
YangYu -Xia
Agronomy College, Sichuan Agricultural University, Yaan, Sichuan 625014, China
Wu Wei
Agronomy College, Sichuan Agricultural University, Yaan, Sichuan 625014, China
ZhengYou- Liang
Agronomy College, Sichuan Agricultural University, Yaan, Sichuan 625014, China
Cai Qian-Rong
Agronomy College, Sichuan Agricultural University, Yaan, Sichuan 625014, China
Pakistan Journal of Biological Sciences
Year: 2008 | Volume: 11 | Issue: 13 | Page No.: 1662-1668







ABSTRACT
Prolamin and albumn variations of the storage proteins in 76 cultivated buckwheat accessions (55 accessions of Fagopyrum tataricum, 21 accessions of F. esculentum) from 7 countries were characterized by A-PAGE and SDS-PAGE, respectively, for the purpose of evaluating the genetic diversity of cultivated buckwheat at the level of proteins. A total of 18 prolamin bands were detected, among which 88.89 £¥bands were polymorphic. The number of albumn bands based on SDS-PAGE observed in accessions ranged from 4 to 10. Most intense bands were in the range of molecular weights from 29 to 97.2 kDa. The average of genetic similarity coefficient based on prolamin bands was 0.784 (in F. tataricum and F. esculentum were 0.892 and 0.681, respectively), while on prolamin and albumn bands was 0.742 (in F. tataricum and F. esculentum were 0.864 and 0.633, respectively). Accessions of F. tataricum and F. esculentum showed significant interspecific variation in the A-PAGE and SDS-PAGE profile of the storage proteins. The cluster analysis indicated that all the accessions could be divided into 3 groups and 3 subgroups. The genetic variations among cultivated buckwheat accessions were associated with their geographic origins in some degree.
PDF Abstract XML References Citation
How to cite this article
YangYu -Xia, Wu Wei, ZhengYou- Liang and Cai Qian-Rong, 2008. Genetic Diversity of Storage Proteins in Cultivated Buckwheat. Pakistan Journal of Biological Sciences, 11: 1662-1668.
DOI: 10.3923/pjbs.2008.1662.1668
URL: https://scialert.net/abstract/?doi=pjbs.2008.1662.1668
DOI: 10.3923/pjbs.2008.1662.1668
URL: https://scialert.net/abstract/?doi=pjbs.2008.1662.1668
INTRODUCTION
Buckwheat is a minor crop in the world but is an indispensable food in the temperate and hill regions of East Asia and Europe. It is a major staple food crop in the Indo-Himalayan region. Buckwheat is a multipurpose crop used for food, feed, medicine and manure. The tender shoots are used as a leafy vegetable, while the flower and green leaves are used for the extraction of rutin (Marshall and Pomeranz, 1982). Buckwheat flowers are a good source of honey. The genus Fagopyrum consists of about 19 species, some of which have been recently discovered (Ohnishi, 1998a, b). Among these species, only two types of buckwheat are used as food around the world: common buckwheat (Fagopyrum esculentum) and tartary buckwheat (Fagopyrum tataricum). Buckwheat is known as a valuable source of protein and its amino acid composition is nutritionally superior to that of cereal grains (Pomeranz and Robbins, 1972; Ikeda, 2002). Although, the digestibility of buckwheat protein is relatively low (Tomotake et al., 2006), it has many physiological activities which are potential for human health, such as hypocholesterolemic activity in rats, suppression in body fat, constipation, mammary carcinogenesis and colon carcinogenesis and in the formation of cholesterol gallstones in hamsters (Kayashita et al., 1996; Kayashita et al., 1997; Liu et al., 2001; Tomotake et al., 2000, 2001).
Assessment of genetic variation in a species is important for initiation of effective breeding programs because it provides the basis for tailoring desirable genotypes. SDS-PAGE profiles have been successfully used for analysis on diversity in buckwheat (Ohnishi, 2000; Morita et al., 2006; Tang, 2007) Even though much work has been done on the analysis of genetic variations between different species of the genus Fagopyrum using isozyme profiling (Huh and Huh, 2000), RAPD ( Murai and Ohnishi, 1996; Kump and Javornik, 2002), AFLP (Tsuji and Ohnishi, 2001; Iwata et al., 2005), not much information is available on inter- as well as intra-specific variations in storage protein variations in cultivated buckwheat. While the storage proteins have the advantage of reflecting numerous variations as they belong to very polymorphic multigenic families (Doll and Brown, 1979). In addition, storage proteins are also reliable bio-chemical makers for the evaluation of genetic diversity (Branlard et al., 1989).
In this study, the genetic variations of storage protein in 76 cultivated buckwheat accessions, which were collected from various areas, were investigated by A-PAGE and SDS-PAGE methods, respectively. The objective of this study was to evaluate the genetic diversity of cultivated buckwheat accessions at the level of proteins with reference to the improvement of buckwheat.
MATERIALS AND METHODS
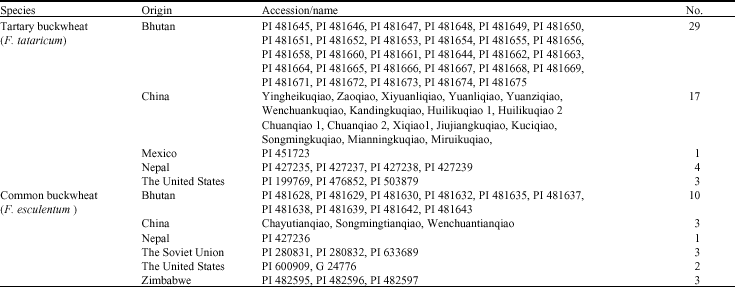
Plant materials: Seventy-six cultivated buckwheat accessions from 7 different countries were analyzed (Table 1), including fifty-five accessions of F. tataricum, twenty-one accessions of F. esculentum. Among them, ‘PI’ accessions were kindly provided by Dr. Croston (American National Plant Germplasm System), the others were collected from Sichuan, Yuannan and Tibet Province of China in 2007.
A-PAGE: Prolamin protein were extracted from single seeds with a solution of 25% (V/V) α-ethanol and 0.05% (W/V) methyl green and fractionated by a standard acid-polyacrylamide-gel electrophoresis (A-PAGE) at pH 3.1 (Draper, 1987). PI 481635 and Chayutianqiao was used as reference in A-PAGE analysis for their bands were more and clear distinct relatively than compared with the others.
SDS-PAGE: Glutein proteins were extracted from grains of different accessions of buckwheat with 50 mmol L-1 Tris-Cl buffer (pH 6.8) containing 100 mmol L-1 NaCl, 10 mmol L-1 EDTA, 100 mmol L-1 Glycine, 10% SDS and 1 mmol L-1 phenyl methyl sulphonyl fluoride (PMSF). The crude homogenate was kept for 45 min at 4°C and centrifuged at 10,000 rpm for 15 min. The extracted proteins were recovered as clear supernatant. SDS-PAGE of the extracted proteins was carried out on 12% isolation gel and 3.7% concentration gel under reducing conditions following the method of Laemmili (1970). For standards we used wide range protein markers (M.W. 6500~200,000, which were synthesized by Shanghai Sangon Biotech Ltd., China). Electrophoresis was carried out at a constant current of 20 mA for 3-4 h. The gels were stained for 3 h in 0.25% (w/v) coomassie brilliant blue R-250 followed by destaining in methanol:water:glacial acetic acid (4:5.3:0.7). Protein bands were visualized in a transilluminator under white light.
Statistic analysis: Evaluation of variation in the storage proteins was performed by calculating the individual band frequency for each accession. Polymorphism was scored for the presence (1) or absence (0) of bands. The data matrix was then used to calculate Genetic Similarity (GS) values according to Nei and Li`s (1979) method:
| GS = 2Nij/(Ni+Nj) |
where, Nij is the number of prolamin and albumn bands in common between accessions i and j and Ni and Nj are the total number of prolamin and albumn bands observed for accessions i and j, respectively. Based on the GS matrix, a dendrogram showing the genetic relationships between accessions was constructed using the Unweighted Pair-Group Method with Arithmetic Average (UPGMA) through the computer software NTSYS-pc version 2.10 (Rohlf, 2000).
| Table 1: | Accessions of cultivated buckwheat used in the study |
 | |
RESULTS
Prolamin variations: A total of 18 prolamin bands were detected in 76 accessions of cultivated buckwheat (Fig. 1), among which 88.89% were polymorphic. Each accession had 6~12 bands with the mean of 9.46. Thirty-two prolamin patterns were identified. The prolamin patterns in 44 accessions of F. tataricum (such as PI 481645, PI 481675, Kangdingkuqiao and Huilikuqiao, etc.) were identical. Thus, these accessions could not be distinguished by A-PAGE.
Based on the prolamin bands, the GS in cultivated buckwheat accessions ranged from 0.375 to 1.000 with the mean of 0.784, indicating that relatively higher genetic diversity existed in cultivated buckwheat. The GS value between PI 451723 and PI 633689 was the lowest. It was showed that the GS values ranged from 0.667 to 1.000 with an average of 0.892 in F. tataricum, the GS value between Huilikuqiao 1 and PI 476852 was the lowest, while in F. esculentum was from 0.400 (between Wenchuantianqiao and PI 482595) to 0.947 (between PI 280831 and G 24776) with an average of 0.681. And the average GS value between F. tataricum and F. esculentum was 0.674. This indicated that distinct quantitative variations were observed between F. tataricum and F. Esculentum and the genetic variation within F. esculentum was more plentiful than in F. tataricum.
Albumn variations: A total of 15 albumn bands were detected in the tested materials, among which 93.33% were polymorphic (Fig. 2). Each accession had 4~10 bands with the mean of 7.76. Thirty-one albumn patterns were identified from 76 accessions. The albumn patterns in 45 accessions were identical.
Most intense bands were in the range of molecular weights from 29 to 97.2 kDa. Distinct quantitative variations were observed in the SDS-PAGE profile of storage proteins of F. tataricum and F. esculentum. Significant intraspecific variations were detected in the storage protein profile of different accessions of F. esculentum. This could be ascribed to a high degree of grain protein polymorphism in F. esculentum. Most of these variations were observed in the protein bands ranging in size between 29 and 66.4 kDa. Interestingly, most of the protein bands in the 26~54 kDa category belong to the legumin type family of seed proteins (Rout and Chrungoo, 1996).While accessions of F. tataricum and F. esculentum did not show any significant interspecific variations in the SDS-PAGE profile of storage proteins.
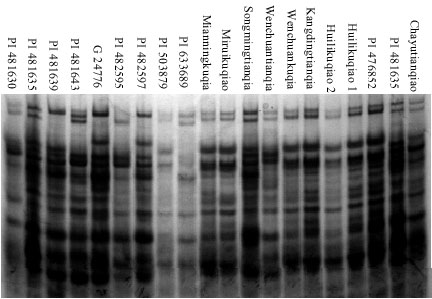 | |
| Fig. 1: | Prolamin patterns of some cultivated buckwheat |
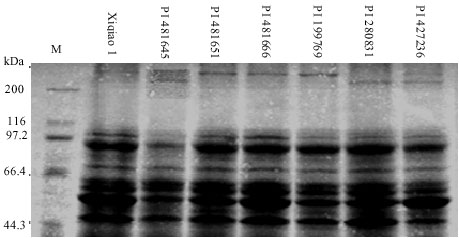 | |
| Fig. 2: | SDS-PAGE patterns of albumn proteins in some accessions of cultivated buckwheat |
Prolamin and albumn variations: Based on the prolamin and albumn bands, the GS in cultivated buckwheat accessions ranged from 0.303 to 1.000 with the mean of 0.742. The GS value between PI 427238 and PI 482595 was the lowest. It was showed that the GS values ranged from 0.563 to 1.000 with an average of 0.864 in F. tataricum, the GS value between PI 503879 and PI 481669 was the lowest, while in F. esculentum were from 0.412 (between PI 280832 and Chayutianqiao) to 0.972 (between PI 280831 and G 24776) with an average of 0.633. Significant intraspecific variations were detected in the storage protein profile of different accessions of F. esculentum. Interspecific variations were existed between F. tataricum and F. esculentum.
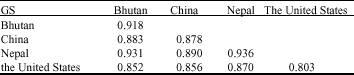
It was showed that the average GS value of F. tataricum accessions within one country or between two countries were all higher than that among all the accessions (0.742), while opposite to F. esculentum. The average GS value within the accessions of F. tataricum from the United States was the lowest, while in Nepal was the highest. The average GS between the United States and other countries were all lower relatively, which inferred that the genetic relationships among the United States were far, relatively. The result was similar to prolamin variations (Table 2).
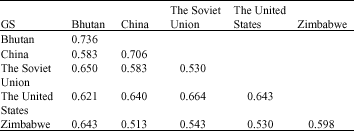
The result was showed that the average GS value of F. esculentum accessions within one country or between two countries were all lower than that among all the accessions (Table 3). The average GS value within Bhutan and China were quite similar (0.736, 0.706, respectively), within the Soviet Union was the lowest. The average GS value between the Untied States and the Soviet Union was 0.664, which was the highest between two countries, the lowest was between China and Zimbabwe. It inferred that the genetic variations among the F. esculentum accessions from different countries were higher, the genetic variations within the Soviet Union accessions was relatively higher than other countries accessions of F. esculentum.
| Table 2: | Average value of GS within and between countries based on prolamin and albumn bands of F. tataricum |
 | |
| Table 3: | Average value of GS within and between countries based on prolamin and albumn bands of F. esculentum |
 | |
Based on the prolamin and albumn bands, 76 cultivated buckwheat accessions could be divided into three major groups, too. Of which group 2 included 11 accessions from Bhutan, the Soviet Union and Zimbabwe and just one accession from Zimbabwe (i.e., PI 452597) consist of group (Fig. 3). Group 1 could be further divided into three subgroups. All accessions of F. tataricum and six F. esculentum (such as G 24776, Chayutianqiao, Songmingtianqiao, etc.) were clustered into subgroup Ia. Two accessions of F. esculentum (PI 633689 and PI 600909) which from the Soviet Union and the United States were clustered into subgroup Ib. Subgroup Ic were constituted by PI 481628 from Bhutan. Most of accessions come from the same country were clustered into one group. These results suggested that most of the accessions with close geographic origins had the tendency to cluster together indicating that the their geographic origins variations among cultivated buckwheat were associated with their geographic origins.
DISCUSSION
It has been proposed that prolamin pattern could provide valuable and exact information for the identification and utilization of germplasm. In this study, relatively higher genetic diversity of prolamin was found in cultivated buckwheat, especially in F. esculentum. Branlard et al. (1989) concluded that the number of diverse patterns was not generally very high for the wheats of a given country. Some results showed that prolamin patterns could reflect the systematic relationships among the type species of four genera in Triticeae. In this study, it was found that the prolamin variations among cultivated buckwheat were associated with their geographic origin. It was likely explained by genetic chemotaxis resulting from the geographical environment or incidental selection of other traits. The prolamin and albumn patterns in most of F. tataricum were identical and caused these accessions could not be distinguished by A-PAGE and SDS-PAGE. The reason might be that F. tataricum is self pollination crop and its self-cross rate is larger than 95%. Furthermore, the locus detected by A-PAGE and SDS-PAGE were few and it couldn`t revealed many information. Moreover, the geographical distribution of some accessions was closer and germplasm resources` exchange could caused fewer genetic variations. For example, in Fig. 3, most of accessions from China, Nepal and Bhutan were clustered together for these counties are bound on each other.
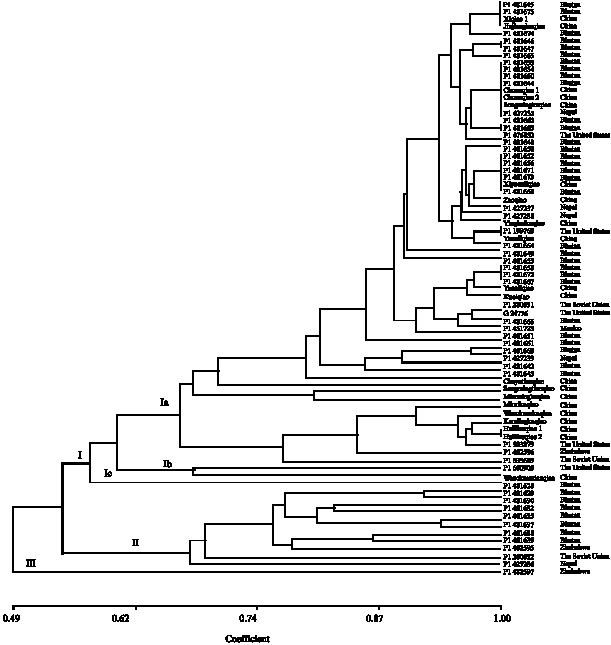 | |
| Fig. 3: | Cluster analysis of 76 accessions from cultivated buckwheat based on prolamin and albumn bands |
Particular accessions of F. esculentum were clustered with F. tataricum. Similar observations have also been made by Yasui and Ohnishi (1998a, b) who compared the nucleotide sequence of the rbcL-accD region of cpDNA in the genus. These results are in agreement with the molecular systematic studies of Ohnishi and Matsuoka (1996) and Ohsako and Ohnishi (1998, 2000) on Fagopyrum. Present results indicate that F. esculentum exhibited very close relationships with F. tataricum. These observations are also brought out by a combined dendrogram generated on the basis variations A-PAGE and SDS-PAGE profiles. These observations are in conformity with earlier findings reported on species relationships in Fagopyrum using different marker approaches (Kishima et al., 1995).
In this study, we detected most intense bands were in the range of molecular weights from 29 to 66.4 kDa. Two most intense bands were in the range of MWs from 50 to 60 kDa. These proteins have been shown not to be associated with allergy (Urisu et al., 1995, Nair and Adachi, 1999). Similar observations have also been made by Licen et al. (2005) who indicated six intense bands were detected from buckwheat endosperm extracts.
ACKNOWLEDGMENTS
The authors thank Dr. Croston of American Germplasm Resources Information Network (GRIN) for kindly providing fifty-six accessions used in this experiment. And the experiment was accomplished in the lab of Gene Resource and Molecular Breeding in Sichuan Agricultural University. This work is financial supported by the Youth Science and Technology Foundation of Sichuan Province in China.
REFERENCES
- Branlard, G., J.C. Aulran and P. Monneveux, 1989. High molecular weight glutenin subunit in durum wheat (T. durum). Theor. Applied Genet., 78: 353-358.
CrossRef - Draper, S.R., 1987. 1987 ISTA and 1987 Variety committee: Report of the working group for biochemical tests for cultivar identification 1983-1986. Seed Sci. Technol., 15: 431-434.
Direct Link - Doll, H. and A.D.H. Brown, 1979. Hordein variation in wild (Hordeum spontaneum) and cultivated (H. vulgare) barley. Can. J. Genet. Cytol., 21: 291-404.
Direct Link - Huh, H.W. and M.K. Huh, 2000. Allozyme variation and population structure of common buckwheat, Fagopyrum esculentum, in Korea. Plant Germplasm Institute, 17: 21-27.
Direct Link - Ikeda, K., 2002. Buckwheat: Composition, chemistry and processing. Adv. Food Nutr. Res., 44: 395-434.
PubMed - Iwata, H., K. Imon, Y. Tsumura and R. Ohsawa, 2005. Genetic diversity among Japanese indigenous common buckwheat (Fagopyrum esculentum) cultivars as determined from amplified fragment length polymorphism and simple sequence repeat markers and quantitative agronomic traits. Genome, 48: 367-377.
CrossRef - Kayashita, J., I. Shimaoka, M. Nakajoh and N. Kato, 1996. Feeding of buckwheat protein extract reduces hepatic triglyceride concentration, adipose tissue weight and hepatic lipogenesis in rats. J. Nutr. Biochem., 7: 555-559.
Direct Link - Kayashita, J., I. Shimaoka , M. Nakajoh, M. Yamazaki and N. Kato, 1997. Hypocholesterolemic effect of buckwheat protein extract in rats fed cholesterol enriched diets. Nutr. Res., 15: 691-698.
CrossRefDirect Link - Kishima, Y.K., O.K. Mizukami, T. Mikami and T. Adachi, 1995. Chloroplast DNA analysis in buckwheat species:phylogenetic relationships, origin of reproductive systems and extended inverted repeats. Plant Sci, 108: 173-179.
CrossRef - Kump, B. and B. Javornik, 2002. Genetic diversity and relationships among cultivated and wild accessions of tartary buckwheat (Fagopyrum tataricum Gaertn.) as revealed by RAPD markers. Genet. Resour. Crop Evol., 49: 565-572.
CrossRefDirect Link - Laemmli, U.K., 1970. Cleavage of structural proteins during the assembly of the head of bacteriophage T4. Nature, 227: 680-685.
CrossRefDirect Link - Ličen, M. and I. Kreft, 2005. Buckwheat (Fagopyrum esculentum Moench) low molecular weight seed proteins are restricted to the embryo and are not detectable in the endosperm. Plant Physiol. Biochem., 43: 862-865.
CrossRef - Liu, Z., W. Ishikawa, X. Huang, H. Tomotake, J. Kayashita and H. Watanabe, 2001. A buckwheat protein product suppresses 1,2-dimethylhydrazine-induced colon carcinogenesis in rats by reducing cell proliferation. J. Nutr., 131: 1850-1853.
PubMedDirect Link - Morita, N., T. Maeda, R. Sai, K. Miyake, H. Yoshioka, A. Urisu, T. Adachi, 2006. Studies on distribution of protein and allergen in graded flours prepared from whole buckwheat grains. Food Res. Int., 39: 782-790.
CrossRef - Murai M. and O. Ohnishi, 1996. Population genetics of cultivated common buckwheat, Fagopyrum esculentum Moench. X. Diffusion routes revealed by RAPD markers. Genes. Genetic. Syst., 71: 211-218.
PubMed - Nair, A. and T. Adachi, 1999. Immunodetection and characterization of allergenic proteins in common buckwheat (Fagopyrum esculentum). Plant Biotechnol., 16: 219-224.
Direct Link - Nei, M. and W.H. Li, 1979. Mathematical model for studying genetic variation in terms of restriction endonucleases. Proc. Natl. Acad. Sci. USA., 76: 5269-5273.
CrossRefPubMedDirect Link - Ohnishi, O., 1998. Search of the wild ancestors of buckwheat I Description of new Fagopyrum species and their distribution in China. Fagopyrum, 15: 18-28.
Direct Link - Ohnishi, O., 1998. Search of the wild ancestors of buckwheat III. The wild ancestor of cultivated common buckwheat and of tatary buckwheat. Econ. Bot., 52: 123-133.
CrossRef - Ohnishi O., 2000. Geographical distribution of allozymes in natural populations of wild tartary buckwheat. Fagopyrum, 17: 29-34.
Direct Link - Ohnishi, T. and Y. Matsuoka, 1996. Search for the wild ancestor of Buckwheat. II Taxonomy of the Fagopyrum (Polygonaceae) species based on morphology, isozyme and chloroplast DNA variability. Genes. Genet. Syst., 71: 383-390.
CrossRefDirect Link - Ohsako, T. and O. Ohnishi, 1998. New Fagopyrum species revealed by morphological and molecular analyses. Genes. Genetic. Syst., 73: 85-94.
CrossRefDirect Link - Ohsako, T. and O. Ohnishi, 2000. Intra and inter-specific phylogeny of the wild Fagopyrum (Polygonaceae) species based on nucleotide sequences of non coding regions of chloroplast DNA. Am. J. Bot., 87: 573-582.
PubMed - Pomeranz, Y. and G.S. Robbins, 1972. Amino acid composition of buckwheat. J. Agric. Food Chem., 20: 270-274.
Direct Link - Rohlf, F.J., 1993. NTSYS-pc: Numerical Taxonomy and Multivariate Analysis System. Version 2.20, Exeter Software, New York, ISBN: 0-925031-30-5, pp: 1-38.
Direct Link - Rout, M.K. and N.K. Chrungoo, 1996. Partial characterization of the lysine rich 13S globulin from buckwheat: Its antigenic homology with seed proteins of some other crops. Biochem. Mol. Biol. Int., 40: 587-595.
PubMed - Rout. A. and N.K. Chrungoo 2007. Genetic variation and species relationships in Himalayan buckwheats as revealed by SDS-PAGE of endosperm proteins extracted from single seeds and RAPD based DNA fingerprints. Genet. Resour. Crop Evol., 54: 767-777.
CrossRefDirect Link - Tang, C.H., 2007. Functional properties and in vitro digestibility of buckwheat protein products: Influence of processing. J. Food Eng., 82: 568-576.
CrossRefDirect Link - Tomotake, H., I. Shimaoka, J. Kayashita, F. Yokoyama, M. Nakajoh and N. Kato, 2000. A buckwheat protein product suppresser gallstone formation and plasma cholesterol more strongly than soy protein isolate in hamster. J. Nutr., 130: 1670-1674.
Direct Link - Tomotake, H., I. Shimaoka, J. Kayashita, F. Yokoyama, M. Nakajoh and N. Kato, 2001. Stronger suppression of plasma cholesterol and enhancement of the fecal excretion of steroids by a buckwheat protein product than by a soy protein isolate in rats fed on a cholesterol-free diet. Biosci. Biotechnol. Biochem., 65: 1412-1414.
CrossRefPubMed - Tomotake, H., N. Yamamoto, N. Yanaka, H. Ohinata, R. Yamazaki, J. Kayashita and N. Kato, 2006. High protein buckwheat flour suppresses hypercholesterolemia in rats and gallstone formation in mice by hypercholesterolemic diet and body fat in rats because of its low protein digestibility. Nutrition, 22: 166-173.
CrossRefPubMedDirect Link - Urisu, A., Y. Kondo, Y. Morita, E. Yagi, M. Tsuruta, T. Yasaki, K. Yamada, H. Kuzuya, I. M. Suzuk , K. Titani and K. Kurosawa, 1995. Isolation and characterization of major allergen in buckwheat seed. In: Current Advances in Buckwheat Research. Matsumoto, Shinshu University Press, Japan, ISBN: 1672-1977 pp: 965-974.
- Yasui, Y. and O. Ohnishi, 1998. Phylogenetic relationships among Fagopyrum species revealed by the nucleotide sequences of the ITS region of the nuclear rRNA gene. Genes. Genetic. Syst., 73: 201-210.
Direct Link - Yasui, Y. and O. Ohnishi, 1998. Interspecific relationships in Fagopyrum (Polygonaceae) revealed by the nucleotide sequences of the rbcL and accD genes and their intergenic region. Am. J. Bot., 85: 1134-1142.
Direct Link