
P. Tajik
Department of Clinical Sciences,
Faculty of Veterinary Medicine, University of Tehran, P.O. Box 14155-6453, Tehran, Iran
P. Mirshokraee
Department of Clinical Sciences,
Faculty of Veterinary Medicine, University of Tehran, P.O. Box 14155-6453, Tehran, Iran
A. Khosravi
Department of Microbiology, Faculty of Veterinary Medicine, University of Tehran, P.O. Box 14155-6453, Tehran, Iran
Pakistan Journal of Biological Sciences
Year: 2007 | Volume: 10 | Issue: 24 | Page No.: 4500-4504







ABSTRACT
The present study was carried to observe the effect of aflatoxin on ovine sperm in vitro. Ram epididymal and ejaculatory sperm cells were added into media containing different concentrations of aflatoxin B1. When ram epididymal sperm were put in different concentrations of aflatoxin, one-hour post incubation in control group, 81.25% of sperm cells were alive of which, 82.88% were motile. Significantly (p< 0.05) higher than medium with 31.25 ppb and higher concentration. The lowest motility (15.93%) was observed in 62.5 ppb aflatoxin. Sperm viability did not significantly changed after 2nd and 3rd h incubation but significantly decreased in 4th and 5th h post incubation. However, motility was not altered in the 2nd, 3rd, 4th and 5th h post incubation. In ejaculatory sperm, one hour post-incubation, in control group, 83.24% of sperm were alive of which 93.98% were motile, significantly (p< 0.05) higher than motility in medium with 7.81 ppb and higher concentrations of aflatoxin but not different with 1.96 ppb aflatoxin. In 62.5 ppb aflatoxin, the lowest motility was observed (18.9%). Sperm motility patterns for both epididymal and ejaculatory sperm cells were different after incubation in different concentrations of aflatoxin. The results of the present experiment showed that aflatoxin could decrease sperm motility obtained from ejaculation or epididymis.
PDF Abstract XML References Citation
How to cite this article
P. Tajik, P. Mirshokraee and A. Khosravi, 2007. Effects of Different Concentrations of Aflatoxin B on Ram Epididymal and Ejaculatory Sperm Viability and Motility in vitro. Pakistan Journal of Biological Sciences, 10: 4500-4504.
DOI: 10.3923/pjbs.2007.4500.4504
URL: https://scialert.net/abstract/?doi=pjbs.2007.4500.4504
DOI: 10.3923/pjbs.2007.4500.4504
URL: https://scialert.net/abstract/?doi=pjbs.2007.4500.4504
INTRODUCTION
Mycotoxins, secondary metabolites of fungus are toxic for animals and human (D`Mello and Macdonald, 1997). These substances are found in animal feed and soon or late will affect different parts of body. There is not so many works indicating different effects of mycotoxins on reproductive tract. Zeralenone is one of the toxins, which is studied in relation to reproduction. It is secreted by the fungi of Fusarium family and has estrogenic effect. It has been suggested to alter estrous cycle in pigs. It also had reduced pregnancy rate and increased vaginal prolapse in this species (Osweiler, 1986). The effect of Zeralenone is also studied on oocytes maturation and its contra effect with estradiol 17 β on granulosa cell development in cow (Minervini et al., 2001). Another detrimental mycotoxin, which has been traced in animal origin food in Iran, is aflatoxin. Some species of Aspergillus which produce this toxin are; A. flavous, A. pominus and A. parasiticus. Aflatoxin is strongly toxic, carcinogenic, tratogenic, mutagenic and immunosuppressive (Puschner, 2002). It has been detected in the milk of animals ate it (D`Mello and Macdonald, 1997) and so it is suspicious to be transferred to human by food products. There are many studies concerning the effect of aflatoxin on reproduction of rats and rabbits. Esteroidogenic and spermatogenic potentials, sperm maturation and storage in the male rat after acute treatment with aflatoxin. He has also studied the Sperm production rates, sperm physiology and fertility in rats chronically treated with sub-lethal doses of aflatoxin (Egbunike et al., 1980; Egbunike, 1982, 1985). Hafez et al. (1983) have studied the effect of dietary aflatoxins on adult fertile male and female rabbits at various reproductive conditions. They showed that feeding rabbits with aflatoxin during 7 days significantly decreased conception rates in this species. Picha et al. (1986) showed that aflatoxin is present in the seminal plasma of boars. They have also postulated its relation to sperm production. After feeding ocharatoxin, Solti et al. (1999) have analyzed the serum and seminal plasma of boars and could find it in plasma. The effects of dietary concentration of aflatoxin with or without vitamin C on meiotic chromosomes, sperm head morphology and sperm count in mice has been studied (Sinha and Prasad, 1990; Sinha and Bose, 1992). Ibeh et al. (1994) have studied the correlation of human serum aflatoxin and infertility. Ibeh and Saxena (1998) studied the effect of aflatoxin on the histopathology and some enzymes in testes of rats. A significant decrease in semen quality, sperm concentration and motility but no change in pH and fructose of rabbits orally received aflatoxin (ElZahar et al., 1996). Uriah et al. (2001) have studied the impact of aflatoxin on human reproduction. In their observation in Nigeria, they noticed that the concentrations of aflatoxin were higher in infertile men comparing to the fertile ones. Alterations of plasma testosterone, ejaculation volume, sperm concentration and motility as well as semen fructose were significant in rabbits treated with aflatoxin (Salem et al., 2001). Reported the reduction of testosterone production in chronic aflatoxicosis in mice (Verma and Nair, 2002). Epididymal sperm, on the other hand has been widely retrieved from the men with obstructive azoospermia and insemination with them resulted to pregnancies (Patrizio, 2000). Agnes and Akbarsha (2003) showed that the fertility of the treated mice with aflatoxin B1 was reduced drastically. Sperm concentration in the epididymis and sperm motility decreased whereas sperm abnormalities increased. When bulls feed with contaminated food sperm motility was greatly reduced and there were a high percentage of sperm abnormalities and plasma testosterone and oesteradiol-17 β concentrations were elevated (Lange et al., 1998). Hafez et al. (1982) showed that three buffalo bulls’ semen that feed with aflatoxin B and G during two weeks had marked decrease in the live percentage of sperm and high increase in the sperm abnormalities.
MATERIALS AND METHODS
Sperm preparation: Ram testicles were prepared from a local slaughterhouse in Tehran suburb and carried to the laboratory in a cool environment (°C) because it has been reported that sperm cells will remain motile when testicles are transported and kept in cool place (James et al., 2002; Yu and Leibo, 2002). Cauda epididymal sperm cells were obtained using a sterile scalpel, the parietal tunic was removed leaving the tail of epididymis exposed as done by Blash et al. (2000) and used for motility assess because it has been show to have the maximum motility comparing the other parts of epididymis (Osweiler, 1986). Sperm cells from ejaculated semen and sperm isolated from cauda epididymis were diluted in BO medium. This medium consisted of 112.0 mM-NaCl, 4.02 mM-KCl, 2.25 mM-CaCl2, 0.83 mM-NaH2PO4, 0.52 mM-MgCl2, 37.0 mM-NaHCO3, 13.9 mM-glucose, 1.25 mM-sodium pyruvate, 31 mg mL-1 potassium penicillin G and contained 0.1% polyvinyl alcohol (PVA) (W/v) because in our previous study (Tajik et al., 1994) this concentration of PVA could support frozen-thawed bovine sperm motility in BO medium. This medium had the osmolarity of almost 300 mOsm, which is shown to have a less detrimental effect on bovine epididymal spermatozoa than hypo and hyper osmotic versions (De Pauw et al., 2003). Ejaculation was preformed 2 days a week because Kaya et al. (2002) have shown that ejaculation frequency could significantly decrease sperm motility in rams. For motility scoring, sperm cells were diluted to have a final concentration of 2-4x106 cells. Sperm motility was assessed under the x20 magnification of a light microscope. Totally 10 healthy Chall rams were used for ejaculation and 3 replications were carried. For epididymal sperm 20 testicles (10 rams) were used and data were pooled for them. Sperm cells stained by eosin nigrosin were observed under a light microscope X 1000 to access sperm viability during study. Experiment was carried during one year (12 month) started in September 2005 and finished the next year. Similar results were observed during the experiment time, so the data were pooled.
Statistical analysis: Motility of epididymal and ejaculatory sperm cells in different concentrations of aflatoxin-a, were analyzed by ANOVA. When the analysis revealed a significant effect, the values were compared by Duncan’s multiple range test.
RESULTS
Epididymal sperm viability is shown in Table 1. One hour after incubation of sperm cells in medium without aflatoxin, in control group, 81.2% of sperm cells were live, which was significantly (p<0.05) higher than live sperm cells in medium supplemented with 7.81, 31.25 and 62.6 aflatoxin (with 72.92, 71.8 and 66.72% live sperm cells, respectively) but not different with that of medium supplemented with 1.96 ppb aflatoxin where sperm motility was 78.84%. In second hour of incubation the proportion of live sperm was significantly altered in all groups. The proportion of live sperm was not altered in control group after 3 h incubation; however it was reduced dose dependently in other groups.
Table 2 shows the proportion of live ejaculatory sperm in different concentrations of aflatoxin B.
| Table 1: | Viability of ram epididymal sperm after incubation in different concentrations of aflatoxin |
 | |
| a-c: Values in columns with different superscripts are significantly different (p<0.05) | |
| Table 2: | Viability of ram ejaculatory sperm after incubation in different concentrations of aflatoxin |
 | |
| a-d: Values in columns with different superscripts are significantly different (p<0.05) | |
| Table 3: | Motility of ram epididymal sperm after incubation in different concentrations of aflatoxin |
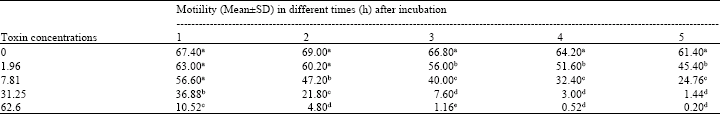 | |
| a-d: Values in columns with different superscripts are significantly different (p<0.05) | |
One hour post-incubation in control group (without aflatoxin), 83.24% of sperm cells were live, which was significantly (p<0.05) higher than proportion of live sperms in medium supplemented with 7.81, 31.25 and 62.6 ppb aflatoxin (with the proportion of 72.48, 69.6 and 63.63% live sperms, respectively) but not different with medium supplemented with 1.96 ppb aflatoxin where 80.84% of sperm cells were alive. In second hour of incubation the highest live percent was observed in control (80.44%), which was not significantly different with 1.96 (76.36%). However, it was decreased in 7.81, 31.25 and 62.5 ppb aflatoxin which viability was 70.68, 63.2 and 57.92%, respectively. As in the epididymal sperm, live percent did not alter in control group after 3 h post-incubation.
To define the real proportion of motility in the present experiment, sperm motility was calculated from the live sperm cell and was called motility ratio. One hour after incubation of sperm cells in medium without aflatoxin, in control group, 82.89% of sperm cells were motile, which was significantly (p<0.05) higher than motility in medium supplemented with 31.25 and 62.6 aflatoxin (with 51.87 and 15.93% motility, respectively) but not different with medium supplemented with 1.96 and 7.81 ppb aflatoxin where sperm motility was 80.03 and 77.83%, respectively. In second hour of incubation motility was not significantly changed in control and 1.96 ppb aflatoxin. However, it was decreased in 7.81, 31.25 and 62.5 ppb aflatoxin which motility was 66.66, 34.63 and 8.27%, respectively (Table 3). Sperm motility was not altered in control group after 3 h post-incubation; however it was reduced dose dependently in other groups. Indeed almost all sperm cells were immotile in 62.6 ppb aflatoxin supplemented medium after 3rd h (2.15%), which was significantly lower than other groups (p<0.05).
Sperm motility patterns were also different after incubation in different concentrations of aflatoxin. One hour post-incubation in control group (without aflatoxin), about 93% of sperm cells were motile, which was significantly (p<0.05) higher than motility in medium supplemented with 31.25 and 62.6 ppb aflatoxin (with motility ration of 52.09 and 18.09%, respectively) but not different with medium supplemented with 1.96 and 7.81 ppb aflatoxin where sperm motility ratio of 92.76 and 85.04, respectively. In second hour of incubation the highest motility ration was observed in control (92.93%), which was not significantly different with 1.96 (89%). However, it was decreased in 7.81, 31.25 and 62.5 ppb aflatoxin which motility was 74.67, 36.09 and 10.15%, respectively. As in the epididymal sperm, motility ratio did not alter in control group after 3 h post-incubation (with motility ratio of 90.15%) (Table 4). It was significantly lower (p<0.05) in 7.81-62.6 ppb dose dependently. Motility ratio was maintained at highest proportion during 4th and 5th h post incubation in control and lowest in 62.6 ppb aflatoxin.
| Table 4: | Motility of ram ejaculatoryl sperm after incubation in different concentrations of aflatoxin |
 | |
| a-d: Values in columns with different superscripts are significantly different (p<0.05) | |
DISCUSSION
Agnes and Akbarsha (2003) have reported the effects of aflatoxin B1 I the albino mouse epididymal sperm concentration. With the prevalence of cell debris in the sperm suspension of mice treated for 35 and 45 days, the concluded a toxic effect of aflatoxin on sperm cells. They suggested that the decreased sperm count might indicate severe impact of AFB1 on spermatogenesis. They did not however compare the proportion of live sperm in the presence of aflatoxin in their study. Salem et al. (2001) have calculated dead sperm proportion in rabbit which is dose dependent. However they did not mentioned if aflatoxin had direct effect on sperm viability. In the present study we showed that aflatoxin has direct spermatotoxic effects on ejaculatory and epididymal ram sperm.
Sperm cell motility on the other hand is of great importance for penetrating the oocyte (Saling, 1990). However, it may not be necessary when intracytoplasmic sperm injection (ICSI) in performed (Ben Rhouma et al., 2003). Sperm motility has been reported to be 89.8% for rams when ejaculation frequency phase was once a day (Kaya et al., 2002). These researchers have not mentioned if it is total motility or progressive motility. In the present study however, the progressive motility of Chall ram ejaculatory sperm was 82.4%. This difference may be due to that the study was carried out in non-reproduction season or due to the species variations or the difference between total and progressive motility, which has been reported by Kaabi et al. (2003) for epididymal sperm. They have reported the progressive motility of ram epididymal sperm between 21.4 and 69.4%. In the present study epididymal sperm motility was 61%, which was similar to the mentioned workers. Ibeh et al. (2000) have incubated epididymal sperm of albino rats by in vitro fertilization medium (IVFM) supplemented with concentrations of 2.0, 4.0, 8.0 and 16.0 ppb aflatoxin B1. They observed that exposure to aflatoxin caused a significant reduction in the fertilization rates of rat oocytes even at the lowest concentration (2 ppb) of aflatoxin. They also noticed a significant reduction in the motility of sperm cells. No other data are available about the effect of aflatoxin on ram sperm motility in vitro. Brio et al. (2003) have reported that mycotoxins reduced sperm motility of boars. In the present study the reduction of sperm motility were observed after 2 h exposure to aflatoxin in al concentrations which were used. However no significant reduction in motility was seen after 1 h incubation in 1.96 ppb aflatoxin. The same motility patterns were observed for ejaculatory sperm.
REFERENCES
- Agnes, V.F. and M.A. Akbarsha, 2003. Spermatotoxic effect of aflatoxin B1 in the albino mouse. Food Chem. Toxicol., 41: 119-130.
CrossRefDirect Link - Ben-Rhouma, K., E. Ben-Miled, K. Attallah, H. Marrakchi, H. Khouja and M. Sakly, 2003. Successful pregnancies after using immotile spermatozoa from ejaculate, epididymis and testis. Eur. J. Obst. Gynecol. Reprod. Biol., 108: 182-185.
Direct Link - Blash, S., D. Melican and W. Gavin, 2000. Cryopreservation of epididymal sperm obtained at necropsy from goats. Theriogenology, 54: 899-905.
CrossRefPubMedDirect Link - Brio, K., I. Barna-Vetro, T. Pecsi, E. Szabo, G. Winkler, J. Fink-Germmels and L. Solti, 2003. Evaluation of spermatological parameters in ochratoxin A-challenged boars. Theriogenology, 60: 199-207.
CrossRefDirect Link - D'Mello, J.P.F. and A.M.C. Macdonald, 1997. Mycotoxins. Anim. Feed Sci. Technol., 69: 155-166.
Direct Link - De-Pauw, I.M.C., A. Van-Soom, K. Mintiens, S. Verberckmoes and A. De-Kruif, 2003. In vitro survival of bovine spermatozoa stored at room temperature under epididymal conditions. Theriogenology, 59: 1093-1107.
Direct Link - Egbunike, G.N., G.O. Emerole, T.A. Aire and F.I. Ikegwuonu, 1980. Sperm production rates, sperm physiology and fertility in rats chronically treated with sublethal doses of aflatoxin B1. Andrologia, 12: 467-475.
CrossRefDirect Link - Egbunike, G.N., 1982. Steroidogenic and spermatogenic potentials of the male rat after acute treatment with aflatoxin B1. Andrologia, 14: 440-446.
CrossRefDirect Link - Egbunike, G.N., 1985. Sperm maturation and storage in the male rat after acute treatment with aflatoxin B1. Andrologia, 17: 379-382.
CrossRefDirect Link - Hafez, A.H., A. Gomma, S.A. Mousa and S.E. Megalla, 1983. Aflatoxin and aflatoxicosis. IV. The effect of dietary aflatoxins on adult fertile male and female rabbits at various reproductive conditions. Mycopathologia, 83: 183-186.
CrossRefDirect Link - Ibeh, I.N., N. Uraih and J.I. Ogonar, 1994. Dietary exposure to aflatoxin in human male infertility in Benin City, Nigeria. Int. J. Fert. Menopausal Stud., 39: 208-214.
Direct Link - Ibeh, I.N. and D.K. Saxena, 1998. Effect of alpha-tocopherol supplementation on the impact of aflatoxin B1 on the testes of rats. Exp. Toxicol. Pathol., 50: 221-224.
Direct Link - Ibeh, I.N., D.K. Saxena and N. Uriah, 2000. Toxicology of aflatoxin: Effects on spermatozoa, oocytes and in vitro fertilization. J. Environ. Pathol. Toxicol. Oncol., 19: 357-361.
Direct Link - James, A.N., H. Green, S. Hoffman, A.M. Landry, D. Paccamonti and R.A. Godke, 2002. Preservation of equine sperm stored in the epididymis at 4°C for 24, 48, 72 and 96 h. Theriogenology, 58: 401-404.
Direct Link - Kaabi, M., P. Paz, M. Alvarez, E. Anel and J.C. Boixo et al., 2003. Effect of epididymis handling conditions on the quality of ram spermatozoa recovered post-mortem. Theriogenology, 60: 1249-1259.
CrossRefDirect Link - Kaya, A., M. Aksoy and T. Tekeli, 2002. Influence of ejaculation frequency on sperm characteristics, ionic composition and enzymatic activity of seminal plasma in rams. Small. Rumin. Res., 44: 153-158.
Direct Link - Minervini, F., M.E. Dell`Aquila, F. Maritato, P. MInoia and A. Visconti, 2001. Toxic effects of the mycotoxin zearalenone and its derivatives on in vitro maturation of bovine oocytes and 17β-estradiol levels in mural granulose cell cultures. Toxicol. In vitro, 15: 489-495Toxic effects of the mycotoxin zearalenone and its derivatives on in vitro maturation of bovine oocytes and 17β-estradiol levels in mural granulose cell cultures. Toxicol. In vitro, 15: 489-495.
Direct Link - Patrizio, P., 2000. Cryopreservation of epididymal sperm. Mol. Cell. Endocrinol., 169: 11-14.
CrossRefDirect Link - Picha, J., J. Cerovsky and D. Pichova, 1986. [Fluctuation in the concentration of sex steroids and aflatoxin B1 in the seminal plasma of boars and its relation to sperm production]. Veterinarni Medicina, 31: 347-357.
PubMed - Puschner, B., 2002. Mycotoxins. Vet. Clin. North Am.: Small Anim. Pract., 32: 409-419.
CrossRefPubMedDirect Link - Salem, M.H., K.I. Kamel, M.I. Yousef, G.A. Hassan and F.D. EL-Nouty, 2001. Protective role of ascorbic acid to enhance semen quality of rabbits treated with sublethal doses of aflatoxin B1. Toxicology, 162: 209-218.
CrossRefDirect Link - Sinha, S.P. and V. Prasad, 1990. Effect of dietary concentration of crude aflatoxin on meiotic chromosomes, sperm morphology and sperm count in mice, Mus musculus. Proc. Indian National Sci. Acad. Part B, Biol. Sci., 56: 269-275.
Direct Link - Sinha, S.P. and S. Bose, 1992. Effect of dietary concentration of aflatoxin B1 and vitamin C on meiotic chromosomes, sperm head morphology and sperm count in mice, Mus musculus. Cytologia, 57: 75-80.
Direct Link - Solti, A., T. Pecsi, I.B. Vetro, F. Szasz, K. Biro and E. Szabo, 1999. Analysis of serum and seminal plasma after feeding ocharatoxin A with breeding boars. Anim. Reprod. Sci., 56: 123-132.
Direct Link - Tajik, P., W.H. Wang, K. Okuda and K. Niwa, 1994. In vitro fertilization of bovine oocytes in a chemically defined, protein-free medium varying the bicarbonate concentration. Biol. Reprod., 50: 1231-1237.
Direct Link - Uriah, N., I.N. Ibeh and F. Oluwafemi, 2001. A study on the impact of aflatoxin on human reproduction. Afr. J. Reprod. Health, 5: 106-110.
Direct Link - Verma, R.J. and A. Nair, 2002. Effect of aflatoxins on testicular esteroidogenesis and amelioration by vitamin E. Food Chem. Toxicol., 40: 669-672.
Direct Link - Yu, I. and S.P. Leibo, 2002. Recovery of motile, membrane-intact spermatozoa from canine epididymides stored for 8 days at 4°C. Theriogenology, 57: 1179-1190.
Direct Link