation and identification of some bacteria from termites gut capable in degrading straw lignin and polysaccharides. J. Fac. Vet. Med. Univ. Tehran, 58: 249-256.
Direct Link - Brandelli, A. and A. Riffel, 2005. Production of an extracellular from Chryseobacterium sp. growing on raw feathers. Elect. J. Biotech., 8: 35-42.
Direct Link - Breznak, J.A. and A. Brune, 1994. Role of microorganisms in the digestion of lignocellulose by termites. Annu. Rev. Entomol., 39: 453-487.
CrossRef - Brune, A. and M. Friedrich, 2000. Microecology of the termite gut: Structure and function on a microscale. Curr. Opin. Microbiol., 3: 263-269.
Direct Link - Droge, S., J. Frohlich, R. Radek and H. Konig, 2006. Spirochaeta coccoides sp. nov., a novel coccoid spirochete from the hindgut of the termite Neotermes castaneus. J. Applied Environ. Microbiol., 72: 392-397.
Direct Link - Dugas, J.E., L. Zurek, B.J. Paster and B.A. Keddie, 2001. Isolation and characterization of a Chryseobacterium strain from the gut of the American cockroach, Periplaneta americana. Arch. Microbiol., 175: 259-262.
Direct Link - Getachew, G., E.J. DePeters, P.H. Robinson and J.G. Fadel, 2005. Use of an in vitro rumen gas production technique to evaluate microbial fermentation of ruminant feeds and its impact on fermentation products. J. Anim. Feed. Sci. Technol., 123-124: 547-559.
CrossRefDirect Link - Ghorbani, G.R., D.P. Morgavi, K.A. Beauchemin and J.A.Z. Leedle, 2002. Effects of bacterial direct-fed microbials on ruminal fermentation, blood variables and the microbial populations of feedlot cattle. J. Anim. Sci., 80: 1977-1985.
PubMedDirect Link - Harazono, K., N. Yamashita, N. Shinzato, H. Watanabe, T. Fukatsu and R. Kurane, 2003. Isolation and characterization of aromatics-degrading microorganisms from the gut of the lower Coptotermes formosanus. J. Biosci. Biotechnol. Biochem., 67: 889-892.
Direct Link - Konig, H., 2006. Bacillus species in the intestine of termites and other soil invertebrates. J. Applied Microbiol., 101: 620-627.
Direct Link - Kuhnigk, T., E.M. Borst, A. Ritter, P. Kampfer, A. Graf, H. Hertel and H. Konig, 1994. Degradation of lignin monomers by the hindgut flora of xylophagous termites. J. Syst. Applied Microbiol., 17: 76-85.
CrossRefDirect Link - Lynch, H.A. and S.A. Martin, 2002. Effects of Saccharomyces cerevisiae culture and Saccharomyces cerevisiae live cells on in vitro mixed ruminal microorganism fermentation. J. Dairy Sci., 85: 2603-2608.
CrossRefDirect Link - Purwadaria, T., P.P. Ketaren, A.P. Sinurat and I. Sutikno, 2003. Identification and evaluation of fiber hydrolytic enzymes in the extract of termites (Glyptotermes montanus) for poultry feed application. Indon. J. Agric. Sci., 4: 40-47.
Direct Link - Ramin, M., A.R. Alimon, N. Abdullah, J.M. Panandam and K. Sijam, 2008. Isolation and identification of three species of bacteria from the termite Coptotermes curvignathus (Holmgren) present in the Vicinity of University Putra Malaysia. Res. J. Microbiol., 3: 288-292.
CrossRefDirect Link - Rymer, C., J.A. Huntington, B.A. Williams and D.I. Givens, 2005. In vitro cumulative gas production techniques: History, methodological considerations and challenges. J. Anim. Feed Sci. Technol 123-124: 9-30.
CrossRefDirect Link - Tokuda, G., L. Nathan and H. Watanabe, 2005. Marked variations in patterns of cellulase activity against crystalline-vs. carboxymethyl-cellulose in the digestive systems of diverse, wood-feeding termites. J. Physiol. Entomol., 30: 372-380.
Direct Link - Varma, A., B. Krishna Kolli, J. Paul, S. Saxena and H. Konig, 1994. Lignocellulose degradation by microorganisms from termite hills and termite guts: A survey on the present state of art. FEMS. Microbiol. Rev., 15: 9-28.
CrossRef - Yang, W.Z., K.A. Beauchemin, D.D. Vedres, G.R. Ghorbani, D. Colombatto and D.P. Morgavi, 2004. Effects of direct-fed microbial supplementation on ruminal acidosis, digestibility and bacterial protein synthesis in continuous culture. Anim. Feed Sci. Technol., 114: 179-193.
CrossRefDirect Link - Schafer, A., R. Konrad, T. Kuhnigk, P. Kampfer, H. Hertel and H. Konig, 1996. Hemicellulose-degradating bacteria and yeasts from the termite gut. J. Applied Microbiol., 80: 471-478.
CrossRefDirect Link