
Abbas Mohammadi
Department of Plant Pathology, Tarbiat Modares University, Tehran, Iran
Azizollah Alizadeh
Department of Plant Pathology, Tarbiat Modares University, Tehran, Iran
Mansore Mirabolfathey
Plant Pests and Disease Research Institute, P.O. Box 19395, Tehran, Iran
Nasrin Nooras Mofrad
College of Agriculture, Islamic Azad University of Birjand, Iran
Pakistan Journal of Biological Sciences
Year: 2008 | Volume: 11 | Issue: 2 | Page No.: 302-305







ABSTRACT
Phytophthora root and stem rot of soybean is a destructive disease of soybean in Iran. Races 1 and 3 of pathogen have already been reported from two major growing regions of the crop, Lorestan and Golestan provinces. In a survey during 2004-2005, 142 isolates of P. sojae were recovered from infected plants and naturally infested soil samples using selective media and soybean leaf baiting technique. The majority of tested isolates (110 isolates) belonged to race one of P. sojae and 32 isolates belonged to race 3. ITS region of 23 isolates were amplified with specific primers Ps1 and Ps2. Sequences of this regions were similar to other gene banks sequences except two isolates from China. This survey showed low diversity in Iranian population of P. sojae.
PDF Abstract XML References Citation
How to cite this article
Abbas Mohammadi, Azizollah Alizadeh, Mansore Mirabolfathey and Nasrin Nooras Mofrad, 2008. Races of Phytophthora sojae in Iran. Pakistan Journal of Biological Sciences, 11: 302-305.
DOI: 10.3923/pjbs.2008.302.305
URL: https://scialert.net/abstract/?doi=pjbs.2008.302.305
DOI: 10.3923/pjbs.2008.302.305
URL: https://scialert.net/abstract/?doi=pjbs.2008.302.305
INTRODUCTION
Phytophthora root and stem rot of soybean (Glycine max L. Merr.) caused by Phytophthora sojae, is widespread throughout soybean growing areas of the world (Doupnik, 1993). This species, formerly named Phytophthora megasperma Drechs. f. sp. glycinea (Kuan and Erwin, 1980) exhibits aggressive, race-specific pathogenicity to soybean and causes few or no symptoms on other hosts (Hansen and Maxwell, 1991). The population of this pathogen is made up of numerous pathogenic or physiological races describe by their virulence on a set of differential soybean varieties (Keeling, 1982; Layton and Kuhn, 1988; Schmitthenner et al., 1994).
Since P. sojae was recorded in the United States in 1955 (Suhovecky and Schmitthenner, 1955), 50 races have been classified on differential soybean genotypes (Abney et al., 1997) and many more isolates with unique virulence patterns have been identified but not given a race number (Schmitthenner et al., 1994).
Control of this disease has been achieved largely through breeding resistant cultivars (Athow et al., 1980; Schmitthenner, 1985). At present 13 resistance genes (Rps gene) have been characterized in soybean (Burnham et al., 2003). However, P. sojae has been found to overcome these resistance genes by generating new races (Keeling, 1984).
Phytophthora root rot of soybean was first recorded in Iran (Rezaee and Alizadeh, 1998) and quickly bean an important problem in soybean production areas. Previous studies (Sadeghi and Mirabolfathy, 2002) on 22 isolates demonstrate the presence of at least two physiologic races, race 1 and race three, of P. sojae. This paper reports the presence of races among 142 isolates of P. sojae isolated from soil and diseased soybean plants from Iran during 2005-2006 growing season.
MATERIALS AND METHODS
Isolation: Between 2004 and 2005, commercial soybean crops in soybean-growing areas of Iran (Lorestan, Golestan, Mazandaran and Ardabil provinces) were surveyed for Phytophthora root and stem rot. Isolation of P. sojae from plants was done by CMA-PARPH medium (Schmitthenner et al., 1994).
The pathogen was isolated from soil by a modification of the soybean leaf baiting technique (Schmitthenner et al., 1994).
Identification: Keys that groups species of Phytophthora by sporangial characteristics and other characteristics were used in the identification of Phytophthora isolates (Waterhouse, 1963).
Race identification: Inoculums for race identification was prepared by growing isolates on OA (Oatmeal Agar) in Petri plates at 25°C for 2 to 4 weeks. Inoculations were performed by the standard hypocotyls method (Laviolette and Athow, 1981). Ten 7-day-old seedling of soybean differentials (Anderson and Buzzell, 1992; Schmitthenner et al., 1994) representing different Rps alleles were inoculated with each isolates and grown in the greenhouse at 25±2°C with supplemental fluorescent and incandescent light. Race determination was repeated twice with each isolate. Soybean seedling hypocotyls reaction was classified resistant where 70% or more of the seedling alive or susceptible where 70% or more of the seedling killed (Schmitthenner et al., 1994; Barreto et al., 1995).
ITS region sequencing: Amplification and sequencing of the ITS region of the genome was performed with specific primers Ps1 and Ps2 (Wang et al., 2006). Each reaction included 30 ng of template DNA in a volume of 25 μL containing 1.5 mM MgCl2, 0.05 mM each dCTP, dATP, dTTP and dGTP, 200 pM of each primer and 1.5 units of Taq polymerase (Fermentase Inc. Co.). The DNA was amplified under the following conditions: 4 min at 94°C; 35 cycles of 30 sec at 94°C, 30 sec at 55°C and 60 sec at 70°C and final elongation at 72°C for 10 min. For analysis, PCR products were electrophoretically separated in 1.5% agarose containing ethidium bromide at 50 μg L-1 and band sizes were estimated by comparison with 1 kb Plus DNA Ladder molecular weight markers (Wang et al., 2006).
Sequence editing and phylogenetic tree construction: Resulting sequences were aligned and edited using BioEdit software (version 4.7.3; T. Hall, North Carolina State University). Phylogenetic trees based on ITS sequences were compiled using PHYLIP programs. The ITS sequences of other P. sojae isolates were obtained from GenBank. Distance matrices were calculated with DNADIST and used to construct trees by neighbor joining as implemented in NEIGHBOR. The phylogenetic trees were displayed using TreeView (Wang et al., 2006).
RESULTS AND DISCUSSION
From plants and soils samples, 142 isolates were obtained and identified as P. sojae based on morphological characteristics (Fig. 1). The 110 isolates were virulent on harosoy which carries the Rps7 gene therefore they were determined as race 1. Other 32 isolates were virulent on Harosoy (Rps7) and Union (Rps1-a), therefore they were determined as race 3 (Fig. 2).
Apparently, the composition of P. sojae races in Iran did not change during the years of this study. Race 1 and race 3 were the only two isolated in all areas of study and race 1 was dominant. Races 1 and 3 of pathogen have already been reported from two major growing region of the crop, Lorestan and Golestan provinces (Sadeghi and Mirabolfathy, 2002). Races 1 and 3 were reported first time
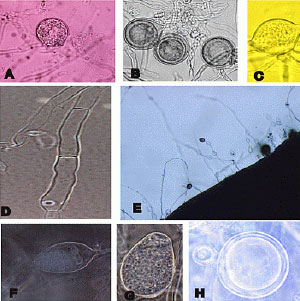 | |
| Fig 1: | Morphological characteristic of Phytophthora sojae (A, B, H: oospore, C: hyphal swelling, D: hyphae, E, F, G: sporangium) |
 | |
| Fig 2: | Race identification test reaction (A: differentials before testing, B: hypocotyls inoculation, C, D: race 3 and 1 reaction (U: union, C: control, H: Harosoy) |
from Moghan province, another major soybean growing area of the country.
PCR with primers Ps1 and Ps2 amplified fragment 328 bp for the Phytophthora sojae isolates but no PCR product was detected from other Phytophthora species (Fig. 3). This is the first report about the detection of P. sojae by specific primers in Iran.
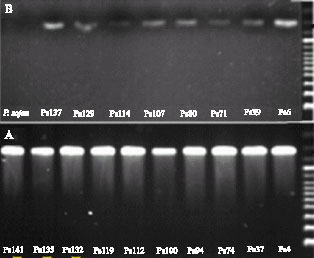 | |
| Fig 3: | 328 bp fragment amplified by specific primers |
Wang et al. (2006) developed a rapid and accurate method for the specific detection of P. sojae in soil. They amplified the Internal Transcribed Spacer (ITS) regions of eight P. sojae isolates using Polymerase Chain Reaction (PCR) with the universal primers DC6 and ITS4, aligned the sequences of PCR products with published sequences of 50 other Phytophthora species and used a region specific to P. sojae to design the specific PCR primers, PS1 and PS2. They used more than 245 isolates representing 25 species of Phytophthora and at least 35 other species of pathogens to test the specificity of the primers. PCR amplification with PS primers resulted in the amplification of a product of approximately 330 bp, exclusively from isolates of P. sojae. Tests with P. sojae genomic DNA determined that the sensitivity of the PS primer set is approximately one fg. This PCR assay allowed the detection of P. sojae from soil within 6 h, with a detection sensitivity of two oospores in 20 g of soil. PCR with the PS primers can detect P. sojae from diseased soybean tissue and residues. The PS primer-based PCR assay provides a rapid and sensitive tool for the detection of P. sojae in soil and infected soybean tissue.
Sequences alignment of ITS regions that amplified by Ps1 and Ps2 primers showed no polymorphism between Iranian isolates of P. sojae. Sequences of Iranian isolates were similar to gene banks sequences except two isolates from china, AY245092 and Ay277278. Phylogenetic analysis put most of isolates in one group and two Chinian isolates in separate groups (Fig. 4).
Sequences alignment of ITS regions that amplified by Ps1 and Ps2 primers showed no polymorphism between Iranian isolates of P. sojae. Sequences of Iranian isolates were similar to gene banks sequences except two isolates from china, AY245092 and Ay277278. Phylogenetic
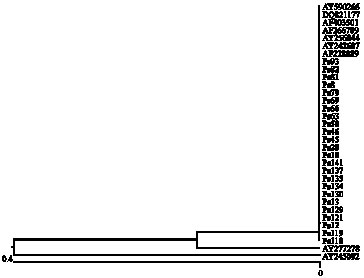 | |
| Fig 4: | Phylogenetic tree based sequences of ITS sequences |
analysis put most of isolates in one group and two Chinian isolates in separate groups (Fig. 4).
Races of P. sojae isolates from United State (Schmitthenner et al., 1994) or Australian (Ryley et al., 1998) had more diversity than the Iranian isolates. In United States 54 races of P. sojae had been found so far (Schmitthenner et al., 1994).
The frequency of occurrence of P. sojae races in Iran is similar to some region of Iran as well as other countries (Yang et al., 1996; White et al., 1983; Tachibana et al., 1975)
Current investigations showed a very low diversity of P. sojae population in Iran and is consistent with earlier studies (Sadeghi and Mirabolfathy, 2002). This is due to restriction in new variety import, absence of other hosts of pathogen, area under culture and absence of Rps gene in commercial varieties in Iran. Soybean cultivars/var. `Williams` and `Clark` are cultured in Lorestan and Moghan respectively, which are susceptible to all or both (please check) races of P. sojae (Alizadeh and Rezaee, 1999) and did not have any selection pressure to appearance of new races and race 1 remain as dominant race in Iranian soybean farms.
Introducing one or two resistance gene-to-gene pool of Iranian commercial variety can reduce disease severity. Some Rps genes are link together and (May et al., 2002) can transfer several genes together to new lines during gene transformation that caused resistance to several races of pathogen.
REFERENCES
- Abney, T.S., J.C. Melgar, T.L. Richards, D.H. Scott, J. Grogan and J. Young, 1997. New races of Phytophthora sojae with Rps1-d virulence. Plant Dis., 81: 653-655.
CrossRef - Anderson, T.R. and R.I. Buzzell, 1992. Inheritance and linkage of the Rps7 gene for resistance to Phytophthora rot of soybean. Plant Dis., 76: 958-959.
Direct Link - Athow, K.L., F.A. Laviolette, E.H. Mueller and J.R. Wilkox, 1980. A new major gene for resistance to Phytophthora megasperma var. sojae in soybean. Phytopathology, 70: 977-980.
Direct Link - Barreto, D., B. Stegman De Gurfimkel and C. Fortugno, 1995. Races of Phytophthora sojae in Argentina and reaction of soybean cultivars. Plant Dis., 79: 599-600.
Direct Link - Burnham, K.D., A.E. Dorrance, D.M. Francis, R.J. Fiorrito and S.K. Martin, 2003. Rps 8, a new locus in soybean for resistance to Phytophthora sojae. Crop Sci., 43: 101-105.
Direct Link - Hansen, E.M. and D.P. Maxwell, 1991. Species of the Phytophthora megasperma complex. Mycologia, 83: 376-381.
Direct Link - Kuan, T.L. and D.C. Erwin, 1980. Form specials differentiation of Phytophthora megasperma isolated from soybean and alfalfa. Phytopathology, 70: 333-338.
Direct Link - Layton, A.C. and D.N. Kuhn, 1988. The virulence of interracial heterokaryons of Phytophthora megasperma f. sp. glycinea. Phytopathology, 78: 961-966.
Direct Link - May, K.J., S.C. Whisson, R.S. Zwart, I.R. Searle and J.A.G. Irwin, 2002. Inheritance and mapping of 11 avirulence genes in Phytophthora sojae. Fungal Genet. Biol., 37: 1-12.
Direct Link - Ryley, M.J., N.R. Obst, J.A.G. Irwin and A. Drenth, 1998. Changes in the racial composition of Phytophthora sojae in Australia between 1979 and 1996. Plant Dis., 82: 1048-1054.
CrossRef - Schmitthenner, A.F., M. Hobe and R.G. Bhat, 1994. Phytophthora sojae races in Ohio over a 10 year interval. Plant Dis., 78: 269-276.
Direct Link - Wang, Y., W. Zhang, Y. Wang and X. Zheng, 2006. Rapid and sensitive detection of Phytophthora sojae in soil and infected soybeans by species-specific polymerase chain reaction assay. Phytopathology, 96: 1315-1321.
Direct Link - White, D.M., J.E. Partridge and J.H. Williams, 1983. Races of Phytophthora megasperma f. sp. glycinea in Eastern Nebraska. Plant Dis., 67: 1281-1282.
Direct Link - Yang, X.B., R.L. Ruff, X.Q. Meng and F. Workneh, 1996. Races of Phytophthora sojae in Iowa soybean fields. Plant Dis., 80: 1418-1420.
Direct Link