
F. Khosravinejad
Department of Biology, Faculty of Science, University of Urmia, Iran
R. Heydari
Department of Biology, Faculty of Science, University of Urmia, Iran
T. Farboodnia
Department of Biology, Faculty of Science, University of Urmia, Iran
Pakistan Journal of Biological Sciences
Year: 2008 | Volume: 11 | Issue: 20 | Page No.: 2438-2442







ABSTRACT
Salinity (NaCl Stress) was applied with 50, 100, 200, 300 and 400 mM NaCl. The shoot and root respiration of two barley cultivars (Hordeum vulgare L. variety Afzal and variety EMB82-12) were determined in various concentrations of NaCl. Chlorophyll a,b and total chlorophyll content were significantly decreased, but carotenoids content increased under salinity. Decrease of chlorophyll content in EMB82-12 was higher than Afzal, but carotenoids content in Afzal variety was higher than EMB82-12. Relative Water Content (RWC) was used to indicate the degree of stress. Oxygen uptake declined in shoot and root with increasing NaCl concentrations. Decrease of oxygen uptake in shoot and root of EMB82-12 variety was higher than Afzal variety. RWC decreased with increasing NaCl concentrations. Lowering of RWC reduced growth and increased shoot/root ratio. Decrease of water content in EMB82-12 plants was higher than Afzal plants. Shoot/root ratio in EMB82-12 variety was higher than Afzal.
PDF Abstract XML References Citation
How to cite this article
F. Khosravinejad, R. Heydari and T. Farboodnia, 2008. Effects of Salinity on Photosynthetic Pigments, Respiration, and Water Content in Two Barley Varieties. Pakistan Journal of Biological Sciences, 11: 2438-2442.
DOI: 10.3923/pjbs.2008.2438.2442
URL: https://scialert.net/abstract/?doi=pjbs.2008.2438.2442
DOI: 10.3923/pjbs.2008.2438.2442
URL: https://scialert.net/abstract/?doi=pjbs.2008.2438.2442
INTRODUCTION
Soil salinization is one of the major factors of soil degradation. It has reached 19.5% of the irrigated land and 2.1% of the dry-land agriculture existing on the globe. Salinity effects are more conspicuous in arid and semi-arid areas where 25% of the irrigated land is affected by salts. The increase of salt-affected soils due to poor soil and water management in the irrigated areas, the salinity problem became of great importance for agriculture production in this region. Salinity inhibition of plant growth is the result of osmotic and ionic effects and the different plant species have developed different mechanisms to cope with these effects (Munns, 2002). In suppressing growth (netsynthesis), salinity must decrease the rate of photosynthesis per unit leaf area, the utilization of photosynthate in growth, or both of these. In the case of barley, salinity affected the utilization of photosynthate rather than photosynthesis itself (Barrett-Lennard, 2002). The rate of photosynthesis per unit leaf area may actually be increased in salt-affected plants (Neocleous and Vasilakakis, 2007). Under salt stress, Na concentration increase of plant tissue can result in an increase in oxidative stress, which causes deterioration in chloroplast structure and an associated lose in chlorophyll. This leads to a decrease in the photosynthetic activity. Chlorophyll content decreased, but carotenoid content increased with increase of NaCl concentrations (El-Tayeb, 2005). Salt tolerance is associated with a high rate of photosynthesis and a low rate of respiration. Respiration rates are often an order of magnitude lower than photosynthesis rates. However, since photosynthesis is limited temporally (i.e., day time hours) and spatially (i.e., to green biomass), while respiration occurs continuously in every plant organ, the latter may be an equally important factor controlling productivity, particularly when photosynthesis is largely depressed, such as under salinity conditions (Alutoin et al., 2001). The relative water content decreased significantly with salinity (El-Tayeb, 2005). Furthermore salinity decreased the relative water content in seedling of salt-sensitive cultivars.
MATERIALS AND METHODS
This study was conducted at Biochemistry Laboratory, Department of Biology, Urmia University, Iran, during the Winter of 2007. Barley (Hordeum vulgare L. variety Afzal and variety EMB82-12) seeds were surface sterilized in 0.5% sodium hypochloride solution for 20 min and grown in pots containing vermiculite. Plants were watered every second day using one-half-strength Hoagland nutrient solution in controlled growth room for 4 days, then seedlings were subjected to treatment with 50, 100, 200, 300 and 400 mM NaCl for 3 days. We choose these concentrations according to the results of testing in which concentrations of NaCl (10-1000 mM) were applied. Shoots and roots to be used for biochemical determinations were frozen and stored in liquid nitrogen immediately after harvest until enzyme extraction. MDA content was measured using a 2-thiobarbituric acid reaction (Logan et al., 2006). The chlorophylls and carotenoids (carotene and xanthophylls) content of shoot measured (Xu et al., 2007). The pigments of 0.1 g of shoot fresh weight extracted by acetone 80%. Extracts filtered by filter paper absorbance of samples was measured at 662, 645 and 470 nm by UV-visible spectrophotometer (WPA model S2100). Chlorophyll a, b, total chlorophyll and carotenoids content were measured with following equations:
Chlorophyll a | = | 11.75 A662 – 2.350 A645 |
Chlorophyll b | = | 18.61 A645 – 3.960 A662 |
Total Chlorophyll | = | Chlorophyll a+ Chlorophyll b |
Total Carotenoid | = | (1000A470 - 2.270 Chl a – 81.4 Chl b)/227 |
Oxygen uptake of shoot and root were measured at 25°C using an oxygen meter (WTW model oxi 730). Shoot and root segments (approximately 0.5 g fresh weight) were placed in 4 mL reaction medium (0.25 M sucrose, 0.01 M tris, 0.01 M2HPO4, 0.005 M MgCl2, 0.005 M EDTA, 0.5 mg Ml BSA) adjusted to ph =7.2 with HCl and O2 measured in period of 2 min (Noctor et al., 2007). Relative water content was determined with following equation:
Fresh weight of the plants was measured and after that plants were dried at 105°C until reached constant weight for the determination of dry weight. To determine the turgid weight, samples were soaked in distilled water for 4 h at room temperature (approximately 20°C) and then turgid weight was measured (Fletcher et al., 2006).
Statistical analysis: Mean values were taken from measurements of four replicates and SE of the means was calculated. Differences between means were determined by One-way ANOVA and Turkey`s multiple range tests (p<0.05). Analysis were done using the Statistical Package for Social Sciences (SPSS) for windows (version 13.0).
RESULTS AND DISCUSSION
Salinity caused a reduction in chlorophyll a, b and total chlorophyll content in both varieties, but the decrease in EMB82-12 variety was higher than Afzal variety In 400 mM NaCl, chlorophyll a content was 0.44 fold in EMB82-12 variety and 0.41 fold in Afzal variety as compared to control plants. In highest salinity, chlorophyll b content was 0.24 fold in EMB82-12 variety and 0.54 fold in Afzal variety as compared to control plants and the decrease in Afzal variety was gradually and in EMB82-12 variety in 400 mM NaCl was enormous. Total chlorophyll content decreased in both varieties and in 400 mM NaCl, this factor was 0.38 fold in EMB82-12 variety and 0.44 fold in Afzal variety as compared to control (Fig. 1). Carotenoids content in both varieties increased, but the increase in Afzal variety was higher than EMB82-12 variety.
In 400 from four replicates mM NaCl, carotenoids content was 4.11 fold in EMB82-12 variety and 3.55 fold in Afzal variety as compared to control plants. It means that Afzal plants have higher carotenoids content and lower chlorophyll content than EMB82-12 variety when salt stressed.
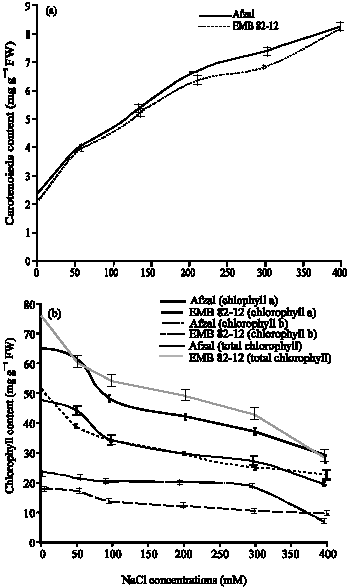 | |
| Fig. 1: | Effects of different (a) NaCl concentrations on photosynthetic pigments carotenoids and (b) chlorophyll in shoots of two barley cultivars. Results are shown as Mean±SE (p<0.05), obtained from four replicates |
The changes in shoot and root respiratory rate could result from damage to the mitochondria themselves or in shoot which altered substrate availability due to inhibition of photosynthesis. Leaf respirations averaged 18.21 μmol O2 g-1 FW min-1 in control plants, EMB82-12 variety and 15.91 μmol O2 g-1 FW min-1 in Afzal variety Under severe salt stress (400 mM NaCl) respiration was lower EMB82-12 variety than Afzal variety, although not significantly different (7.38 μmol O2 g-1 FW min-1in EMB82-12 variety and 8.22 μmol O2 g-1 FW min-1 in Afzal variety). In roots of control plants, respiration averaged 11.0 μmol O2 g-1 FW min-1 in EMB82-12 variety and 10.13 μmol O2 g-1 FW min-1 in Afzal variety Under severe salt stress (400 mM NaCl) respiration was lower than control (4.28 μmol O2 g-1 FW min-1 in EMB82-12 variety Therefore, respiration decreased with increasing NaCl treatments (Fig. 2). In 50 mM NaCl, shoot respiration was 0.85 fold in EMB82-12 variety and 0.8 fold in Afzal variety as compared to control plants and in 400 mM NaCl, shoot respiration was 0.4 fold in EMB82-12 plants and 0.51 fold in Afzal plants as compared to control plants. About root respiration, in 50 mM NaCl, this factor was 0.82 fold in EMB82-12 variety and 0.95 fold in Afzal variety and in 400 mM NaCl, root respiration was 0.36 fold in EMB82-12 variety and 0.52 fold in Afzal variety as compared to control plants. The decrease of respiration rates in roots and shoots in EMB82-12 variety were higher than Afzal variety. The percentage of oxygen consumption decreased gradually with increasing NaCl concentrations (Fig. 2). The percentage of oxygen consumption in 400 mM NaCl, in roots decreased to 42.27% in EMB82-12 variety and 50.16% in Afzal variety and in shoots decreased to 58.25% in EMB82-12 variety and 67.67% in Afzal variety. The decrease of oxygen consumption in EMB82-12 variety was higher than Afzal variety and in roots was higher than shoots.
In roots and shoots in control plants, EMB82-12 variety have higher water content than Afzal variety, but in severe salinity, Afzal plants roots and shoots have higher water content than EMB82-12 plants (Fig. 3). Relative water content in roots and shoots decreased with increasing NaCl concentrations. In 400 mM NaCl, this decrease was enormous in roots and shoots in both varieties. In 400 mM NaCl, RWC was 0.4 fold in EMB82-12 variety and 0.62 fold in Afzal variety in roots and 0.53 fold in EMB82-12 variety and 0.75 fold in Afzal variety in shoots as control plants. With increase of NaCl concentrations, the shoot/root ratio was increased and shoot/root ratio in EMB82-12 plants was higher than Afzal plants (Fig. 3). In 400 mM NaCl, shoot/root ratio was 1.34 fold in EMB82-12 plants and 1.22 fold in Afzal plants as compared to control.
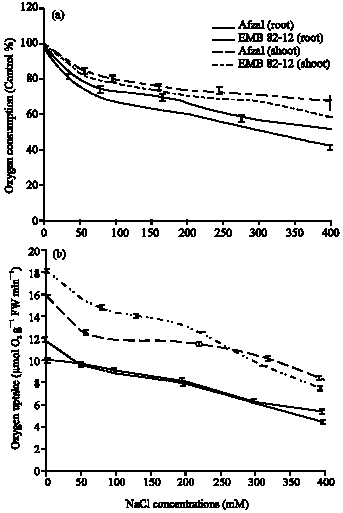 | |
| Fig. 2: | Effects of different (a) NaCl concentrations on oxygen uptake and oxygen consumption (b) percent in roots and shoots of two barley cultivars. Results are shown as Mean±SE (p<0.05), obtained from four replicates |
Reductions of chlorophyll content under salinity conditions were observed for some salt–Bsensitive plant species (Delfine et al., 1999; Ashraf et al., 2002; Jungklang et al., 2004; Lee et al., 2004). The decrease of chlorophyll content was dependent on the salinity level, the time of exposure to salts and the species. In contrast, Chl. content in salt-tolerant plants either dose not decline or else rises with increasing salinity (Brugnoli and Björkman, 1992; Qiu and Lu, 2003). Chlorophyll concentration can be used as a sensitive indicator of the cellular metabolic state; thus its decrease signifies toxicity in tissues due to accumulation of ions. In present experiments, chlorophyll a and b contents and total chlorophyll decreased with increasing NaCl supply (Yeo and Flowers, 2006). In EMB82-12 plants chlorophyll content in shoots decreased higher than Afzal plants, but in severe salinity, EMB82-12 plants have higher total chlorophyll content than Afzal plants.
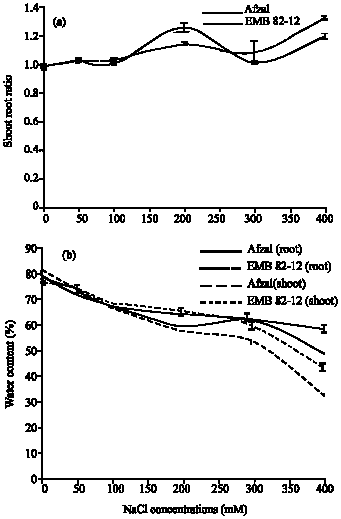 | |
| Fig. 3: | Effects of different (a) NaCl concentrations on shoot/root ratio and water content (b) in roots and shoots of two barley cultivars. Results are shown as Mean±SE (p<0.05), obtained from four replicates |
Increase of carotenoids content in Afzal variety was higher than EMB82-12 plants. Therefore, Afzal plants have a better protection than EMB82-12 plants, because carotenoids have a protective role and protect chlorophyll from photo oxidation. Increasing NaCl concentration clearly depressed both roots and shoots respiration. Leaf respiration rates decreased under salinity in most species, but the decline was always smaller than that of photosynthesis, therefore resulting in decreased photosynthesis to respiration ratio (indicative of shoot carbon balance). The decline in respiration in response to salinity seems to be part of a systemic metabolic response, which occurs under conditions where salinity severely restricts CO2 availability inside leaf cells, therefore, creating the risk of a secondary oxidative stress (Flexas et al., 2008). The original objective from this study was finding the changes in respiration rates in shoots and roots with increasing NaCl treatments. We found that with increase of salinity, respiration rates decreased in both shoots and roots. These results supported earlier findings (Valentini et al., 2000). In 400 mM NaCl, respiration rate in EMB82-12 plants in roots and shoots were not only lower than the control, but were also lower than in Afzal variety. It means that in leaves and roots the decrease of oxygen uptake in EMB82-12 variety was higher than Afzal variety. With increase of NaCl concentrations, oxygen consumption percentage decrease in both roots and shoots. The decrease of oxygen consumption percentage in EMB82-12 plants was higher than Afzal plants. Therefore salinity has a higher effect in EMB82-12 plant`s respiration than Afzal plants. Oxygen consumption percentage in high salinity in roots was lower than shoots and roots were more sensitive than shoots. Salinity probably acted directly on roots, because the roots were immersed in NaCl solutions and salinity in roots was higher than shoots, whereas shoots could reduce oxygen consumption by stoma regulations. There were a positive and strong correlation between respiration rates and water content. The decreased respiration rate was positively correlated to decrease of relative water content. Respiration rates affected by a decrease in water content. It means that a decrease in water content caused a decreased in oxygen consumption and respiration rates (Takemura et al., 2000). The decrease of water content in EMB82-12 plants was higher than Afzal plants in roots and shoots. Afzal plants have higher water content than EMB82-12 plants in roots and shoots. The shoot/root ratio was increased in salinity and this factor in EMB82-12 variety was higher than Afzal variety. The above results suggest that plants of the Afzal variety have a better tolerance to salinity as compared to EMB82-12 variety.
ACKNOWLEDGMENTS
This study supported by Department of Biology of Urmia University, Iran. We greatly acknowledge Dr. L. Purakbar and Dr. S. Fallahi for their support.
REFERENCES
- Alutoin, S., J. Boberg, M. Nystrom and M. Tedengren, 2001. Effects of the multiple stressors copper and reduced salinity on the metabolism of the hermatypic coral Porites lutea. Mar. Environ. Res., 52: 289-299.
CrossRef - Brugnoli, E. and O. Bjorkman, 1992. Growth of cotton under continuous salinity stress: Influence on allocation pattern, stomatal and non-stomatal components of photosynthesis and dissipation of excess light energy. Planta, 187: 335-345.
CrossRef - El-Tayeb, M.A., 2005. Response of barley grains to the interactive e.ect of salinity and salicylic acid. Plant Growth Regul., 45: 215-224.
CrossRefDirect Link - Flexas, J., J. Bota, F. Loreto, G. Cornic and T.D. Sharkey, 2004. Diffusive and metabolic limitations to photosynthesis under drought and salinity in C3 plants. Plant Biol., 6: 269-279.
CrossRefDirect Link - Jungklang, J., Y. Sunohara and H. Matsunoto, 2004. Antioxidative enzymes response to NaCl stress in salt-tolerant Sesbania rostrata. Weed Biol. Manage., 4: 81-85.
CrossRef - Lee, G., R.N. Carrow and R.R. Duncan, 2004. Photosynthetic responses to salinity stress of halophytic seashore paspalum ecotypes. Plant Sci., 166: 1417-1425.
CrossRef - Logan, B.A., D. Kornyeyev, J. Hardison and A.S. Holaday, 2006. The role of antioxidant enzymes in photoprotection. Photosynthesis Res., 88: 119-132.
CrossRef - Munns, R., 2002. Comparative physiology of salt and water stress. Plant Cell Environ., 25: 239-250.
CrossRefDirect Link - Neocleous, D. and M. Vasilakakis, 2007. Effects of NaCl stress on red raspberry (Rubus idaeus L. Autumn Bliss). Scientia Hortic., 112: 282-289.
CrossRefDirect Link - Noctor, G., R. de Paepe and Ch.H. Foyer, 2007. Mitochondrial redox biology and homeostasis in plants. Trends Plant Sci., 12: 125-134.
CrossRef - Qiu, N. and C. Lu, 2003. Enhanced tolerance of photosynthesis against high temperature damage in salt-adapted halophyte Atriplex centralasiatica plants. Plant Cell Environ., 26: 1137-1145.
CrossRef - Takemura, T., N. Hanagata, K. Sugihara, S. Baba, I. Karube and Z. Dubinsky, 2000. Physiological and biochemical responses to salt stress in the mangrove, Bruguiera gymnorrhiza. Aquat. Bot., 68: 15-28.
CrossRefDirect Link - Valentini, R., G. Matteucci, A.J. Dolman, E.D. Schulze and C. Rebmann et al., 2000. Respiration at the main determinant of carbon balance in European forests. Nature, 404: 861-865.
CrossRef - Xu, C.M., B. Zhao, X.D. Wang and Y.C. Wang, 2007. Lanthanum relieves salinity-induced oxidative stress in Saussurea involucrate. Biol. Plantarum, 51: 567-570.
CrossRef - Yeo, A.R. and T.J. Flowers, 1983. Varietal differences in the toxicity of sodium ions in rice leaves. Physiologia Plantarum, 59: 189-195.
CrossRefDirect Link - Barrett-Lennard, E.G., 2002. Restoration of saline land through revegetation. Agric. Water Manage., 53: 213-226.
CrossRef - Ashraf, M., F. Karim and E. Rasul, 2002. Interactive effects of gibberellic acid (GA3) and salt stress on growth, ion accumulation and photosynthetic capacity of two-spring wheat (Triticum aestivum L.) cultivars differing in Salt tolerance. Plant Growth Reg., 36: 49-59.
CrossRef - Fletcher, R.A., M. Santakumari and D.P. Murr, 1988. Imposition of water stress in wheat seedlings improves the efficiency of uniconazole induced thermal resistance. Physiol. Plant., 47: 360-364.
CrossRefDirect Link