
P.F. Omojasola
Department of Microbiology, University of Ilorin, Ilorin, Nigeria
O.P. Jilani
Department of Microbiology, University of Ilorin, Ilorin, Nigeria
Pakistan Journal of Biological Sciences
Year: 2008 | Volume: 11 | Issue: 20 | Page No.: 2382-2388







ABSTRACT
The wastes materials from the sweet orange (Citrus sinensis) were used as substrate for the production of cellulase. The rind, the pericarp or albedo and the pulp were hydrolyzed by cellulolytic enzymes of Trichoderma longibrachiatum, Aspergillus niger and Saccharomyces cerevisiae after they were treated with alkali and steam. The amount of glucose released from the substrates following the secretion of cellulase by the three microorganisms was measured. The orange wastes released amounts of glucose ranging from 0.76-0.96 mg mL-1 by Trichoderma longibrachiatum, 0.90-1.08 mg mL-1 by A. niger and 0.60-0.76 mg mL-1 by S. cerevisiae after five days of fermentation. The conditions of the fermentation were then varied to determine their effect on cellulase production. Fermentation parameters varied were time, pH, substrate concentration, temperature and inoculum size. After this, conditions that produced highest amounts of glucose were combined in an optimization experiment. Glucose production under optimized conditions were 0.94 mg mL-1 by T. longibrachiatum, 0.83 mg mL-1 by A. niger and 0.67 mg mL-1 by S. cerevisae. The activity of the test organisms` cellulase against CMC on the orange wastes was also determined with T. longibrachiatum producing 3.86 mg mL-1, A. niger 2.94 mg mL-1 and S. cerevisiae 2.30 mg mL-1 glucose amounts all from orange pulp.
PDF Abstract XML References Citation
How to cite this article
P.F. Omojasola and O.P. Jilani, 2008. Cellulase Production by Trichoderma longi, Aspergillus niger and Saccharomyces cerevisae Cultured on Waste Materials from Orange. Pakistan Journal of Biological Sciences, 11: 2382-2388.
DOI: 10.3923/pjbs.2008.2382.2388
URL: https://scialert.net/abstract/?doi=pjbs.2008.2382.2388
DOI: 10.3923/pjbs.2008.2382.2388
URL: https://scialert.net/abstract/?doi=pjbs.2008.2382.2388
INTRODUCTION
Orange is widely planted and is relatively less perishable than most other tropical fruits. In some fruits, the discarded portion can be very high, for example, orange 30-50% (Lerner, 2003). The recent thrust in bioconversion of agricultural and industrial wastes to chemical feedstock has led to extensive studies on cellulolytic enzymes produced by fungi and bacteria (Baig et al., 2004). Cellulase production from different agro-wastes have been reported using a variety of microorganisms. These include banana peel, millet, guinea corn, rice husk and maize straw using Saccharomyces cerevisae and Aspergillus niger (Baig et al., 2004; Milala et al., 2005). Shahera and Sanaa (2002) have also reported cellulase production by yeast strains from orange wastes. Despite a worldwide and enormous utilization of natural cellulosic sources, there are still abundant quantities of cellulose-containing raw materials and waste products that are not exploited or which could be used more efficiently. The problem in this respect is however to develop processes that are economically profitable. Cellulose is a useful as a digestive aid especially in animals. It is also useful in the production of glucose syrups, paper milling, production of single-cell protein and improving the palatability of poor quality vegetables. Cellulose-containing wastes may be agricultural, urban, or industrial in origin, sewage sludge might also be considered a source of cellulose since its cellulosic content provides the carbon needed for methane production in the anaerobic digestion of sludge. Cellulose which forms about 40-50% of plant`s composition is the most abundant organic matter on earth. Proper biotechnological utilization of these wastes in the environment will eliminate pollution and convert them into useful by-products (Milala et al., 2005). Thus the agro wastes left behind for natural degradation can be utilized effectively to yield fermentable sugars which can be converted into other substance like alcohol. The crystallinity and lignification limit the accessibility and susceptibility of cellulose to cellulolytic enzymes and other hydrolytic agents (Caritas and Humphrey, 2006).
Pretreatment of cellulose opens the structure and removes secondary interaction between glucose chains (Tang et al., 1996). Various physical and chemical treatments of lignocellulosic substrates are necessary for lignin removal by effective disruption of the lignin carbohydrate linkage and the highly ordered cellulose itself. Pretreatment of the substrate results in the reduction of the particle size by increasing surface to volume ratio and causes reduction in crystallinity. The chemical treatment of lignocelluloses causes swelling leading to an increase in internal surface area, decrease in the degree of polymerization, decrease in crystallinity, separation of structural linkages between lignin and carbohydrates, thus, increasing the cellulose hydrolysis (Ravindra and Bhaathi, 2006).
Nigeria possesses abundant cellulolytic wastes as leftovers from agricultural practices. These wastes can be used to produce alcohol which can be exported to gain much needed foreign exchange and boost our foreign reserves. Processing of these wastes can also help provide jobs and assist in controlling environmental pollution. This study therefore evaluates the possibility of the re-use of agro-industrial wastes namely orange peel as a substrate for cellulose production.
MATERIALS AND METHODS
Plant material and microorganisms: This study was carried out in 2007. Orange fruit was washed thoroughly with water, peeled and sliced. The juice was removed with the aid of a squeezer and the pulp separated from the pericarp (albedo) and the three materials were sun-dried separately. They were later oven-dried at 70°C, still being handled separately and then pounded using a mortar and pestle. These materials were subjected to alkali and steam pre-treatment, after which the materials were washed and dried in the oven at 70°C. All waste substrates were then ground separately using a blender and kept in containers on the shelf. The organisms used for this study were Trichoderma longibrachiatum, Aspergillus niger and Saccharomyces cerevisiae. These were isolated from three sources: Aspergillus niger was isolated from rotten wood (RW) picked up on the premises of Unilorin permanent site, Saccharomyces cerevisiae was isolated from palm wine (PW) bought from a palm wine tapper at Offa garage in Ilorin and Trichoderma longibrachiatum was collected from the Faculty of Agriculture, University of Ilorin.
Screening and identification of microorganisms: Plate screening of the isolated organisms for cellulase production was carried out according to the method of Brown et al. (1987). A point of inoculation the spores of each organism was grown on PDA supplemented with 2% (w/v) carboxymethylcellulose (CMC). The plates were incubated at 29±1°C for 48 h after which they were stained with Congo red stain for 15 min. Excess dye was removed by washing with 1 M NaCl and the plates were fixed with 1 N HCl. The production of extracellular cellulase by the organisms was indicated by a zone of clearance around the colony.
The microorganisms were identified in Microbiology Laboratory in the University of Ilorin according to Chaturvedi (2001). All organisms were maintained on PDA slants. A spore suspension from 3 to 4 day old 10 mL PDA slants of each culture in 10 mL sterile distilled water was made. Mineral salts glucose medium was prepared and approximately 2.8x106 spores cells-1 of each culture were inoculated into 500 mL flasks containing 100 mL of medium each. The spores cells-1 were counted using a Neubauer counting chamber. The flasks were incubated for 24 h at 29±1°C on a Gallenkamp (England) rotary shaker at 250 rpm to develop the inoculum. Mary Mandels` Mineral salts solution was used for the fermentation and it was prepared as described by Jeffries (1996). Culture conditions involved autoclaving the fermenting media containing 10 g L-1 of waste substrate at pH 5.0 and inoculating with pure suspension of germinated spores of Trichoderma longibrachiatum, Aspergillus niger and Saccharomyces cerevisiae.
Chemicals and reagents: Carboxymethylcellulose (CMC), Sodium potassium tartarate, Glucose, Dinitrosalicylic acid, Sodium hydroxide and all other salts used were of analytical grade and products of British Drug House (BDH), England.
Enzyme assays: The waste substrates represent the carbon sources in the fermentation media. These were combined with Mary Mandels` Mineral Salts Medium to give Mineral salts glucose medium (MSGM) that was used in the inoculum development, Mineral salts orange peel medium (MSOpeM), Mineral salts orange pulp medium (MSOpuM) and Mineral salts orange albedo medium (MSOalM).
All the media mentioned above were prepared separately and dispensed in conical flasks. They were sterilized in the autoclave at 121°C for 15 min.
The final pH was adjusted to 5.0 using a pH meter (Denver Instrument, Model 20 pH/Conductivity meter).
The pH of the fermenting media containing the waste substrates at a level of 10 g L-1 was adjusted to 5.0. The suspension of germinated spores was inoculated at a level of 10% (v/v) into the production medium contained in flasks. These were incubated at 29±1°C on a shaker at 100 rpm. Glucose production in the medium was measured on Day 5 of fermentation (Srivastava et al., 1987; Jeffries, 1996).
Cellulase activity was determined colorimetrically by measuring the increase in reducing groups by the hydrolysis of a carboxymethylcellulose (CMC) substrate. The procedure followed the 0.5 mL assay described by Jeffries (1996).
Samples were withdrawn from the culture at 2 day intervals over a period of 7-9 days and the supernatant that resulted following centrifugation at 3,000 rpm for 15 min to remove solids, were assayed for total reducing sugars using DNSA method of Miller (1959). Enzyme solutions were diluted in 0.05 M citrate buffer, pH 4.8. The enzyme diluted in buffer and 1% CMC (0.5 mL each) was mixed well and incubated for 30 min at 50°C. Three milliliters of the DNSA was added and the tubes were placed in boiling water bath for 5 min. The tubes were cooled and the reducing sugar, glucose was determined (Jeffries, 1996). The sample, enzyme blank, glucose standard and control were boiled together and absorbance was read at 540 nm using a spectrophotometer (CamSpecM105). A control (substrate and buffer) otherwise called spectro zero, was used to set the spectrophotometer at zero absorbance. During the course of the experiments, the absorbance of the sample tube, corrected by subtraction of the enzyme blank was translated into glucose during the reaction using a glucose standard. The linear glucose standard was used to translate the absorbance values of the sample tubes into glucose i.e., mg glucose produced during the reaction. For a 30 min assay, 1 mg of glucose equals 0.185 unit
Optimization experiments were carried out and each of the organisms were grown on each of the substrates and hydrolyzed under conditions that produced maximal activity of the enzyme from all the previous experiments. In accord with the International Union of Biochemistry, one enzyme unit equals 1 μmol of substrate hydrolyzed per minute.
Determination of optimal conditions for enzyme production
Effect of varying time: Cellulase activity was measured at regular intervals while fermentation was observed at 29±1°C for a period of 9 days and the period of maximum enzyme production was determined. Samples were withdrawn on Days 0, 1, 3, 5, 7 and 9.
Effect of varying pH: The pH of the fermentation media were adjusted to various values ranging from 2.0-6.0 with 0.1 N NaOH or 0.1 N HCl. The pH was determined using the pH meter (Denver Instrument, Model 20 pH/Conductivity meter).
Effect of varying substrate concentration: Different concentration of the waste substrates (orange pulp, albedo and peel), ranging from 1.0 to 5.0% were used in the fermentation media.Effect of varying temperature: The fermentation was carried out at different temperatures ranging from 29±1 to 45°C.
Effect of varying inoculum size: Each cellulosic waste was fed with varying sizes of inoculum of the organisms. The inoculum size was varied from 2 to 10%.
RESULTS AND DISCUSSION
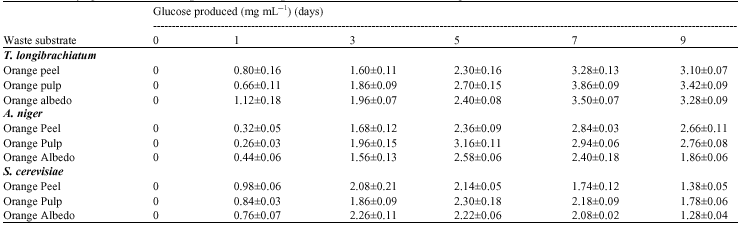
The three test organisms can produce cellulase from the orange wastes (Table 1). In determining optimal conditions for glucose production, T. longibrachiatum produced highest amounts of glucose on day 7, A. niger on day 5 and from S. cerevisiae on day 3 for orange peel and albedo and day 5 for orange pulp (Table 2). Hydrolysis rates decline with time due to depletion of the more amorphous substrates, product inhibition and enzyme inactivation (Ghose, 1987). Caritas and Humphrey (2006) and Narasimha et al. (2006) also gave similar time course reports of maximum glucose yield on 5th day of fermentation using A. niger. Effect of pH on glucose production from the three waste substrates by the three microorganisms (Table 3) supports the findings of Lee et al. (2002) who reported that CMCase, Avicelase and FPase activities exhibit a pH optimum of approximately 4, while the pH optimum of β-glucosidase was between pH 5 and 6.
Further increase in cellulose concentration beyond the level that gave the optimum glucose did not result in proportionate increase in glucose yield. Haapela et al. (1995) and Jeffries (1996) reported that maximum endoglucanase activity was recovered on the medium with cellulose at 10 g L-1. Mandels and Reese (1959) also reported that maximal yields of cellulase were obtained on 1% substrate (cellulose, lactose, cellobiose and glucose) using T. viride and Myrothecium verrucia. These reports support the findings of this study as substrate concentration of 10 g L-1 gave the highest amount of glucose from T. longibrachiatum on orange albedo.
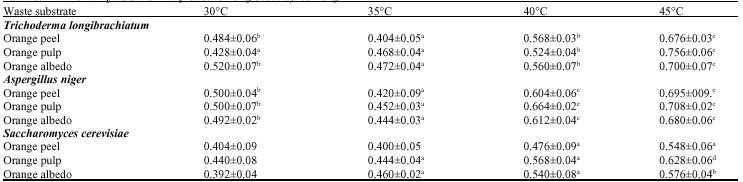
| Table 1: | Fermentation of waste substrates by test fungi |
 | |
| Values are presented as Mean±SD (n = 3) | |
| Table 2: | Effect of varying time on glucose production by test fungi |
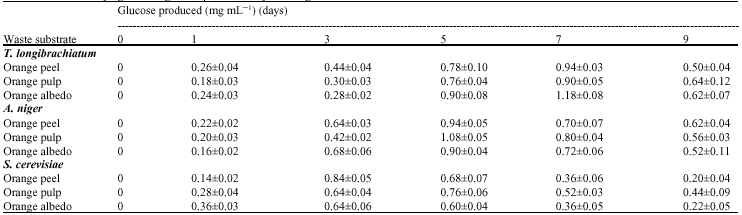 | |
| Values are presented as Mean±SD (n = 3) | |
| Table 3: | Effect of pH on the production of glucose by test fungi |
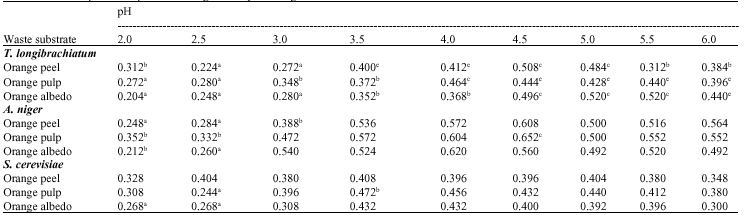 | |
| (Substrate concentration: 1%, Temp: 29±1°C, inoculum size: 10%), Waste substrate: 1%, Temperature: 29±1°C, pH: 5.0, inoculum size: 10%, Values with different superscripts are statistically different. Values are presented as Mean±SD (n = 3) | |
| Table 4: | Effect of substrate concentration on the production of glucose by test fungi |
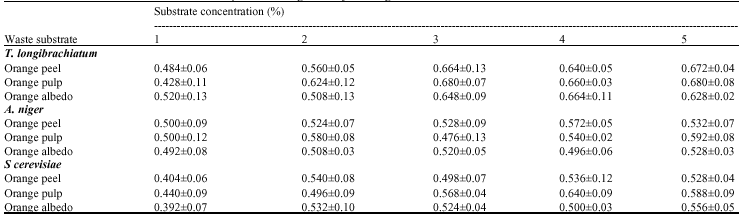 | |
| (pH: 5.0, Temperature: 29±1°C , Inoculum size: 10%), Values are presented as Mean±SD (n = 3), All groups are compared to each other at p<α = 0.05. Values with different superscripts are statistically different | |
Since the substrates contain different minerals apart from carbon which may serve as nutrient supplements, increase in substrate concentration leads to increase in these nutrients which may adversely affect the cell concentration (Table 4). The increase in glucose production until the optimum that was obtained was due to the availability of cellulose in the medium; while a decrease in production beyond optimum concentration is explained to be as a result of an inhibitory effect of accumulated cellobiose and cellodextrins of low degree of polymerization to the growth medium. It might also be due to the specific binding of the enzymes with the substrates (Wang et al., 2006). Low glucose production after optimum very probably highlights sugar depletion from the substrates into the medium (Brien and Craig, 1996).
Decrease in amounts of glucose production resulted at inoculum sizes above 6% for fermentations using A. niger (Table 5). This decrease in glucose production with further increase in inoculum might be due to clumping of cells which could have reduced sugar and oxygen uptake rate and also, enzyme release (Srivastava et al., 1987). The optimum temperature for the synthesis of enzymes for saccharification of agrowaste in all cases to enzymatic hydrolysis can be attributed to lignin content of the material (Table 6). Pretreatment of lignocellulosic material enhances enzyme activity and maximum saccharification was achieved within the range 30-45°C coinciding with the characteristics of mesophiles (Baig et al., 2004).
Optimum glucose from the waste substrates using T. longibrachiatum, was produced at 10% inoculum size at 45°C on day 7 but at pH 5.0 and 3% substrate concentration for orange peel but 1% substrate concentration for orange albedo and pH 4.0 and 3% substrate concentration for orange pulp.
Optimum glucose from the waste substrates using A. niger was produced at pH 4.5, 6% inoculum size on day 5 but at 2% substrate concentration and 40°C for orange pulp; 4% substrate concentration and 45°C for orange peel (Table 7).
For S. cerevisiae fermentation, optimum glucose was produced at pH 4.5, 2% substrate concentration, 6% inoculum size, 45°C on day 3 for orange peel, pH 3.5, 4% substrate concentration, 2% inoculum size and 45°C on day 5 for orange pulp and pH 3.5, 2% substrate concentration, 2% inoculum size and 45°C on day 3 for orange albedo.
These optimal conditions were combined in single fermentations for each organism and cellulase activity was measured (Table 8). Cellulase activity from orange peel was 1.64 U mL-1 when hydrolyzed by T. longibrachiatum, 1.42 U mL-1 when hydrolyzed by A. niger and 1.07 U mL-1 when hydrolyzed by S. cerevisiae. Cellulase activity from orange pulp was 1.93 U mL-1 when hydrolyzed by T. longibrachiatum, 1.58 U mL-1 when hydrolyzed by A. niger and 1.15 U mL-1 when hydrolyzed by S. cerevisiae. Cellulase activity from orange albedo was 1.75 U mL-1 when hydrolyzed by T. longibrachiatum, 1.29 U mL-1 when hydrolyzed by A. niger and 1.13 U mL-1 when hydrolyzed by S. cerevisiae. Cellulase activity of S. cerevisiae was lowest. The exo-β-1, 3-glucanases produced by S. cerevisiae yield glucose as the end product, whereas endo-β-1, 3-glucanase releases a mixture of oligosaccharides with glucose as the minor product. Because β-1, 3-glucan is the main structural polysaccharide responsible for the strength and rigidity of the yeast cell wall, β-1, 3-glucanases have been suggested to play a role in important morphogenetic processes involving the controlled autolysis of β-1, 3 glucan. During vegetative growth, several endo-and exo-1, 3-β-glucanases are synthesized, some of which are secreted only to remain entrapped in the cell wall whereas others are released to the surrounding medium (Lee et al., 2002).
| Table 5: | Effect of inoculum size on the production of glucose by test fungi |
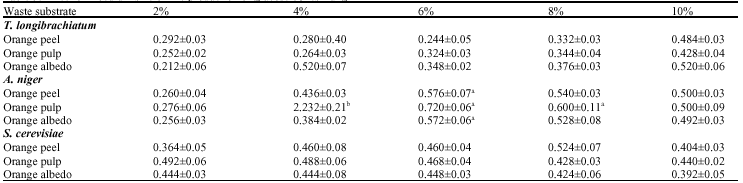 | |
| (Temperature: 29±1°C, pH: 5.0, Substrate concentration: 1%), Values are presented as Mean±SD (n = 3), All groups are compared to each other at p<α = 0.05. Values with different superscripts are statistically different | |
| Table 6: | Effect of temperature on the production of glucose by test fungi |
 | |
| Values are presented as Mean±SD (n = 3), All groups are compared to each other at p<α=0.05. Values with different superscripts are statistically different | |
| Table 7: | Optimized glucose production by the test organisms from the different waste substrates |
 | |
| Values are presented as Mean±SD (n = 3), All groups are compared to each other at p<α = 0.05. Values with different superscripts are statistically different | |
| Table 8: | Activity against CMC of T. longibrachiatum, A. niger and S. cerevisae cellulase on orange wastes |
 | |
| Values are presented as Mean±SD (n = 3) | |
In conclusion, this study revealed that orange peel, pulp and albedo, which are examples of domestic and industrial agro-wastes, produce large amounts of cellulase enzymes when hydrolyzed by cellulolytic microorganisms and instead of being left behind for natural degradation can be utilized effectively under these conditions, to produce cellulase.
REFERENCES
- Baig, M.M.V., M.L.B. Baig, M.I.A. Baig and Y. Majeda, 2004. Saccharifycation of banana agro waste by cellulolytic enzymes. Afr. J. Biotechnol., 3: 447-450.
Direct Link - Brien, D.J.O. and J.C. Jr. Craig, 1996. Ethanol production in a continuous fermentation/membrane evaporation system. Appied Microbiol. Biotechnol., 44: 699-704.
CrossRef - Brown, J.A., D.J. Falconer and T.M. Wood, 1987. Isolation and Properties of Mutants of the fungus Penicillium pinophilum with enhanced Cellulase and β-glucosidase production. Enzyme Microbiol. Technol., 9: 169-175.
CrossRef - Caritas, U.O. and C. N. Humphrey, 2006. Effect of Acid Hydrolysis of Garcinia kola (bitter Kola) pulp waste on the production of CM-cellulase and β-glucosidase using Aspergillus niger. Afr. J. Biotechnol., 5: 819-822.
Direct Link - Ghose, T.K., 1987. Measurement of cellulase activities. Pure Applied Chem., 59: 257-268.
CrossRefDirect Link - Haapela, R., E. Parkkinen, P. Susminen and S. Unko, 1995. Production of extracellular enzymes by immobilized Trichoderma reesei in shake flask cultures. Applied Microbiol. Biotechnol., 43: 815-821.
CrossRef - Lynd, L.R., P.J. Weimer, W.H. van Zyl and I.S. Pretorius, 2002. Microbial cellulose utilization: Fundamentals and biotechnology. Microbiol. Mol. Biol. Rev., 66: 506-577.
CrossRefPubMedDirect Link - Mandels, N. and E.T. Reese, 1959. Induction of cellulase in fungi by cellobiose. Res. Eng., 79: 816-826.
PubMed - Miller, G.L., 1959. Use of dinitrosalicylic acid reagent for determination of reducing sugar. Anal. Chem., 31: 426-428.
CrossRefDirect Link - Narasimha, G., A. Sridevi, B. Viswanath, M.S. Chandra and R.B. Rajasekhar, 2006. Nutrient effects on production of cellulolytic enzymes by Aspergillus niger. Afr. J. Biotechnol., 5: 472-476.
Direct Link - Ravindra, P. and K. Bharathi, 2006. Pretreatment studies of rice bran for the effective production of cellulase. EJEAFChe, 5: 1253-1264.
Direct Link - Srivastava, S.K., K.S. Gopalkrishnan and K.B. Ramachandran, 1987. The production of β-glucosidase in shake-flasks by Aspergillus wentii. J. Ferment. Technol., 65: 95-99.
CrossRef - Tang, L.G., D.N.S. Hon, S.H. Pan, Y.Q. Zhu, Z. Wang and Z.Z. Wang, 1996. Evaluation of microcrystalline cellulose changes in ultra structural characteristics during preliminary acid hydrolysis. J. Appied Polymer Sci., 59: 483-488.
Direct Link - Wang, X.J., J.G. Bai and Y.X. Lian, 2006. Optimization of multienzyme production by two mixed strains in solid-state fermentation. Applied Microbiol. Biotechnol., 73: 533-540.
CrossRefDirect Link