
M. Keshavarzi
Department of Biology, Faculty of Science, Alzahra University, Vanak, Tehran, Iran
R. Jahandideh
Department of Biology, Faculty of Science, Alzahra University, Vanak, Tehran, Iran
Z. Nazem Bokaee
Department of Biology, Faculty of Science, Alzahra University, Vanak, Tehran, Iran
Pakistan Journal of Biological Sciences
Year: 2008 | Volume: 11 | Issue: 23 | Page No.: 2599-2605







ABSTRACT
In this study 39 macro-morphological features of 8 populations of Z. clinopodioides Lam. were studied to find their diagnostic features for distinguishing their different subspecies in Iran. Micro-morphological characters of epidermis and pollen grains were considered. In order to determine the most variable morphological characters among the species studied, factor analysis based on principal components analysis was performed using SPSS ver. 9. Palynological studies show the general form of tribe Mentheae. According to the present results, the pollen morphology in Ziziphora clinopodioides may be helpful in elucidating the infra specific relationships. Leaf micro-characters of epidermis were shown to be of great importance in the taxonomy of the Z. clinopodioides subspecies. Using complement macro-characters for taxonomic purposes is highly recommended in Ziziphora L.
PDF Abstract XML References Citation
How to cite this article
M. Keshavarzi, R. Jahandideh and Z. Nazem Bokaee, 2008. Morphological and Anatomical Studies on Ziziphora clinopodioides Lam. (Labiatae). Pakistan Journal of Biological Sciences, 11: 2599-2605.
DOI: 10.3923/pjbs.2008.2599.2605
URL: https://scialert.net/abstract/?doi=pjbs.2008.2599.2605
DOI: 10.3923/pjbs.2008.2599.2605
URL: https://scialert.net/abstract/?doi=pjbs.2008.2599.2605
INTRODUCTION
Ziziphora L. comprises annual and perennial herbs in Iran. These herbaceous or sub shrubby plants have verticil lasters which are scattered in leaf axils or crowded in a terminal capitulum. This genus by about 25 to 30 species in Africa, Asia and Europe has only 4 species in Iran. The only perennial Ziziphora in Iran is Z. clinopodioides Lam. This species contains some subspecies according to Flora iranica (Rechinger, 1982). These subspecies are mainly distinguished from each other by means of calyx characters.
Lamiaceae family has been used for centuries as flavoring agent or spice in different foods and also in traditional medicines for treatment of digestive and viral diseases. Utilization of Ziziphora clinopodioides in yoghurt in many parts of Iran is an example of its daily medicinal usage. This species is used with yogurt, salad and many dishes. Some research has considered this species as a medicinal herb in Iran. Khodaparast et al. (2007) studied the interaction between these compounds and yoghurt to evaluate the effect of Ziziphora clinopodioides on growth of yoghurt starter culture.
Ziziphora in Iran is represented by four species. These are Z. capitata L., Z. clinopodioides Lam, Z. persica Bunge. and Z. tenuior L. All these species are aromatic and montane taxa of Iran. From these taxa Z. clinopodioides is distributed in Caucasia, North of Iraq, Iran, Afghanistan and Central Asia. These species are often a mat-forming perennial with prostrate to erect stems mainly much-branched at base. Their leaves show a high variation in size and shape. In Iran it is a plant of rocky slopes and scree. Subdivision of this polymorphic, basically Irano-Turanian, species into formal infra-specific taxa does not seem to be practical, since the variation of different characters is far from homologous and is poorly correlated with variation within the species (Sensu lato). Various morphological groups of this species in Turkey have also been recognized on the basis of their habit, leaf-shape and indumentums according to Hedge and Lamond (1982).
MATERIALS AND METHODS
Populations of Z. clinipodioides were collected from 8 different localities of Iran from 2007 till 2008 (Table 1). In each sampling site 10 individuals were gathered. Macro and micro morphological characters were studied (Table 2, 3). In Macro-morphological studies 43 features were studied from vegetative and reproductive parts of individuals. Micro-morphological studies considered epidermis and pollen grains. Dorsal epidermis was studied by tissue removal using methylene green.
Pollen for examination by LM was prepared according to the Woodehouse method (Woodehouse, 1935). Pollen grains of the 4 recognized subspecies taxa of Zizizohora clinopodioides were mostly taken from herbarium material deposited at the herbarium of Alzahra University (Tehran, Iran). The polar axis (P), equatorial axis (E), corpus length (Clg) and width (Clt), apocolpium diameter and mesocolpium, exine and intine thickness were measured on the pollen grains. Measurements and light micrographs were taken with an Olympus DP12 microscope. The measured pollen diameters were based on at least 20 samples and other characters on approximately 10 samples under the light microscope. For SEM, pollen grains were transferred directly to a stub and micrographs were obtained using an XL-30 ESEM-FEG/PHILIPS microscope. Pollen grains were treated by Physical vapor deposition method under sputter coater. In general, the pollen terminology follows Punt et al. (1994).
| Table 1: | Address of Z. clinopodioides populations |
 | |
| Table 2: | Qualitative studied characters in Z. clinopodioides populations |
 | |
| Table 3: | Quantitative characters studied in Ziziphora clinopodioides |
 | |
Pollens were studied by SEM without any treatment. Anatomical features of leaf were studied by hand made cross sections which were double colored by Carmen and Methylene green.
In order to detect significant differences in studied characters among populations, Analysis of Variance (ANOVA) followed by the Least Significant Differences (LSD) tests were performed. To reveal taxa relationships we used cluster analysis and Principal Component Analysis (PCA). For multivariate analysis the mean of quantitative characters were used while qualitative characters were coded as binary/multi-state characters. Standardized variables were used for multivariate statistical analysis. The average taxonomic distances and squared euclidean distances were used as dissimilarity coefficient in cluster analysis of morphological data. In order to determine the most variable morphological characters among the species studied, factor analysis based on principal components analysis was performed. SPSS ver. 15 was used for statistical analysis.
RESULTS
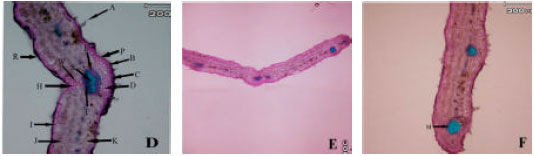
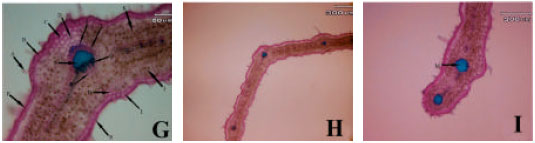
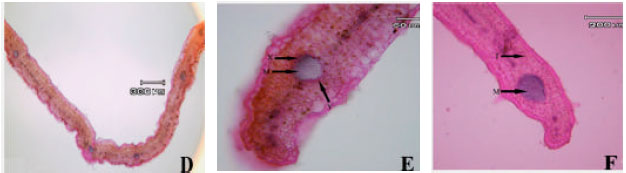
Anatomical observations: In leaf transactions of different populations some differences were observed. In subsp. rigida (Boiss) Rech. f., hair frequency was more in dorsal leaf surface, both surface had cuticle, a single epidermis layer and there are fibers in both adaxial and abaxial leaf surface but adaxial fibers are larger. Sclerenchyma are present between xylem and phloem in mid rib (Hour population) or absent (Alvares population), both leaf surfaces are smooth (Fig. 1A-I). In elburzensis Rech. f. subsp. hair frequency is same in both leaf surface (28 km to Germi population) or dorsal is more (Neor and 5 km to Germi populations). Mid rib surface is smooth, cuticle thickness is same in both leaf surface and fibers are present in both side of leaf (Neor and 5 km to Germi populations) or only in adaxial surface (28 km to Germi) (Fig. 2A-F). Subsp. szowitsii Rech. f., showed more hair frequency in leaf abaxial surface, mid rib surface is smooth, both leaf surfaces had cuticle. There were some sclerenchyma layers between xylem and phloem and some fiber layers in dorsal leaf surface (Fig. 1). In subsp. pseudodasyanta Rech. f., leaf dorsal surface had more hairs and there were observed some glandular hairs. Midrib surface was smooth. There was no sclerenchyma between xylem and phloem (Fig. 1).
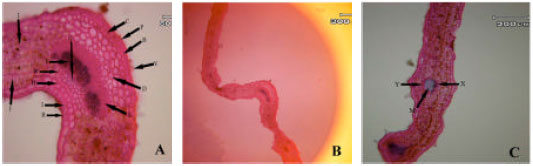 | |
| Fig. 1: | Leaf blade anatomical variations in different Z. clinopodioides subspecies in Iran. First row (A, B and C Z. clinopodioides subsp. pseudodasyantha), middle row (D, E and F, Z. clinopodioides subsp. rigida) and lower row (G, H and I, Z. clinopodioides subsp. szowitsii). (A: Simple hair, B: Glandular hair, C: Upper collenchyma, D: Upper parenchyma, E: Phloem, F: Xylem, G: Lower fibers, H: Lower collenchyma, I: Lower epidermis, J: Palisade parenchyma, K: Spongy parenchyma, L: Upper fibers, S: Sclerenchyma, P: Upper cuticle, R: Lower cuticle, N: Secondary veins, M: Orbicular sclerenchyma, X: Xylem along M, Y: Phloem along M) |
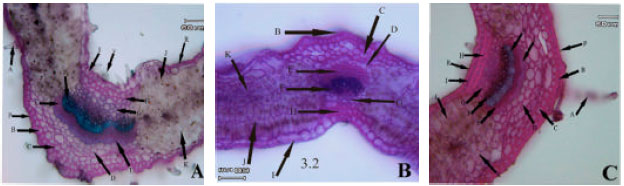 | |
| Fig. 2: | Leaf blade anatomical variations in different populations of Z. clinopodioides subsp. elburzensis in Iran (A, D and E, Neor Lake), (B, 28 km to Germi) and (C and F, 5 km to Germi). Abbreviations as Fig. 1 |
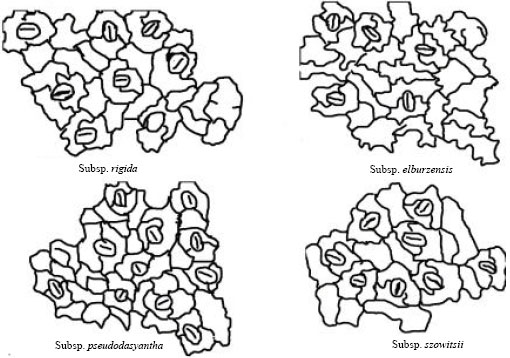 | |
| Fig. 3: | Stomata types of Ziziphora clinopodioides subspecies |
Studying epidermis characters showed that in different subspecies of Z. clinopodioides there are some differences in cell wall shape and cell shape. The epidermal cells are variously shaped with sinuous walls in all studied subspecies. The epidermal cells have wavy walls. The cells exhibit a somewhat quadrangular shape and wavy cell walls in elburzensis subspecies. In subsp. rigida some how arc shape cell walls without much sinuate cell walls were observed (Fig. 3). In all subspecies the stomata are superficial. Although, other stomata types have been reported from the Lamiaceae only anomocytic stomata is recorded in the Z. clinopodioides subspecies here.
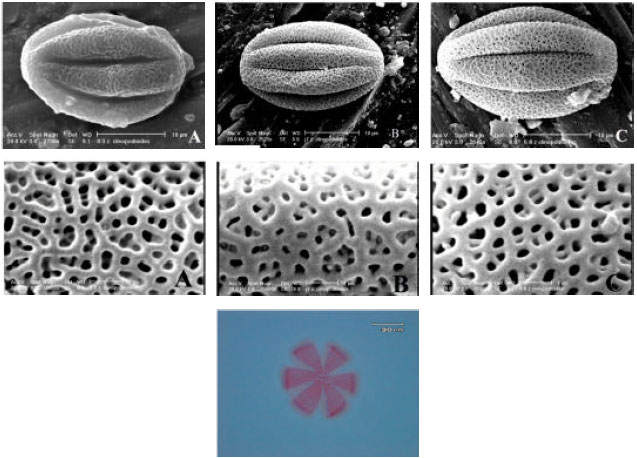 | |
| Fig. 4: | SEM and LM micrographs of Z. clinopodioides subspecies pollen grains. Upper row: Equatorial view, Middle row; Surface detail, Lower row; LM at polar view. (A) elburzensis, (B) rigida and (C) szowitzii |
Palynological comparisons: The pollen grains are monads, six-zonocolpate, prolate, prolate-spheroidal to spheroid as pollen grains did not show particular differences in LM observations, SEM micrographs were used. Polar axis was of 25.9-26.9 μ and an equatorial diameter of 17.1-19.7 μ of the examined populations, the largest pollen grains, on average, were observed in Mazandaran population (Z. clinopodioides subsp. szowitsii) and the smallest pollen grains, were observed in Ghozlou population (subspecies rigida). The longest corpus was observed in subsp. rigida and the shortest was in subsp. elburzensis. The exine sculpture is bireticulate. In the bireticulate ornamentations number of perforations and mesh size are not identical. The average diameter of the small perforations is less than 1.50 μ. Present results are a confirmation of the genus position in Mentheae and Nepetoideae. Pollen grains are illustrated in LM and SEM (Fig. 4). The ectocolpi are distributed symmetrically, elongated, usually shallow and narrowing at the poles.
Morphological observations: Although, there are many thing in common in studied populations but there are fine visible modifications. Some of these differences are related to calyx features. In elburzensis subspecies, calyx length is 3-4.02 mm, plants are glabrous to hairy with hairs up to 0.25 mm length. Hair are very infrequent and sparsely distributed. These hair are straight and white colored. The rigida subspecies have calyx with 3.3-4.7 mm length. Calyx is pedicled and its base is hairy. Hair length is 0.37 to 0.81 mm. Hair are white colored and with medium frequency. In szowitsii calyx is 5.13 mm and calyx hair are about 0.4 mm length. Hair are whitish and dense. Ovate shape leaves are observed in pseudodasyatha subspecies which is mainly covered with dense hairs, 0.31 to 0.597 mm length. Leaves width is more than their length. A comparison of hair at dorsal surface of calyx is given in Fig. 5.
The cluster analysis of Z. clinopodioides subspecies of Iran, based on both quantitative and qualitative characters presented two major clusters (Fig. 6). At first step three subspecies is separated from Z. clinopodioides subsp. szowitsii. There are sub clusters, one is containing populations belonging to Z. clinopodioides subsp. rigida but Z. clinopodioides subsp. elburzensis are inserted in different clusters (one population near rigida, another near pseudodasyantha and one population makes a separate cluster itself). As it is evident Z. clinopodioides subsp. szowitsii, pseudodasyantha and subsp. rigida are clearly separated but it seems that subsp. elburzensis is not clearly limited by the studied set of characters (Fig. 6).
 | |
| Fig. 5: | Calyx dorsal surface in different subspecies of Z. clinopodioides in Iran |
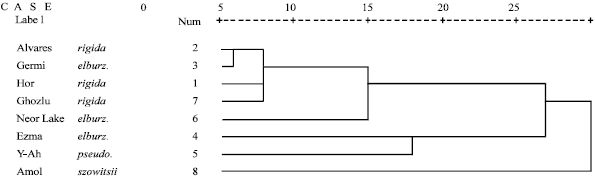 | |
| Fig. 6: | WARD clustering of Z. clinopodioides subspecies based on mean of quantitative and qualitative morphological characters in Iran |
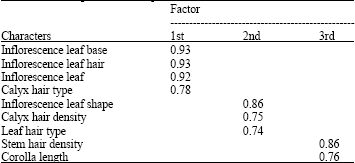
In order to determine the most variable characters among the studied species, Factor analysis based on PCA was performed revealing that the first two factors comprise about 58% of total variation. In the first factor with about 32% of total variation, characters such as inflorescence leaf base and hair and calyx hair type possessed the highest positive correlation (≥0.7). In the second factor with about 26% of total variation, characters as inflorescence leaf shape, calyx hair density and leaf hair type possessed the highest positive correlations. In the third factor with about 14% of total variation, characters as stem hair density and corolla length cause high variation. These are the most variable morphological characters among Z. clinopodioides subspecies in Iran (Table 4).
| Table 4: | Factor analysis results based on morphological characters for Z. clinopodioides subspecies of Iran |
 | |
DISCUSSION
As no one has studied Ziziphora pollen grains before, observations could not be compared with others findings. The studied subspecies are considered to be most closely related to Lycopus L. and Mentha L. in the tribe Mentheae (Briquet, 1896). According to the present results, the pollen morphology in Ziziphora clinopodioides may be helpful in elucidating the infra specific relationships. Studies so far in the Ziziphora indicate that additional sources of potentially very useful characters are available from the pollen morphology. As with any morphological study, the more complete the data, the more convincing the subsequent analyses regarding phylogeny and relationships.
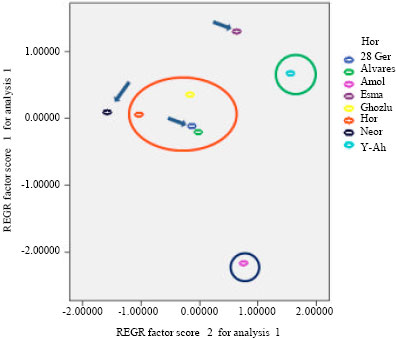 | |
| Fig. 7: | PCA ordination of the Z. clinopodioides subspecies based on morphological characters |
In this study all leaf micro-characters of epidermis were shown to be of great importance in the taxonomy of the Z. clinopodioides subspecies. They allowed in many instances clear separation between various taxa (Fig. 7). The rigida subspecies seems to be clearly separated (red circle in Fig. 7). Other clearly separated subspecies are pseudodasyantha (green circle in Fig. 7) and szowotsii (Dark blue circle). As in Fig. 7 it is evident the elburzensis subspecies has not been clearly separated and is distributed between other subspecies and its limitation needs further considerations. Due to the studied micro and macro-morphological characters an identification key to separate four Z. clinopodioides subspecies has been provided.
| 1- | Inflorescence leaf wider than other leaves | Subsp. szowitsii |
| Inflorescence leaf not wider than other leaves | 2 | |
| 2- | Calyx hair frequent and dense | Subsp. pseudodasyantha |
| Calyx hair sparse | 3 | |
| 3- | Leaf hair dense | Subsp. elburzensis |
| Leaf hair sparse | Subsp. rigida |
CONCLUSION
The use of micro morphological characters as complement of macro-characters for taxonomic purposes is highly recommended to study the specific or infra-specific limits of Lamiaceae. These subspecies were not studied before and this project was the first step in Iran to study these subspecies. The separation of Z. clinopodioides subsp. elburzensis should be revised due to the differences in soil types and habitats occupied by this subspecies.
REFERENCES
- Khodaparast, H., M. Hosein, M.S. Masoumeh, K.R.H. Najafi, M. Bagher and B.T. Shahram, 2007. Effect of essential oil and extract of Ziziphora clinopodioides on yoghurt starter culture activity. World Applied Sci. J., 2: 194-197.
Direct Link
Kalidass Reply
Pollen grains structure is very clear, polynogical study also very good and morphological observation. it is very useful for references.