
L.S. Khorsandi
Department of Anatomical Sciences, Faculty of Medicine, Ahwaz Jundi-Shapour University of Medical Sciences, Ahwaz, Iran
M. Hashemitabar
Department of Anatomical Sciences, Faculty of Medicine, Ahwaz Jundi-Shapour University of Medical Sciences, Ahwaz, Iran
M. Orazizadeh
Department of Anatomical Sciences, Faculty of Medicine, Ahwaz Jundi-Shapour University of Medical Sciences, Ahwaz, Iran
N. Albughobeish
Department of Histology, Faculty of Veterinary Sciences, University of Shahid Chamran, Ahwaz, Iran
Pakistan Journal of Biological Sciences
Year: 2008 | Volume: 11 | Issue: 18 | Page No.: 2231-2236







ABSTRACT
The aim of this study is to investigate the effect of Dex, a widely used GC, on apoptosis and expression of FasL protein in the mouse testicular germ cells. The effects of dexamethasone (Dex) on expression of Fas ligand (FasL), an important proapoptotic protein, in the mouse testicular germ cells were investigated. Six groups, each of 8 male NMRI mice were chosen for the experiment. Experimental groups received one of the following treatments daily for 7 days: 4, 7 and 10 mg kg-1 Dex. Control groups were treated with equivalent volumes of saline. Experimental and control animals were sacrificed 24 h after the last injection. Immunohistochemical procedure was used to evaluation of FasL expression and the deoxyuridine nick-end labeling (TUNEL) was applied to assessment of the apoptotic germ cells. FasL expression of testicular germ cells were significantly increased in 10 mg kg-1 Dex treated mice (p<0.05), particularly at stages VII-VIII of spermatogenic cycle. Apoptotic indexes (AIs) of germ cells were significantly increased in 7 and 10 mg kg-1 Dex treated mice.
PDF Abstract XML References Citation
How to cite this article
L.S. Khorsandi, M. Hashemitabar, M. Orazizadeh and N. Albughobeish, 2008. Dexamethasone Effects on Fas Ligand Expression in Mouse Testicular Germ Cells. Pakistan Journal of Biological Sciences, 11: 2231-2236.
DOI: 10.3923/pjbs.2008.2231.2236
URL: https://scialert.net/abstract/?doi=pjbs.2008.2231.2236
DOI: 10.3923/pjbs.2008.2231.2236
URL: https://scialert.net/abstract/?doi=pjbs.2008.2231.2236
INTRODUCTION
GCs are extraordinary hormones that influence the activity of almost every cell in the body. They modulate the expression of approximately 10% of our genes and are essential for life but are also increasingly implicated in the pathogenesis of disease and produce many unwanted effects when given therapeutically (Julia, 2006). In therapeutic concentrations, GCs are strongly immunosuppressive and anti-inflammatory, which has made them one of the most prescribed drugs worldwide.
It has been reported that GCs cause changes in plasma gonadotrophin levels and their pituitary content and indirectly contribute to the inhibition of reproductive functions (Calogero et al., 1999). Earlier studies have been shown that elevation of GC concentration precedes a decline in testosterone concentration in the male (Dong et al., 2004; Bernier et al., 1999; Hardy et al., 2005). Presence of testosterone is essential for normal function and survival of the germ cells in seminiferous tubules (Sinha-Hikim and Swerdloff, 1999; Sofikitis et al., 2008). When the testicular environment can not support spermatogenesis, specific pathways leading to germ cell apoptosis are activated.
Abnormally accelerated apoptosis of germ cells may lead to an imbalance of cell proliferation and death, resulting in spermatogenic impairment (Kimura et al., 2003). Two major proapoptotic pathways have been defined in mammalian cells. One initiated at the cell surface via Apoptosis Stimulating Fragment (Fas) system and the second occurring in the mitochondria including Bax (Koji and Hishikawa, 2003). The Fas system is a receptor-ligand signaling system in which Fas ligand (FasL) binds to and activates the Fas receptor (Fas) to initiate a cascade of intracellular events that leads to the elimination of the Fas-bearing cells via apoptosis (Koji et al., 2001; Kavurma and Khachigian, 2003). The Fas system is involved in maintaining homeostasis in various systems. The Fas system in the testis has been identified as one paracrine signaling system by which Sertoli cells, expressing FasL, can initiate killing of Fas-expressing germ cells (Nagata, 1997). In present study, the effects of Dex, a widely used GC, on apoptosis and expression of FasL protein in the mause testicular germ cells were investigated.
MATERIALS AND METHODS
This study was conducted from 8/2007 to 5/2008 in Ahwaz Jundi-Shapour University of Medical Sciences, Faculty of Medicine, Ahwaz, Iran.
Chemicals: Primary antibody FasL secondary antibody biotinylated anti-mouse IgG (ABC Peroxidase Mouse IgG Staining Kit) and diaminobenzidine (DAB) were from Santa Cruz, USA. In situ cell death detection kit, for TdT-mediated dUTP nick end labeling (TUNEL) assessment, was the product of Roche Diagnostic, Germany. Dexamethasone was purchased from Daru Pakhsh Company, Iran. All other solvents and reagents were of the highest grade commercially available.
Animals: NMRI male mice weighing 25-30 g were kept in individual stainless steel cages under standardized conditions (constant temperature and humidity, 12 h light-dark cycle). They were fed with commercial chow and tap water ad lib.
Experimental protocol: The animals were randomly divided into 6 groups of 8 each. The experimental groups (E1-E3) received intrapritoneal injections of 4 (E1), 7 (E2) and 10 (E3) mg kg-1 Dex (dissolved in 0.9% saline) on 7 consecutive days. Control groups (C1-C3) were injected with 0.9% saline in equal volumes as for the experimental groups. Since TUNEL and immunohistochemical analysis showed no significant differences between three controls, all data were combined into one control group (C). One day after the last injection animals were sacrificed by decapitation under ether anesthesia. The testes were excised, fixed in formalin 10% for 48 h and embedded in paraffin wax. Five micrometer thick sections were prepared by using Leitz microtome for subsequent immunohistochemistry and deoxy-UTP-digoxigenin nick end labeling (TUNEL) studies.
Immunohistochemistry: Five-micron thick tissue sections were deparaffinized in xylol and hydrated in decreasing series of ethanol. Endogenous peroxidase activity was blocked by incubation in methanol containing 0.3% H2O2 for 15 min at room temperature. The sections were then treated with citrate buffer (pH = 6) for 15 min at 98°C as antigen retrieval. Then the sections incubated overnight at 4°C with primary antibodies, including the monoclonal antibody against FasL at 1/100 diluted in phosphate buffered saline (PBS; pH = 7.4) containing 10% normal goat serum (NGS). After washing twice with PBS the sections were incubated with secondary antibody biotinylated anti-mouse IgG at 1/100 for 50 min. Then the specimens were incubated with peroxidase-conjugated avidin-biotin for 30 min at room temperature. After washing, the sections were incubated with diaminobenzidine (DAB substrate) as chromogen and counterstained with haematoxyline. Three immunohistochemical sections from each animal were blindly assessed and staining intensity was estimated using a semiquantitative score, H-score, as earlierly described (Pallares et al., 2005; Ariel et al., 2001). The H-score (Histo-score) was calculated for each section by application of the following algorithm: H-score = ΣPi(i+1), where, i is the intensity of staining (0: no staining, 1: weak, 2: moderate and 3: strong) and Pi is the percentage of stained cells for each intensity (0 to 100%). For each mouse, at least 10 tubules/stage were used. The stages were identified according to the criteria proposed for paraffin sections. This method provides 12 stages of the spermatogenic cycle in mice. H-score assessment was repeated at least 3 times for each section by 3 workers.
TUNEL assay: The deoxyuridine nick-end labeling (TUNEL) assay for apoptotic cell detection was performed with the in situ cell death detection kit. Briefly, deparaffinized tissue sections were predigested with 20 μg mL-1 proteinase K for 20 min and incubated in PBS containing 3% H2O2 for 10 min to block the endogeous peroxidase activity. After incubating in 0.1% triton X-100 in 0.1% sodium citrate for two min on ice (4°C), the sections were incubated with the TUNEL reaction mixture, fluorescin-dUTP for 60 min at 37°C. The slides were then rinsed three times with PBS and incubated with secondary antifluorescine-POD-conjugate for 30 min. After washing three times in PBS, diaminobenzidine-H2O2 (DAB) chromogenic reaction was added to the sections. As a control for method specificity, the step using the TUNEL reaction mixture omitted in negative control section and nucleotide mixture in reaction buffer was used instead. A cell was considered TUNEL-positive when the nuclear staining was intense, dark brown and homogenous. Apoptotic index (AI) was calculated by dividing the number of TUNEL-positive germ cells in a randomly focused seminiferous tubule by the total number of germ cells in that tubule and the result was multiplied by 100 (Yu et al., 2001; Glicella et al., 2005). The AIs of 10 randomly selected tubules for each spermatogenic stage were evaluated and the mean AI of each case was calculated.
Statistical analysis: The data were analyzed using one-way ANOVA followed by LSD test and presented as the mean±SD. p<0.05 was considered significant.
RESULTS
Expression of FasL: C group showed weak immunoreactivity in cell membrane of spermatogonia at different stages of spermatogenic cycle (Fig. 1A). In E1 group spermatogonia showed strong immunostaining at stages VII-VIII (Fig. 1B). The pattern of FasL expression in other stages was similar to the C group. No detectable immunostaining was observed in primary spermatocytes or spermatids. In E2 group immunoreactivity was observed only at stages VII-VIII and IV-VI. Spermatogonia showed strong or moderate immunostaining and other germ cells were moderate or weak at stages IV-VI. In stages VII-VIII spermatogonia and primary spermatocytes were strongly stained and spermatids showed strong or moderate immunostaining (Fig. 1C). In E3 group all stages of spermatogenic cycle showed positive immunostaining. In stages I-III, strong staining in spermatogonia and weak immunostaining in primary spermatocyte and spermatids were observed. In stages IV-VI, spermatogonia and primary spermatocytes showed strong or moderate immunostaining and moderate or weak immunostaining was observed in spermatids. Stages VII-VIII showed the strongest immunostaining. Strong immunoreactivity was observed in spermatogonia and primary spermatocytes. Spermatids showed strong or moderate immunoreactivity. In stages IX-XII, moderate staining in spermatogonia and weak or moderate immunostaining in spermatids were observed (Fig. 1D). No expression of FasL was observed in Sertoli cells or Leydig cells in control and three experimental groups. The results of H-score assessments of FasL expression are reported in Fig. 2.
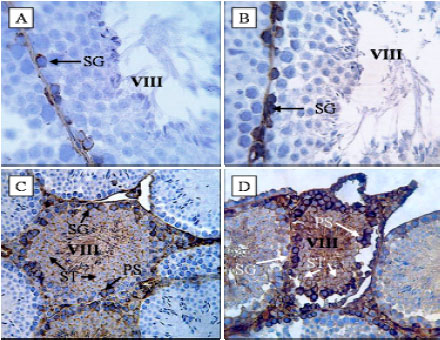 |
| Fig. 1: | FasL-immunodetection in control and experimental testes. A: Control testis; with moderate immunostaining in spermatogonia (x1000), B: E1 testis; showing strong imunoreactivity in spermatogonia (x1000), C: E2 testis; with moderate immunoreactivity only at stages VII-III (x400) and D: E3 testis; showing strong immunoreactivity at stages VII-VIII (x400). SG: Spermatogonia, PS: Primary spermatocytes, ST: Spermatid |
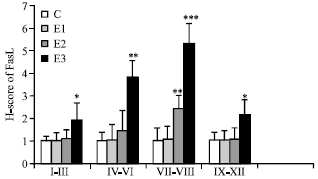 |
| Fig. 2: | H-score assessment of FasL expression in different stages of spermatogenic cycle in control and experimental groups. Values are expressed as Mean±SD for 8 mice, *p<0.05, **p<0.01, ***p<0.001, vs. C (control) |
Assessment of germ cell apoptosis: In C group, spermatogonia showed a low frequency of apoptosis in different stages (Fig. 3A). In E1 group, TUNEL-reactivity was observed in some of the spermatogonia and primary spermatocytes (Fig. 3B). There was no significant change in the AI of germ cells between E1 and C group (p>0.05). In E2 group, all types of germ cells showed TUNEL-positive reaction (Fig. 3C). The AIs of germ cells were significantly increased in all stages of spermatogenic cycle apart from stages I-III. In E3 group, all types of germ cells showed TUNEL-positive staining (Fig. 3D). The AI of germ cells were significantly increased in all stages of spermatogenic cycle, particularly in stages of VII-VIII. The results of the AIs of germ cells are reported in Fig. 4.
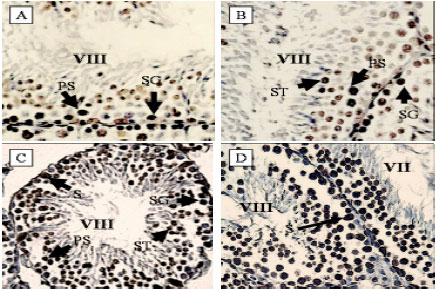 |
| Fig. 3: | TUNEL staining in control and experimental testes, A: Control testis; a few numbers of spermatogonia and primary spermatocytes showed TUNEL staining (x1000), B: E1 testis; a few numbers of spermatogonia, primary spermatocytes and spermatids showed TUNEL-positive reaction (x1000), C: E2 testis; TUNEL-positive reaction in spermatogonia, primary spermatocytes, spermatids and sertoli cells (x400) and D: E3 testis; the majority of germ cells and Sertoli cells showed TUNEL-positive reaction (x400). SG: Spermatogonia, PS: Primary spermatocytes, ST: Spermatid, S: Sertoli cell |
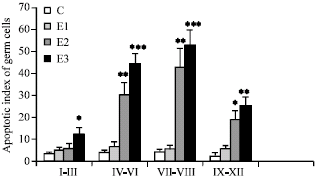 |
| Fig. 4: | Effects of Dex on apoptotic index (AI) of germ cells in different stages of spermatogenic cycle in mice. Values are expressed as Mean±SD for 8 mice, *p<0.05, **p<0.01, ***p<0.001 vs. C (control) |
Sertoli cells showed TUNEL-positive staining in both E2 and E3 groups. AIs of Sertoli cells were significantly increased in E2 (36.4±6.5% vs. 0%; p<0.01) and E3 (44.8±9.3% vs. 0%; p<0.01) groups. Leydig cells showed TUNEL-negative reaction in all groups.
DISCUSSION
The results of this study demonstrated that Dex dose-dependently increased FasL expression in testicular germ cells. Stages VII-VIII showed the most susceptible to apoptotic effects of Dex. Sinha et al. (1997) have previously reported that deprivation of gonadotropins and testosterone by GnRH antagonist treatment is followed by a stage-specific increase in germ cell apoptosis. There is abundant evidence suggesting that stages VII-VIII of the rat spermatogenic cycle exhibit the strongest levels of immunohistochemically detectable androgen receptors expression and are considered to be androgen dependent stages (Bremner et al., 1994; Nisrine et al., 2005).
It is possible that high susceptibility to apoptosis evidenced by FasL expression does not necessarily commit all FasL -immunopositive cells to the apoptotic cell death. For this reason we applied the TUNEL method. This method is based on the detection of oligonucleosomal DNA fragments which are characteristic of cells in the later stages of the apoptotic process (Walker et al., 1994). The measurement of apoptosis showed an increase in germ cell apoptosis after Dex treatment.
The reason responsible for the increased apoptosis induced by Dex in germ cells is not known. Although the endocrine control in testicular function is clear, the complexity of the intratesticular events highlights the importance of regulatory mechanisms and interactions. Intratesticular androgens, secreted by Leydig cells, play an important paracrine role in preventing germ cell degeneration (Tapanainen et al., 1993). Dex and other synthetic GCs exert a direct inhibitory effect on testosterone production by Leydig cells in vitro (Bernier et al., 1999). In addition to the inhibition of androgen biosynthesis, excessive exposure to GC in rodents initiate apoptosis in Leydig cells (Gao et al., 2002).
In contrast previous studies, which showed Dex induces apoptosis in rat Leydig cells (Gao et al., 2002, 2003), this study demonstrated that Leydig cells were TUNEL negative in Dex treated mice. Thus, the increase of FasL expression in testicular germ cells is probably related to the inhibition effect of Dex on androgen biosynthesis in Leydig cells. In present study androgen-independent stages of spermatogenic cycle also showed TUNEL-reactivity and expression of the proapototic protein. Thus, the increasing of apoptosis can not be exclusively due to hormonal influence.
Sertoli cells showed TUNEL-positive reaction and AI of Sertoli cells were significantly increased in E2 and E3 groups. Sertoli cells, the supportive cells in the seminiferous epithelium, orchestrate spermatogenesis by providing structural and nutritional support to germ cells (Boekelheide et al., 2000). In the rat, physiological apoptosis occurs continuously to limit the size of the germ cell population to numbers that can be adequately supported (Franca et al., 1993). Presence apoptosis in Sertoli cells, which demonstrated in this study, causes these cells loss their supporting action on germ cells and this may stimulate apoptotic signaling in germ cells. In this study, there was no detectable FasL imunoreactivity in Sertoli cells, while these cells showed TUNEL-positive staining. Additionally, in E2 group all stages of spermatogenic cycle showed TUNEL-positive reaction, while FasL expression was observed only in stages IV-VIII. These data suggest that other apoptotic factors are involved in Dex induced apoptosis in testicular tissue. Further experiments are needed to clarify the mechanisms of the effect of Dex on different apoptotic signaling pathways in testicular tissue.
ACKNOWLEDGMENTS
This research was a part of Ph.D project and supported by a grant (u-86044) from the research council of the Ahwaz Jundi-Shapour University of Medical Sciences.
REFERENCES
- Ariel, R., R. Hila, Y. Edward, X.U. Jing, H. Robin, S. Jean-Francois and F.C. Robert, 2001. Resveratrol, a nutural aryl hydrocarbone receptore antagonist, protects sperm from DNA damage and apoptosis caused by benzo(a) pyrene. Reprod. Toxicol., 15: 479-486.
CrossRef - Bernier, M., W. Gibb, R. Collu and J.R. Ducharme, 1999. Effect of glucocorticoids on testosterone production by porcine Leydig cells in primary culture. Can. J. Pharmacol., 62: 1166-1169.
PubMed - Boekelheide, K., S.L. Fleming, K.J. Johnson, S.R. Patel and H.A. Schoenfeld, 2000. Role of sertoli cells in injury-associated testicular germ cell apoptosis. Proc. Soc. Exp. Biol. Med., 225: 105-115.
CrossRefDirect Link - Bremner, W.J., M.R. Millar, R.M. Sharpe and P.T. Saunders, 1994. Immunohistochemical localization of androgen receptors in the rat testis: Evidence for stage-dependent expression and regulation by androgens. Endocrinology, 135: 1227-1234.
CrossRefPubMedDirect Link - Calogero, A.E., N. Burrello, A.M. Bosboom, M.R. Garofalo, R.F. Weber and R. D'Agata, 1999. Glucocorticoids inhibit gonadotropinreleasing hormone by acting directly at the hypothalamic level. J. Endocrinol. Invest., 22: 666-670.
PubMed - Dong, Q., A. Salva, C.M. Sottas, E. Niu, M. Holmes and M.P. Hardy, 2004. Rapid glucocorticoid mediation of suppressed testosterone biosynthesis in male mice subjected to stress. J. Androl., 25: 973-981.
PubMed - Gao, H.B., M.T. Tong, H.Y. Hu, H.Y. You, Q.S. Guo and R.S. Ge, 2002. Glucocorticoid induces apoptosis in rat Leydig cells. Endocrinology, 143: 130-138.
CrossRef - Gao, H.B., M.T. Tong, H.Y. Hu, H.Y. You, Q.S. Guo and R.S. Ge, 2003. Mechanism of glucocorticoid-induced Leydig cell apoptosis. Mol. Cell. Endocrinol., 199: 153-163.
CrossRef - Glicella, S., J. Ayesha, L. Dong, W. Hongquan, L. Jenny and J.W. Debra, 2005. Induction of apoptosis involving multiple pathways is a primary response to cyclin A1-deficiency in male meiosis. Dev. Dyn., 234: 114-123.
CrossRef - Hardy, M.P., H.B. Gao, Q. Dong, R. Ge and Q. Wang et al., 2005. Stress hormone and male reproductive function. Cell. Tissue Res., 322: 147-153.
CrossRef - Julia, C.B., 2006. Glucocorticoids: Exemplars of multi-tasking. Br. J. Pharmacol., 147: 258-268.
CrossRef - Kavurma, M.M. and L.M. Khachigian, 2003. Signaling and transcriptional control of fast ligand gene expression. Cell, Death Differ., 10: 36-44.
CrossRef - Kimura, M., N. Itoh, S. Takagi, T. Sasao, A. Takahashi, N. Masumori and T. Tsukamoto, 2003. Balanced of apoptosis and proliferation of germ cells related to spermatogenesis in aged men. J. Androl., 24: 185-191.
PubMed - Koji, T.Y. Hishikawa, H. Ando and Y. Nakanishi, 2001. Expression of fast and fast ligand in normal and ischemia-reperfusion testes: involvement of the fast system in the induction of germ cell apoptosis in the damaged mouse testis. Boil. Reprod., 64: 946-954.
Direct Link - Koji, T. and Y. Hishikawa, 2003. Germ cell apoptosis and its molecular trigger in mouse testes. Arch. Histol. Cytol., 66: 1-16.
CrossRef - El Chami, N., F. Ikhlef, K. Kaszas, S. Yakoub and E. Tabone et al., 2005. Androgen-dependent apoptosis in male germ cells is regulated through the proto-oncoprotein Cbl. J. Cell. Biol., 171: 651-661.
CrossRefPubMedDirect Link - Pallares, J., E. Bussaglia, J.L. Martínez-Guitarte, X. Dolcet and D. Llobet et al., 2005. Immunohistochemical analysis of PTEN in endometrial carcinoma: A tissue microarray study with a comparison of four commercial antibodies in correlation with molecular abnormalities. Mod. Pathol., 18: 719-727.
CrossRefDirect Link - Sinha, A.P., T.B. Rajavashisth, H.I. Sinha, Y. Leu and J.J. Bonavera et al., 1997. Significance of apoptosis in the temporal and stage-specific loss of germ cells in the adult rat after gonadotropin-relasing hormone antagonist treatment. Boil. Repord., 57: 1193-1201.
PubMed - Sinha-Hikim, A.P. and R.S. Swerdloff, 1999. Hormonal and genetic control of germ cell apoptosis in the testis. Rev. Reprod., 4: 38-47.
CrossRef - Sofikitis, N., N. Giotitsas, P. Tsounapi, D. Baltogiannis, D. Giannakis and N. Pardalidis, 2008. Hormonal regulation of spermatogenesis and spermiogenesis. J. Steroid Biochem. Mol. Biol., 109: 323-330.
CrossRefDirect Link - Tapanainen, J.S., J.L. Tilly, K.K. Vihko and A.J. Hsueh, 1993. Hormonal control of apoptotic cell death in the testis: gonadotropins and androgens as testicular cell survival factors. Mol. Endocrinol., 7: 643-650.
CrossRefPubMedDirect Link - Walker, P.R., V.M. Weaver, B. Lach and M. Sikorsa, 1994. Endonuclease activities associated with high molecular weight and internucleosomal DNA fragmentation in apoptosis. Exp. Cell. Res., 213: 100-106.
CrossRef - Yu, Q., J. Brain, P. Laneuville and D.G. Osmond, 2001. Suppressed apoptosis of pre-B cells in bone marrow of pre-leukemic p190bcr/ abl transgenic mice. Leukemia, 15: 819-827.
CrossRef